
The Tumor Suppressor p53 in Mucosal Melanoma of the Head and Neck

{kind=link}
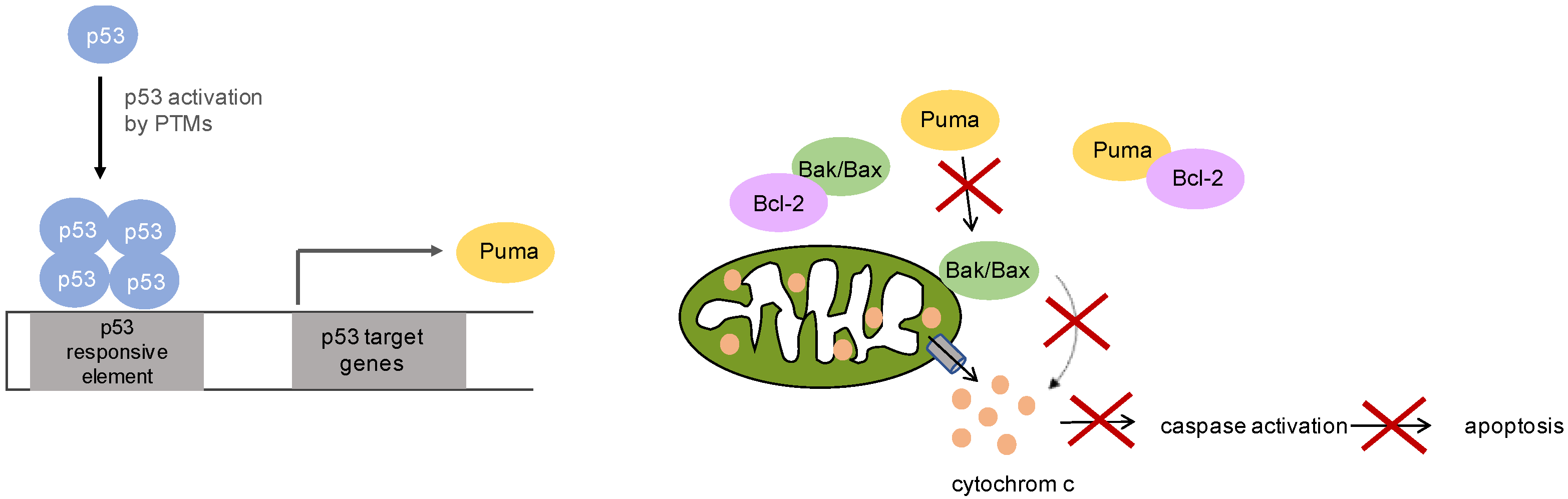{kind=link}
Abstract
:1. Introduction
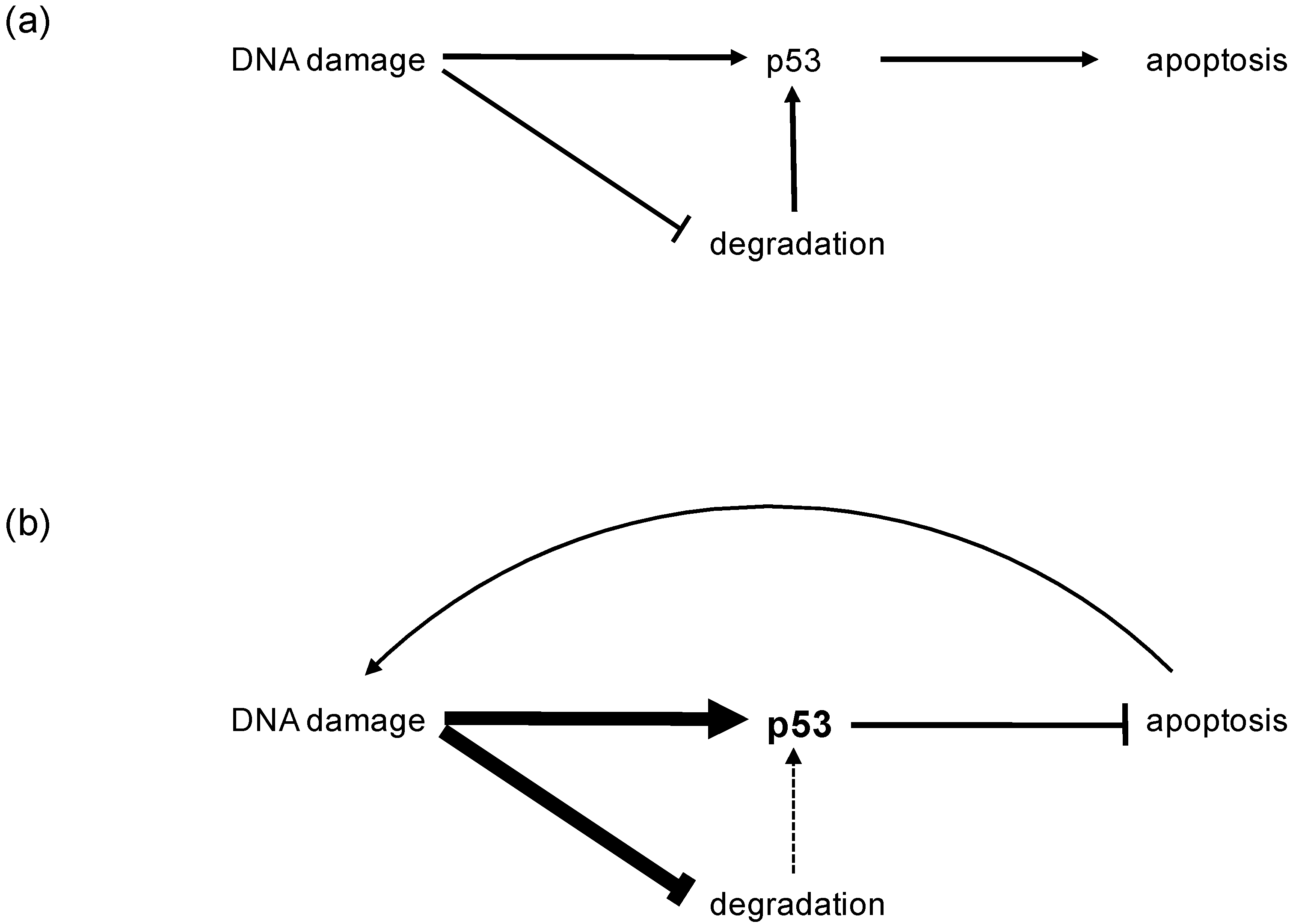
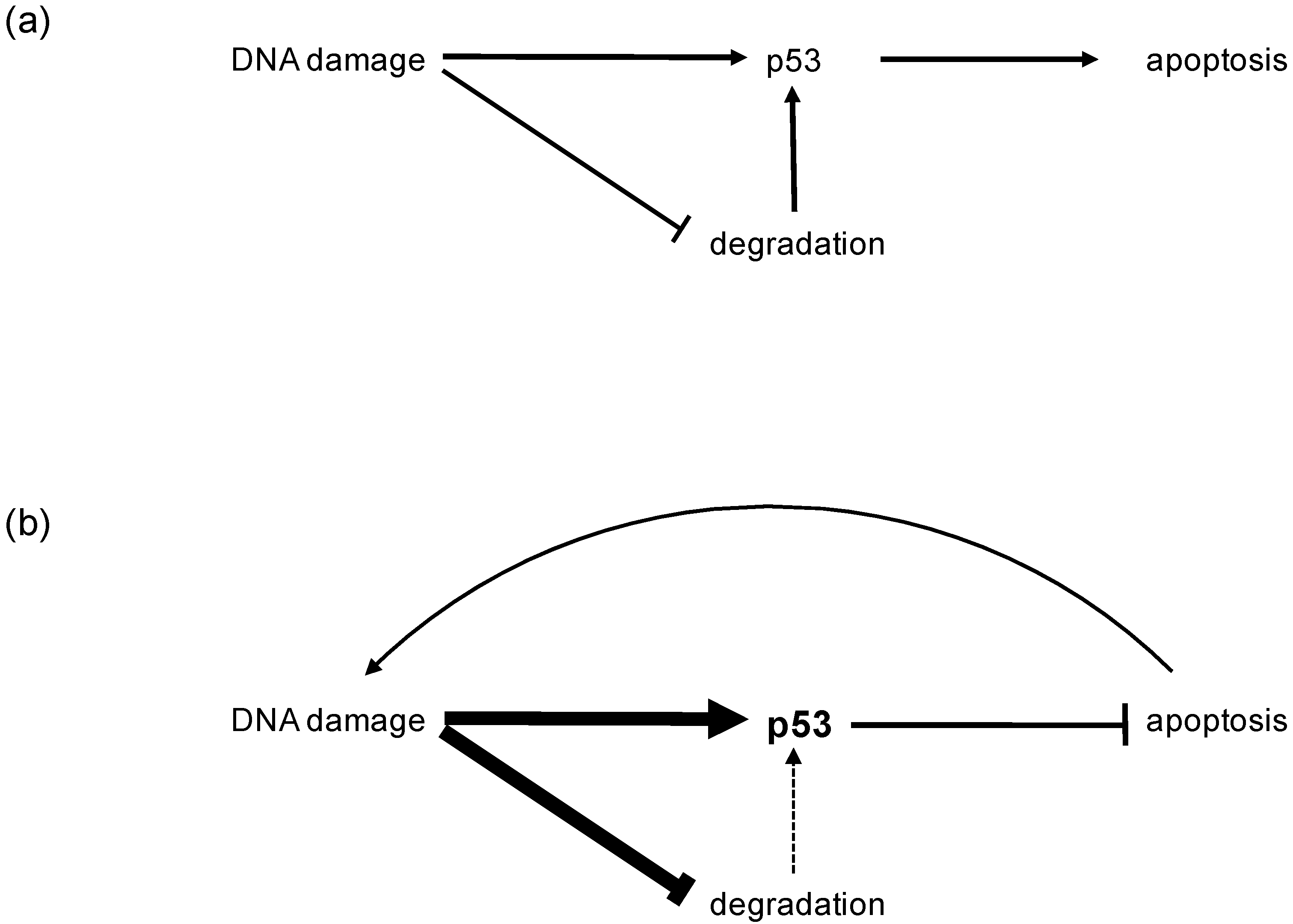
2. Physiology of the p53 Tumor Suppressor
3. Genetic Aberrations in Mucosal Melanoma
4. p53 Protein Expression in Mucosal Melanoma
5. p53 Signaling in Mucosal Melanoma
6. Conclusions
Author Contributions
Conflicts of Interest
Financial Disclosure
References
- Kim, H.S.; Jung, M.; Kang, H.N.; Kim, H.; Park, C.W.; Kim, S.M.; Shin, S.J.; Kim, S.H.; Kim, S.G.; Kim, E.K.; et al. Oncogenic BRAF fusions in mucosal melanomas activate the MAPK pathway and are sensitive to MEK/PI3K inhibition or MEK/CDK4/6 inhibition. Oncogene 2017, 36, 3334–3345. [Google Scholar] [CrossRef] [PubMed]
- Lengyel, E.; Gilde, K.; Remenár, E.; Esik, O. Malignant mucosal melanoma of the head and neck. Pathol. Oncol. Res. 2003, 9, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Papaspyrou, G.; Garbe, C.; Schadendorf, D.; Werner, J.A.; Hauschild, A.; Egberts, F. Mucosal melanomas of the head and neck: New aspects of the clinical outcome, molecular pathology, and treatment with c-kit inhibitors. Melanoma Res. 2011, 21, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Mendenhall, W.M.; Amdur, R.J.; Hinerman, R.W.; Werning, J.W.; Villaret, D.B.; Mendenhall, N.P. Head and neck mucosal melanoma. Am. J. Clin. Oncol. 2005, 28, 626–630. [Google Scholar] [CrossRef] [PubMed]
- Tas, F.; Keskin, S.; Karadeniz, A.; Dagoglu, N.; Sen, F.; Kilic, L.; Yildiz, I. Noncutaneous melanoma have distinct features from each other and cutaneous melanoma. Oncology 2011, 81, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Frakes, J.M.; Strom, T.J.; Naghavi, A.O.; Trotti, A.; Rao, N.G.; McCaffrey, J.C.; Otto, K.J.; Padhya, T.; Caudell, J.J. Outcomes of mucosal melanoma of the head and neck. J. Med. Imaging Radiat. Oncol. 2016, 60, 268–273. [Google Scholar] [CrossRef] [PubMed]
- Lourenco, S.V.; Fernandes, J.D.; Hsieh, R.; Coutinho-Camillo, C.M.; Bologna, S.; Sangueza, M.; Nico, M.M. Head and neck mucosal melanoma: A review. Am. J. Dermatopathol. 2014, 36, 578–587. [Google Scholar] [CrossRef] [PubMed]
- Mackintosh, J.A. The antimicrobial properties of melanocytes, melanosomes and melanin and the evolution of black skin. J. Theor. Biol. 2001, 211, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Thierauf, J.; Veit, J.; Doscher, J.; Theodoraki, M.N.; Greve, J.; Hoffmann, T.K. Mucosal melanoma of the head and neck. Laryngorhinootologie 2015, 94, 812–818. [Google Scholar] [PubMed]
- Fritsche, M.K.; Metzler, V.; Becker, K.; Plettenberg, C.; Heiser, C.; Hofauer, B.; Knopf, A. Cisplatin fails to induce puma mediated apoptosis in mucosal melanomas. Oncotarget 2015, 6, 9887–9896. [Google Scholar] [CrossRef] [PubMed]
- Mihajlovic, M.; Vlajkovic, S.; Jovanovic, P.; Stefanovic, V. Primary mucosal melanomas: A comprehensive review. Int. J. Clin. Exp. Pathol. 2012, 5, 739–753. [Google Scholar] [PubMed]
- Kandoth, C.; McLellan, M.D.; Vandin, F.; Ye, K.; Niu, B.; Lu, C.; Xie, M.; Zhang, Q.; McMichael, J.F.; Wyczalkowski, M.A.; et al. Mutational landscape and significance across 12 major cancer types. Nature 2013, 502, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Soussi, T.; Ishioka, C.; Claustres, M.; Beroud, C. Locus-specific mutation databases: Pitfalls and good practice based on the p53 experience. Nat. Rev. Cancer 2006, 6, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Brooks, C.L.; Gu, W. New insights into p53 activation. Cell Res. 2010, 20, 614–621. [Google Scholar] [CrossRef] [PubMed]
- Riley, T.; Sontag, E.; Chen, P.; Levine, A. Transcriptional control of human p53-regulated genes. Nat. Rev. Mol. Cell Biol. 2008, 9, 402–412. [Google Scholar] [CrossRef] [PubMed]
- Oren, M. Regulation of the p53 tumor suppressor protein. J. Biol. Chem. 1999, 274, 36031–36034. [Google Scholar] [CrossRef] [PubMed]
- Harris, S.L.; Levine, A.J. The p53 pathway: Positive and negative feedback loops. Oncogene 2005, 24, 2899–2908. [Google Scholar] [CrossRef] [PubMed]
- Blagosklonny, M.V. Loss of function and p53 protein stabilization. Oncogene 1997, 15, 1889–1893. [Google Scholar] [CrossRef] [PubMed]
- Ragnarsson-Olding, B.K.; Karsberg, S.; Platz, A.; Ringborg, U.K. Mutations in the TP53 gene in human malignant melanomas derived from sun-exposed skin and unexposed mucosal membranes. Melanoma Res. 2002, 12, 453–463. [Google Scholar] [CrossRef] [PubMed]
- Gwosdz, C.; Scheckenbach, K.; Lieven, O.; Reifenberger, J.; Knopf, A.; Bier, H.; Balz, V. Comprehensive analysis of the p53 status in mucosal and cutaneous melanomas. Int. J. Cancer 2006, 118, 577–582. [Google Scholar] [CrossRef] [PubMed]
- Ragnarsson-Olding, B.; Platz, A.; Olding, L.; Ringborg, U. p53 protein expression and TP53 mutations in malignant melanomas of sun-sheltered mucosal membranes versus chronically sun-exposed skin. Melanoma Res. 2004, 14, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Prasad, M.L.; Patel, S.G.; Shah, J.P.; Hoshaw-Woodard, S.; Busam, K.J. Prognostic significance of regulators of cell cycle and apoptosis, p16INK4a, p53, and bcl-2 in primary mucosal melanomas of the head and neck. Head Neck Pathol. 2012, 6, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Prasad, M.L.; Patel, S.G.; Huvos, A.G.; Shah, J.P.; Busam, K.J. Primary mucosal melanoma of the head and neck: A proposal for microstaging localized, Stage I (lymph node-negative) tumors. Cancer 2004, 100, 1657–1664. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Li, Y.; Long, Y.; Tang, E.; Wang, R.; Huang, K.; Xie, C.; Chen, G. Increased p16 and p53 protein expression predicts poor prognosis in mucosal melanoma. Oncotarget 2017, 8, 53226–53233. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, S.; Tykodi, S.S.; Thompson, J.A. Treatment of metastatic melanoma: An overview. Oncology 2009, 23, 488–496. [Google Scholar] [PubMed]
- Moll, U.M.; Petrenko, O. The MDM2-p53 interaction. Mol. Cancer Res. 2003, 1, 1001–1008. [Google Scholar] [PubMed]
- Knopf, A.; Plettenberg, C.; Pickhard, A.; Bas, M.; Reifenberger, J.; Bier, H.; Balz, V. Analysis of the functional integrity of the p53 tumor-suppressor gene in malignant melanoma. Melanoma Res. 2011, 21, 380–388. [Google Scholar] [CrossRef] [PubMed]
- Karst, A.M.; Dai, D.L.; Martinka, M.; Li, G. PUMA expression is significantly reduced in human cutaneous melanomas. Oncogene 2005, 24, 1111–1116. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Zhang, L. PUMA: A potent killer with or without p53. Oncogene 2008, 27 (Suppl. 1), S71–S83. [Google Scholar] [CrossRef] [PubMed]
- Amaravadi, R.K. PUMA: A puzzle piece in chloroquine's antimelanoma activity. J. Investig. Dermatol. 2013, 133, 2133–2135. [Google Scholar] [CrossRef] [PubMed]
- Lakhter, A.J.; Sahu, R.P.; Sun, Y.; Kaufmann, W.K.; Androphy, E.J.; Travers, J.B.; Naidu, S.R. Chloroquine promotes apoptosis in melanoma cells by inhibiting BH3 domain-mediated PUMA degradation. J. Investig. Dermatol. 2013, 133, 2247–2254. [Google Scholar] [CrossRef] [PubMed]
- Egger, M.E.; Huang, J.S.; Yin, W.; McMasters, K.M.; McNally, L.R. Inhibition of autophagy with chloroquine is effective in melanoma. J. Surg. Res. 2013, 184, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.Z.; Zhang, J.X.; Zhang, A.L.; Shi, Z.D.; Han, L.; Jia, Z.F.; Yang, W.D.; Wang, G.X.; Jiang, T.; You, Y.P.; et al. MiR-221 and miR-222 target PUMA to induce cell survival in glioblastoma. Mol. Cancer 2010, 9, 229. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Zhang, J.; Zhang, A.; Wang, Y.; Han, L.; You, Y.; Pu, P.; Kang, C. PUMA is a novel target of miR-221/222 in human epithelial cancers. Int. J. Oncol. 2010, 37, 1621–1626. [Google Scholar] [PubMed]
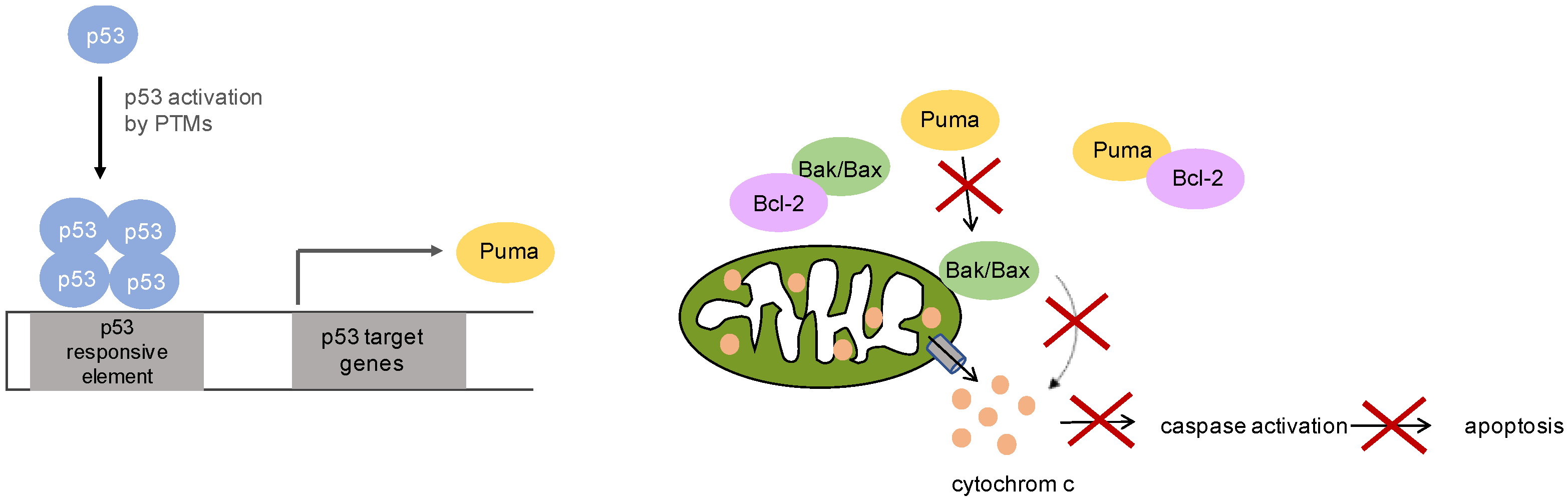
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fritsche, M.K.; Knopf, A. The Tumor Suppressor p53 in Mucosal Melanoma of the Head and Neck. Genes 2017, 8, 384. https://doi.org/10.3390/genes8120384
Fritsche MK, Knopf A. The Tumor Suppressor p53 in Mucosal Melanoma of the Head and Neck. Genes. 2017; 8(12):384. https://doi.org/10.3390/genes8120384
Chicago/Turabian StyleFritsche, Marie Kristin, and Andreas Knopf. 2017. "The Tumor Suppressor p53 in Mucosal Melanoma of the Head and Neck" Genes 8, no. 12: 384. https://doi.org/10.3390/genes8120384
APA StyleFritsche, M. K., & Knopf, A. (2017). The Tumor Suppressor p53 in Mucosal Melanoma of the Head and Neck. Genes, 8(12), 384. https://doi.org/10.3390/genes8120384