Constructed Wetlands: A Review on the Role of Radial Oxygen Loss in the Rhizosphere by Macrophytes

Abstract
:1. Applications and Mechanism of Constructed Wetlands
2. Role of Macrophytes in CWs
2.1. Direct Functions
2.2. Coupling Effects between Plants and Microorganisms
2.3. Other Functions
3. Role of ROL of Macrophytes
3.1. Definition and Theory of ROL
3.2. Factors that Affect ROL
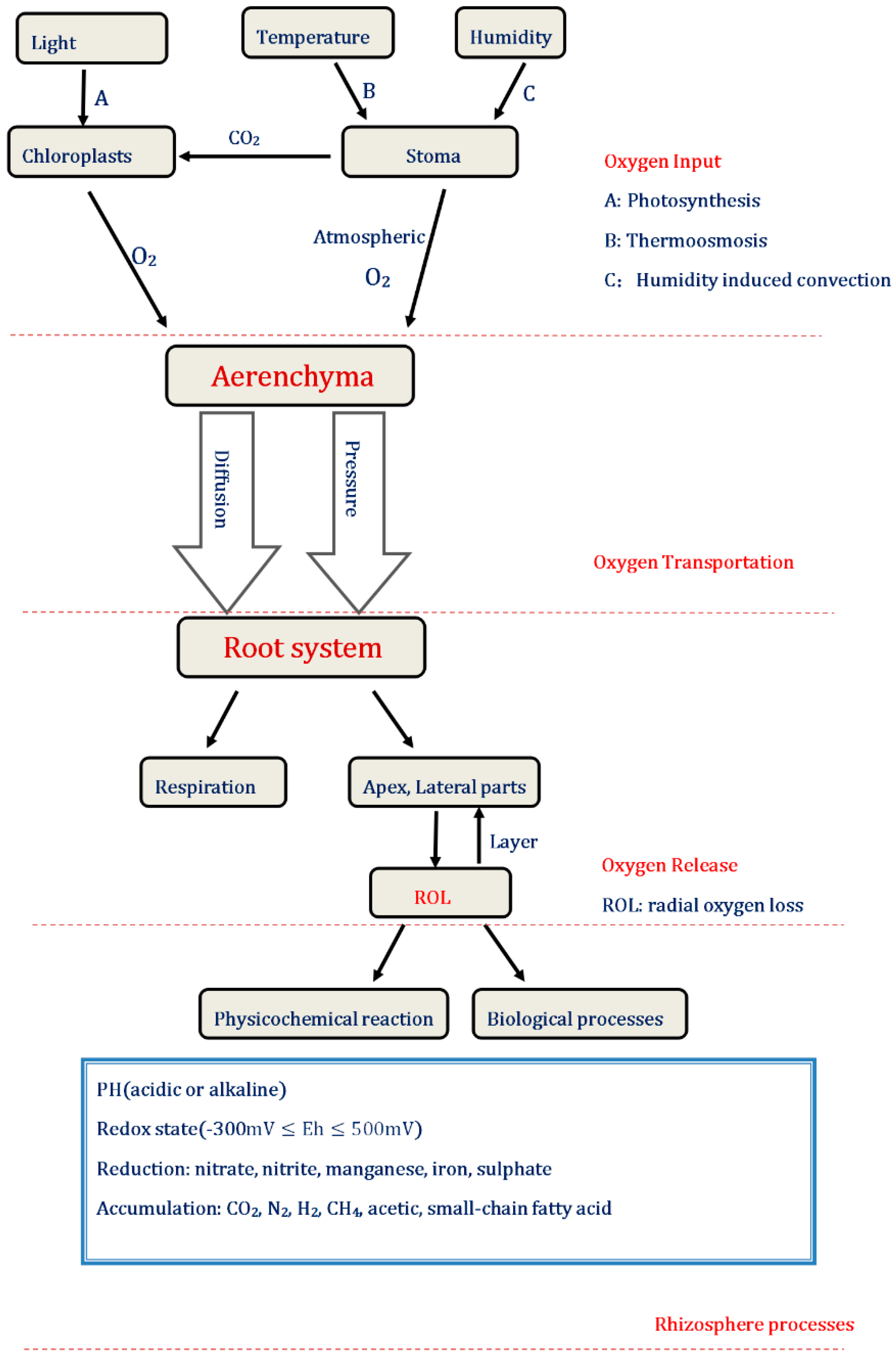
3.2.1. Perspectives on the Input of Oxygen
3.2.2. Perspectives on the Transportation of Oxygen
3.2.3. Perspectives on the Release of Oxygen
4. Measurement of ROL
5. Role of ROL in Wastewater Treatment
5.1. Effects on Organic Matter Removal
5.2. Effects on Nitrogen Removal
5.3. Effects on Heavy Metal Uptake
6. Concluding Remarks
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Vymazal, J. Constructed wetlands for wastewater treatment: Five decades of experience. Environ. Sci. Technol. 2010, 45, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Cao, F.-Q.; Kong, Q.; Zhou, L.-L.; Yuan, Q.; Zhu, Y.-J.; Wang, Q.; Du, Y.-D.; Wang, Z.-D. Electricity production and evolution of microbial community in the constructed wetland-microbial fuel cell. Chem. Eng. J. 2018, 339, 479–486. [Google Scholar] [CrossRef]
- Yang, C.H.; Crowley, D.E. Rhizosphere microbial community structure in relation to root location and plant iron nutritional status. Appl. Environ. Microbiol. 2000, 1, 345–351. [Google Scholar] [CrossRef]
- Yu, Y.; Li, X.; Cheng, J. A Comparison Study of Mechanism: Cu2+ Adsorption on Different Adsorbents and Their Surface-Modified Adsorbents. J. Mater. Chem. 2016, 2016, 7936258. [Google Scholar] [CrossRef]
- Kong, Q.; Zhang, J.; Miao, M.; Tian, L.; Guo, N.; Liang, S. Partial nitrification and nitrous oxide emission in an intermittently aerated sequencing batch biofilm reactor. Chem. Eng. J. 2013, 217, 435–441. [Google Scholar] [CrossRef]
- Tao, M.; He, F.; Xu, D.; Li, M.; Wu, Z. How artificial aeration improved sewage treatment of an integrated vertical-flow constructed wetland. Pol. J. Environ. Stud. 2010, 19, 183–191. [Google Scholar]
- Wu, H. Long-Term Performance and Mechanism in the Surface Constructed Wetland for Treating Polluted River Water in Northern China. Master’s Thesis, Shandong University, Jinan, Shandong, China, 2011. [Google Scholar]
- Ávila, C.; Reyes, C.; Bayona, J.M.; García, J. Emerging organic contaminant removal depending on primary treatment and operational strategy in horizontal subsurface flow constructed wetlands: Influence of redox. Water Res. 2013, 47, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Braeckevelt, M.; Reiche, N.; Trapp, S.; Wiessner, A.; Paschke, H.; Kuschk, P.; Kaestner, M. Chlorobenzene removal efficiencies and removal processes in a pilot-scale constructed wetland treating contaminated groundwater. Ecol. Eng. 2011, 37, 903–913. [Google Scholar] [CrossRef]
- Kickuth, R. Degradation and Incorporation of Nutrients from Rural Waste Waters by Plant Rhizosphere under Limnic Conditions; European Commission: London, UK, 1977; pp. 335–343. [Google Scholar]
- Stottmeister, U.; Wießner, A.; Kuschk, P.; Kappelmeyer, U.; Kästner, M.; Bederski, O.; Müller, R.A.; Moormann, H. Effects of plants and microorganisms in constructed wetlands for wastewater treatment. Biotechnol. Adv. 2003, 22, 93–117. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.Q.; Chen, Y.H.; Shen, Z.G.; Zheng, L.Q. Roles of Cell Wall in Plant Heavy Metal Tolerance. J. Plant Physiol. 2014, 50, 605–611. [Google Scholar]
- Sandermann, H., Jr. Plant metabolism of xenobiotics. Trends Biochem. Sci. 1992, 17, 82–84. [Google Scholar] [CrossRef]
- Jackson, M.B.; Armstrong, W. Formation of Aerenchyma and the Processes of Plant Ventilation in Relation to Soil Flooding and Submergence. J. Plant Physiol. 1999, 1, 274–287. [Google Scholar] [CrossRef]
- Walker, T.S.; Bais, H.P.; Grotewold, E.; Vivanco, J.M. Root exudation and rhizosphere biology. Plant Physiol. 2003, 132, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Gagnon, V.; Chazarenc, F.; Comeau, Y.; Brisson, J. Influence of macrophyte species on microbial density and activity in constructed wetlands. Water Sci. Technol. 2007, 56, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Bais, H.P.; Weir, T.L.; Perry, L.G.; Gilroy, S.; Vivanco, J.M. The role of root exudates in rhizosphere interactions with plants and other organisms. Annu. Rev. Plant Biol. 2006, 57, 233–266. [Google Scholar] [CrossRef] [PubMed]
- Rehman, F.; Pervez, A.; Khattak, B.N.; Ahmad, R. Constructed Wetlands: Perspectives of the Oxygen Released in the Rhizosphere of Macrophytes. Clean-Soil Air Water 2017, 45. [Google Scholar] [CrossRef]
- Sun, T.R.; Long, C.; Wang, Q.Y.; Zhou, D.M.; Cheng, J.M.; Xu, H. Roles of abiotic losses, microbes, plant roots, and root exudates on phytoremediation of PAHs in a barren soil. J. Hazard. Mater. 2010, 176, 919–925. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R. Rhizosphere biology of aquatic microbes in order to access their bioremediation potential along with different aquatic macrophytes. Recent Res. Sci. Technol. 2013, 5, 29–32. [Google Scholar]
- Bucher, M. Functional biology of plant phosphate uptake at root and mycorrhiza interfaces. New Phytol. 2007, 173, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Spaepen, S.; Dobbelaere, S.; Croonenborghs, A.; Vanderleyden, J. Effects of Azospirillum brasilense indole-3-acetic acid production on inoculated wheat plants. Plant Soil 2008, 312, 15–23. [Google Scholar] [CrossRef]
- Armstrong, J.; Armstrong, W. Rice and Phragmites: Effects of organic acids on growth, root permeability, and radial oxygen loss to the rhizosphere. Am. J. Bot. 2001, 88, 1359–1370. [Google Scholar] [CrossRef] [PubMed]
- Shelef, O.; Gross, A.; Rachmilevitch, S. Role of Plants in a Constructed Wetland. Current and New Perspectives. Water 2013, 5, 405–419. [Google Scholar] [CrossRef]
- Zong, W.; Sun, F.; Pei, H.; Hu, W.; Pei, R. Microcystin-associated disinfection by-products: The real and non-negligible risk to drinking water subject to chlorination. Chem. Eng. J. 2015, 279, 498–506. [Google Scholar] [CrossRef]
- Miao, M.S.; Liu, Q.; Shu, L.; Wang, Z.; Liu, Y.Z.; Kong, Q. Removal of cephalexin from effluent by activated carbon prepared from alligator weed: Kinetics, isotherms, and thermodynamic analyses. Process Saf. Environ. 2016, 104, 481–489. [Google Scholar] [CrossRef]
- Caffrey, J.M.; Kemp, W.M. Seasonal and spatial patterns of oxygen production, respiration and root-rhizome release in Potamogeton perfoliatus L. and Zostera marina L. Aquat. Bot. 1991, 40, 109–128. [Google Scholar] [CrossRef]
- Konnerup, D.; Sorrell, B.K.; Brix, H. Do tropical wetland plants possess convective gas flow mechanisms? New Phytol. 2011, 190, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Xie, H.; Zhang, J.; Liang, S.; Ngo, H.H.; Guo, W.; Liu, C.; Zhao, C.; Li, H. Effect of plant harvesting on the performance of constructed wetlands during winter: Radial oxygen loss and microbial characteristics. Environ. Sci. Pollut. Res. 2015, 22, 7476–7484. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Yang, Y.; Li, L.I.; Mai, X. Characteristics of growth and radial oxygen loss of eight wetland plants. J. Lake Sci. 2015, 27, 1042–1048. [Google Scholar]
- Liu, Z.K.; Niu, K.K.; Ma, Q.L.; Bai, X.H.; Su, L.X. Study on rate of roots radial oxygen loss of eight wetland plants. Guizhou Agric. Sci. 2010, 38, 47–50. [Google Scholar]
- Brix, H.; Schierup, H.H. Soil oxygenation in constructed reed beds. The role of macrophyte and soil-atmosphere interface oxygen transport. In Proceedings of the International Conference on the Use of Constructed Wetlands in Water Pollution Control, Cambridge, UK, 24–28 September 1990; pp. 53–66. [Google Scholar]
- Dong, C.; Zhu, W.; Gao, M.; Zhao, L.F.; Huang, J.Y.; Zhao, Y.Q. Diurnal fluctuations in oxygen release from roots of Acorus calamus Linn in a modeled constructed wetland. J. Environ. Sci. Health B 2011, 46, 224–229. [Google Scholar] [CrossRef] [PubMed]
- Tao, M.A.; Neng, Y.I.; Zhang, Z.H.; Wang, Y.; Gao, Y.; Yan, S.H. Oxygen and Organic Carbon Releases from Roots of Eichhornia Crassipes and Their Influence on Transformation of Nitrogen in Water. J. Agroenviron. Sci. 2014, 33, 2003–2013. [Google Scholar]
- Tian, Q.; Wang, P.F.; Ouyang, P.; Wang, C.; Zhang, W.M. Purification of eutrophic water with five submerged hydrophytes. Water Resour. Prot. 2009, 25, 14–17. [Google Scholar]
- Su, W.-H.; Zhang, G.-F.; Zhang, Y.-S.; Xiao, H.; Xia, F. Photosynthetic characteristics of 5 species of Submerged Macrophytes. Acta Ecol. 2004, 28, 391–395. [Google Scholar]
- Armstrong, W. Root aeration in the wetland condition. Plant Life Anaerob. Environ. 1978, 1, 269–297. [Google Scholar]
- Armstrong, J.; Armstrong, W.; Beckett, P.M. Phragmites australis. Venturi and humidity-induced pressure flows enhance rhizome aeration and rhizosphere oxidation. New Phytol. 1992, 120, 197–207. [Google Scholar] [CrossRef]
- Connell, E.L.; Colmer, T.D.; Walker, D.I. Radial oxygen loss from intact roots of Halophila ovalis as a function of distance behind the root tip and shoot illumination. Aquat. Bot. 1999, 63, 219–228. [Google Scholar] [CrossRef]
- Emerson, R.; Lewis, C.M. Factors influencing the efficiency of photosynthesis. Am. J. Bot. 1939, 26, 808–822. [Google Scholar] [CrossRef]
- Agnew, D.J.; Taylor, A.C. Effects of oxygen tension, temperature, salinity, and humidity on the survival of two intertidal gammarid amphipods. Mar. Ecol. Prog. Ser. 1986, 32, 27–33. [Google Scholar] [CrossRef]
- Armstrong, W. Aeration in Higher Plants. Adv. Bot. Res. 1980, 7, 225–332. [Google Scholar]
- Colmer, T.D. Aerenchyma and an Inducible Barrier to Radial Oxygen Loss Facilitate Root Aeration in Upland, Paddy and Deep-water Rice (Oryza sativa L.). Ann. Bot. Lond. 2003, 91, 301–309. [Google Scholar] [CrossRef]
- Garthwaite, A.J.; Rv, B.; Colmer, T.D. Diversity in root aeration traits associated with waterlogging tolerance in the genus Hordeum. Funct. Plant Biol. 2003, 30, 875–889. [Google Scholar] [CrossRef]
- Nikolausza, M.; Székely, A.; Rusznyák, A.; Márialigeti, K.; Kästner, M. Diurnal redox fluctuation and microbial activity in the rhizosphere of wetland plants. Eur. J. Soil Biol. 2008, 44, 324–333. [Google Scholar] [CrossRef]
- Soda, S.; Ike, M.; Ogasawara, Y.; Yoshinaka, M.; Mishima, D.; Fujita, M. Effects of light intensity and water temperature on oxygen release from roots into water lettuce rhizosphere. Water Res. 2007, 41, 487–491. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wu, H.; Hu, Z.; Liang, S.; Fan, J. Examination of oxygen release from plants in constructed wetlands in different stages of wetland plant life cycle. Environ. Sci. Pollut. Res. 2014, 21, 9709–9716. [Google Scholar] [CrossRef] [PubMed]
- Visser, E.J.W.; Colmer, T.D.; Blom, C.W.P.M.; Voesenek, L.A.C.J. Changes in growth, porosity, and radial oxygen loss from adventitious roots of selected mono- and dicotyledonous wetland species with contrasting types of aerenchyma. Plant Cell Environ. 2000, 23, 1237–1245. [Google Scholar] [CrossRef]
- Luxmoore, R.J.; Stolzy, L.H.; Letey, J. Oxygen diffusion in the soil-plant system. 2. Respiration rate, permeability, and porosity of consecutive excised segments of maize and rice roots. Agron. J. 1970, 62, 322–324. [Google Scholar] [CrossRef]
- Vymazal, J. Emergent plants used in free water surface constructed wetlands: A review. Ecol. Eng. 2013, 61, 582–592. [Google Scholar] [CrossRef]
- Xu, F.; Qi, X.-Y.; Kong, Q.; Shu, L.; Miao, M.-S.; Xu, S.; Du, Y.-D.; Wang, Q.; Liu, Q.; Ma, S.-S. Adsorption of sunset yellow by luffa sponge, modified luffa and activated carbon from luffa sponge. Desalin. Water Treat. 2017, 96, 86–96. [Google Scholar] [CrossRef]
- Lawson, G. Cultivating Reeds (Phragmites australis) for Root Zone Treatment of Sewage; Contract Report ITE Project 965; Water Research Center: Cumbria, UK, 1985. [Google Scholar]
- Taylor, C.R.; Hook, P.B.; Stein, O.R.; Zabinski, C.A. Seasonal effects of 19 plant species on COD removal in subsurface treatment wetland microcosms. Ecol. Eng. 2011, 37, 703–710. [Google Scholar] [CrossRef]
- Pedersen, O.; Binzer, T.; Borum, J. Sulphide intrusion in eelgrass (Zostera marina L.). Plant Cell Environ. 2004, 27, 595–602. [Google Scholar] [CrossRef]
- Colmer, T.D. Long-distance transport of gases in plants: A perspective on internal aeration and radial oxygen loss from roots. Plant Cell Environ. 2003, 26, 17–36. [Google Scholar] [CrossRef]
- Kojima, M.; Sakuragi, H.; Tokumaru, K. ChemInform Abstract: The role of oxygen as an electron acceptor in dimerization of some styrene derivatives. Tetrahedron Lett. 1981, 12, 2889–2892. [Google Scholar] [CrossRef]
- Cheng, X.Y.; Wang, M.; Zhang, C.F.; Wang, S.Q.; Chen, Z.H. Relationships between plant photosynthesis, radial oxygen loss and nutrient removal in constructed wetland microcosms. Biochem. Syst. Ecol. 2014, 54, 299–306. [Google Scholar] [CrossRef]
- Wang, Z.B.; Miao, M.S.; Kong, Q.; Ni, S.Q. Evaluation of microbial diversity of activated sludge in a municipal wastewater treatment plant of northern China by high-throughput sequencing technology. Desalin. Water Treat. 2016, 57, 1–6. [Google Scholar] [CrossRef]
- Fonseca, A.L.D.S.; Marinho, C.C.; Esteves, F.D.A. Dynamics of dissolved organic carbon from aerobic and anaerobic decomposition of Typha domingensis Pers. and Eleocharis interstincta (Vahl) Roem. & Schult. in a tropical coastal lagoon. Acta Limnologica Brasiliensia 1959, 25, 279–290. [Google Scholar]
- Kristanto, G.A.; Asaloei, H. Assessment of anaerobic biodegradability of five different solid organic wastes. AIP Conf. Proc. 2017, 1826, 020029. [Google Scholar] [CrossRef]
- Kim, J.K.; Park, K.J.; Cho, K.S.; Nam, S.W.; Park, T.J.; Bajpai, R. Aerobic nitrification-denitrification by heterotrophic Bacillus strains. Bioresour. Technol. 2005, 96, 1897–1906. [Google Scholar] [CrossRef] [PubMed]
- Brix, H. Treatment of Wastewater in the Rhizosphere of Wetland Plants-the Root-Zone Method. Water Sci. Technol. 1987, 19, 107–118. [Google Scholar] [CrossRef]
- Pochana, K.; Keller, J. Study of factors affecting simultaneous nitrification and denitrification (SND). Water Sci. Technol. 1999, 39, 61–68. [Google Scholar] [CrossRef]
- Kong, Q. Impact of ammonium and salinity concentrations on Nitrous Oxide emission in partial nitrification system. KSCE J. Civ. Eng. 2015, 19, 873–879. [Google Scholar] [CrossRef]
- Yang, J.; Ma, Z.; Ye, Z.; Guo, X.; Qiu, R. Heavy metal (Pb, Zn) uptake and chemical changes in rhizosphere soils of four wetland plants with different radial oxygen loss. J. Environ. Sci. 2010, 22, 696–702. [Google Scholar] [CrossRef]
- Ju, L.W. Iron and Mn plaques on the surface of roots of wetland plants. Acta Chim. Sin. 2005, 25, 358–363. [Google Scholar]
- Lv, J.; Liu, Y.; Zhang, Z.; Dai, J. Factorial kriging and stepwise regression approach to identify environmental factors influencing spatial multi-scale variability of heavy metals in soils. J. Hazard. Mater. 2013, 261, 387–397. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.L.; Hu, Z.Y.; Ziadi, N.; Xia, X. Excessive sulfur supply reduces cadmium accumulation in brown rice (Oryza sativa L.). Environ. Pollut. 2010, 158, 409–415. [Google Scholar] [CrossRef] [PubMed]
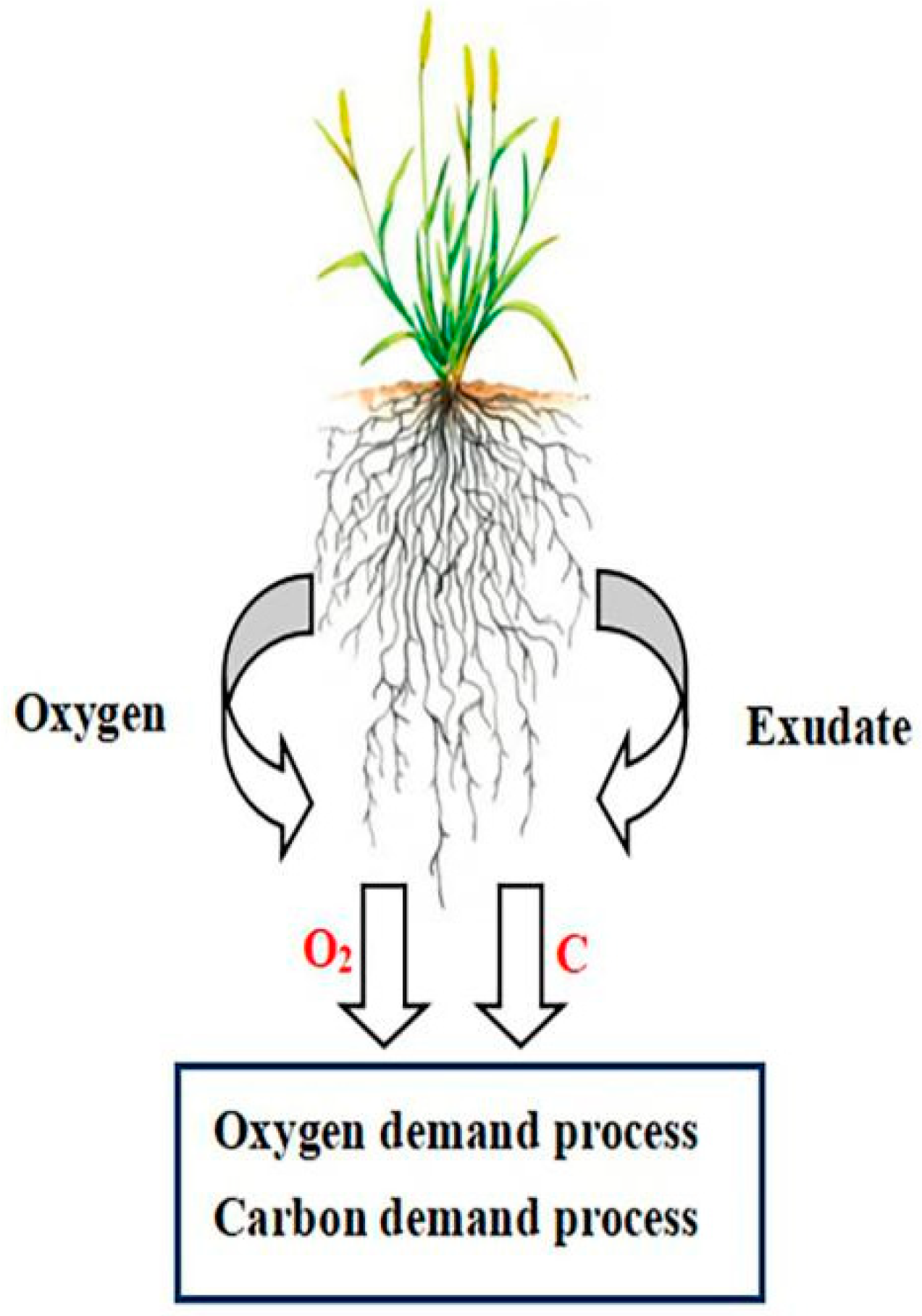

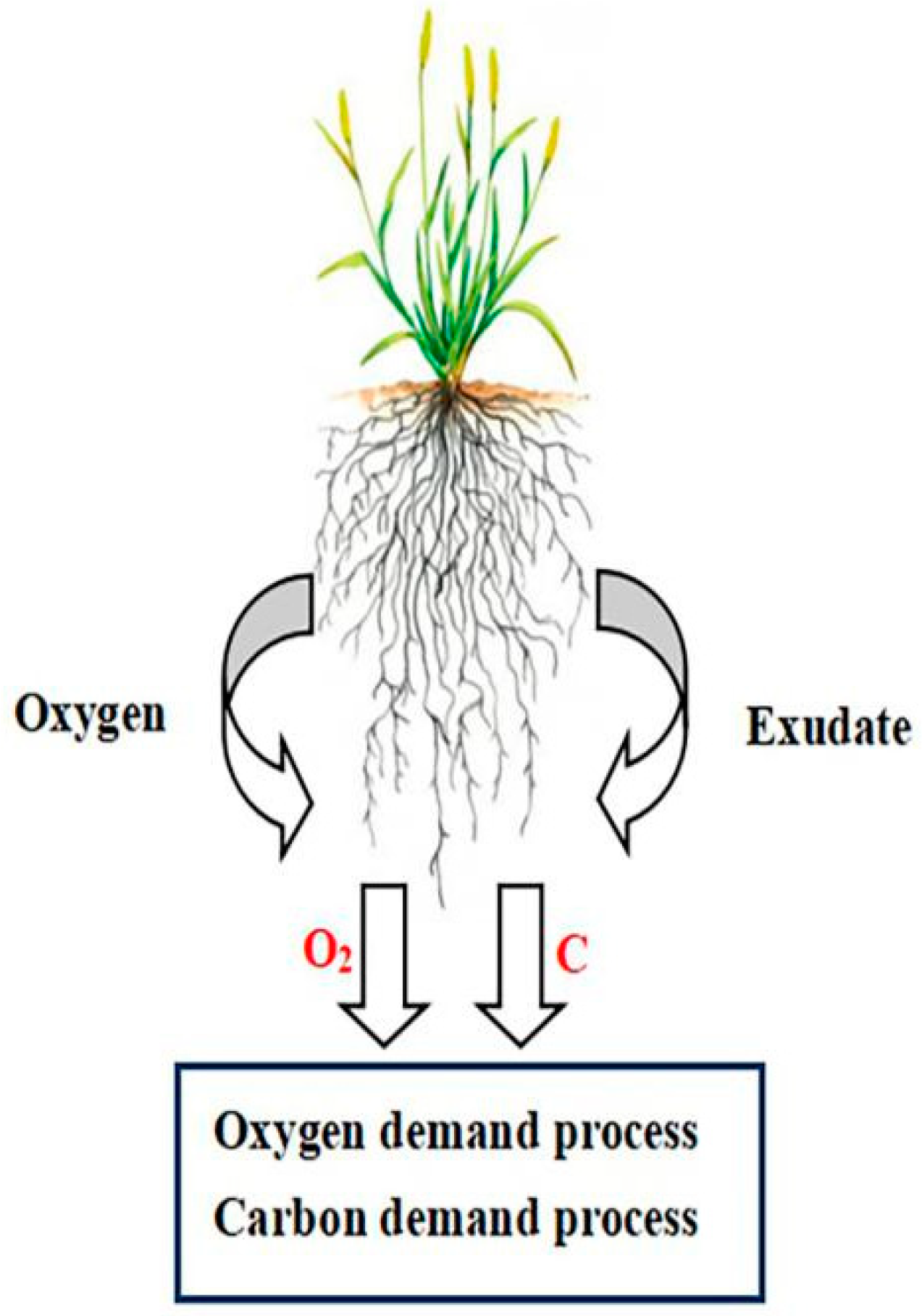{kind=link}
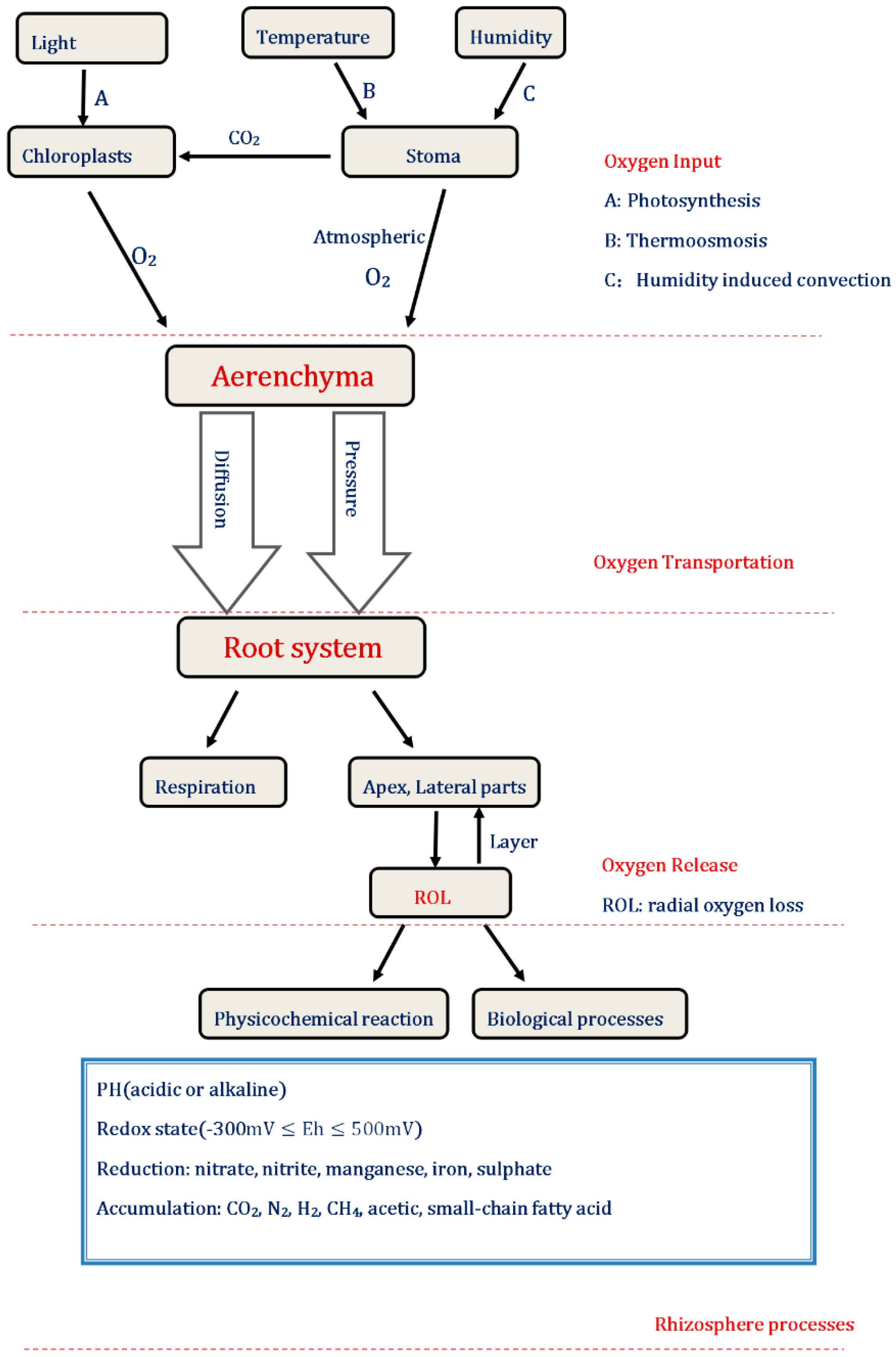{kind=link}
| Type | Macrophytes | ROL Rate (µmol O2 d−1 g−1 DWroot) 1 | Ref. |
|---|---|---|---|
| Emerging plant | Bog rush | 177.87 ± 10.02 | [30] |
| Cattail | 124.82 ± 26.97 | [30] | |
| Bamboo reed | 84.13 ± 5.99 | [30] | |
| Zizania aquatica | 50.98–95.89 | [31,32] | |
| Phragmites australis | 105.71–253.72 | [32] | |
| Calamus | 117.46 ± 17.35 | [33] | |
| Canna | 120.54 ± 1.52 | [30] | |
| Floating plant | Arrowhead | 126.64 | [31] |
| Calla | 103.05 | [31] | |
| Water hyacinth | 19.8–67.15 | [34] | |
| Submerged plant 2 | Eel grass | 190.66 | [35,36] |
| Hornwort | 367.88 | [35,36] | |
| Black algae | 320.96 | [35,36] | |
| Myriophyllum verticillatum | 263.26 | [35,36] | |
| Water caltrop | 332.91 | [35,36] |
| Measurement of ROL | Advantages | Disadvantages |
|---|---|---|
| The soaking method of root solution | Measure whole macrophyte, low damage | Low measured value |
| Titrimetric method | Analogy the conditions of the redox potential in the soil | Higher measurement of oxygen, less accurate |
| Oxygen micro-electrode method | Able to simulate soil conditions from the day to night | More expensive, fragile electrode |
| Sediment redox potential | Actual time determination | Easily influenced by environmental factors |
| Mathematical model method | Provide a theoretical basis | Easily limited by the data and assumptions |
| Measurement of rhizoplane | Well studied the root/soil interface soil conditions, easy to control | Different characteristics of the root system compared with non-interfering soil macrophytes |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Hu, Y.; Xie, H.; Yang, Z. Constructed Wetlands: A Review on the Role of Radial Oxygen Loss in the Rhizosphere by Macrophytes. Water 2018, 10, 678. https://doi.org/10.3390/w10060678
Wang Q, Hu Y, Xie H, Yang Z. Constructed Wetlands: A Review on the Role of Radial Oxygen Loss in the Rhizosphere by Macrophytes. Water. 2018; 10(6):678. https://doi.org/10.3390/w10060678
Chicago/Turabian StyleWang, Qian, Yanbiao Hu, Huijun Xie, and Zhongchen Yang. 2018. "Constructed Wetlands: A Review on the Role of Radial Oxygen Loss in the Rhizosphere by Macrophytes" Water 10, no. 6: 678. https://doi.org/10.3390/w10060678
APA StyleWang, Q., Hu, Y., Xie, H., & Yang, Z. (2018). Constructed Wetlands: A Review on the Role of Radial Oxygen Loss in the Rhizosphere by Macrophytes. Water, 10(6), 678. https://doi.org/10.3390/w10060678