Functional Gene-Set Analysis Does Not Support a Major Role for Synaptic Function in Attention Deficit/Hyperactivity Disorder (ADHD)

Abstract
:1. Introduction
2. Methods
2.1. Sample
2.2. Defining Functional Gene Sets
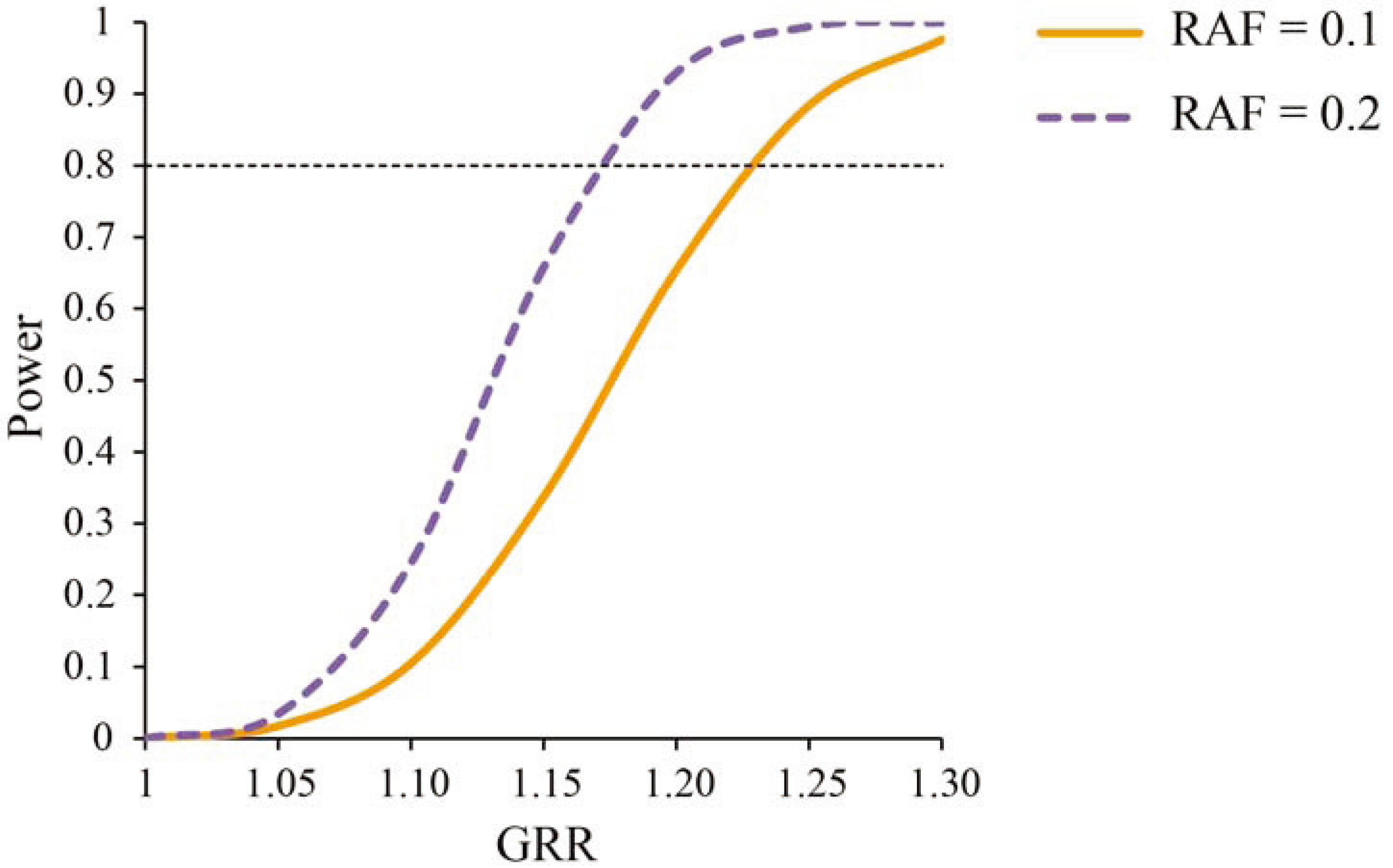
2.3. Power Analysis
2.4. Gene-Set Analysis
3. Results
3.1. Power Analysis
3.2. Gene-Set Analysis

{kind=link}
| Gene Set | Number of Genes in Original Set | Number of Genes Present in GWAS Data | Number of SNPs Present in GWAS Data | Self-Contained p-Value (α = 2.8 × 10−3) | Competitive p-Value (α = 0.05) |
|---|---|---|---|---|---|
| All synaptic genes | 1028 | 956 | 61413 | 0.0393 * | 0.1733 |
| Ion balance/transport | 43 | 40 | 1454 | 0.0118 | NA |
| Cell metabolism | 57 | 51 | 1059 | 0.0429 | NA |
| Endocytosis | 26 | 26 | 1075 | 0.0554 | NA |
| Cell adhesion and trans-synaptic signaling | 81 | 76 | 13550 | 0.0709 | NA |
| Exocytosis | 87 | 83 | 4855 | 0.0962 | NA |
| Protein cluster | 47 | 42 | 4182 | 0.1491 | NA |
| Peptide/neurotrophin signals | 28 | 25 | 1742 | 0.1659 | NA |
| Structural plasticity | 98 | 90 | 4655 | 0.1764 | NA |
| Tyrosine kinase signaling | 7 | 7 | 1281 | 0.2030 | NA |
| Neurotransmitter metabolism | 29 | 27 | 1059 | 0.2959 | NA |
| RNA and protein synthesis, folding and breakdown | 71 | 64 | 1152 | 0.4994 | NA |
| Ligand-gated ion channel signaling | 36 | 32 | 2935 | 0.6500 | NA |
| G-protein-coupled receptor signaling | 41 | 40 | 3129 | 0.6578 | NA |
| Unassigned | 61 | 53 | 2258 | 0.6644 | NA |
| Intracellular signal transduction | 150 | 145 | 9563 | 0.7001 | NA |
| G-protein relay | 27 | 25 | 946 | 0.7047 | NA |
| Intracellular trafficking | 80 | 75 | 2024 | 0.7334 | NA |
| Excitability | 59 | 56 | 4508 | 0.7914 | NA |
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Polanczyk, G.; de Lima, M.S.; Horta, B.L.; Biederman, J.; Rohde, L.A. The worldwide prevalence of ADHD: A systematic review and metaregression analysis. Am. J. Psychiatry 2007, 164, 942–948. [Google Scholar] [PubMed]
- Polanczyk, G.V.; Willcutt, E.G.; Salum, G.A.; Kieling, C.; Rohde, L.A. ADHD prevalence estimates across three decades: An updated systematic review and meta-regression analysis. Int. J. Epidemiol. 2014, 43, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Posthuma, D.; Polderman, T.J.C. What have we learned from recent twin studies about the etiology of neurodevelopmental disorders? Curr. Opin. Neurol. 2013, 26, 111–121. [Google Scholar] [CrossRef]
- Visscher, P.M.; Brown, M.A.; McCarthy, M.I.; Yang, J. Five years of GWAS discovery. Am. J. Hum. Genet. 2012, 90, 7–24. [Google Scholar] [PubMed]
- Neale, B.M.; Medland, S.E.; Ripke, S.; Asherson, P.; Franke, B.; Lesch, K.P.; Faraone, S.V.; Nguyen, T.T.; Schäfer, H.; Holmans, P.; et al. Meta-analysis of genome-wide association studies of attention-deficit/hyperactivity disorder. J. Am. Acad. Child Adolesc. Psychiatry 2010, 49, 884–897. [Google Scholar] [CrossRef] [PubMed]
- Lesch, K.P.; Timmesfeld, N.; Renner, T.J.; Halperin, R.; Röser, C.; Nguyen, T.T.; Craig, D.W.; Romanos, J.; Heine, M.; Meyer, J.; et al. Molecular genetics of adult ADHD: Converging evidence from genome-wide association and extended pedigree linkage studies. J. Neural Transm. 2008, 115, 1573–1585. [Google Scholar] [CrossRef] [PubMed]
- Franke, B.; Neale, B.M.; Faraone, S.V. Genome-wide association studies in ADHD. Hum. Genet. 2009, 126, 13–50. [Google Scholar] [CrossRef] [PubMed]
- Poelmans, G.; Pauls, D.L.; Buitelaar, J.K.; Franke, B. Integrated genome-wide association study findings: Identification of a neurodevelopmental network for attention deficit hyperactivity disorder. Am. J. Psychiatry 2011, 168, 365–377. [Google Scholar] [CrossRef] [PubMed]
- Gizer, I.R.; Ficks, C.; Waldman, I.D. Candidate gene studies of ADHD: A meta-analytic review. Hum. Genet. 2009, 126, 51–90. [Google Scholar] [CrossRef] [PubMed]
- Williams, N.M.; Zaharieva, I.; Martin, A.; Langley, K.; Mantripragada, K.; Fossdal, R.; Stefansson, H.; Stefansson, K.; Magnusson, P.; Gudmundsson, O.O.; et al. Rare chromosomal deletions and duplications in attention-deficit hyperactivity disorder: A genome-wide analysis. Lancet 2010, 376, 1401–1408. [Google Scholar] [CrossRef] [PubMed]
- Elia, J.; Glessner, J.T.; Wang, K.; Takahashi, N.; Shtir, C.J.; Hadley, D.; Sleiman, P.M.; Zhang, H.; Kim, C.E.; Robison, R.; et al. Genome-wide copy number variation study associates metabotropic glutamate receptor gene networks with attention deficit hyperactivity disorder. Nat. Genet. 2012, 44, 78–84. [Google Scholar]
- Stergiakouli, E.; Hamshere, M.; Holmans, P.; Langley, K.; Zaharieva, I.; Hawi, Z.; Kent, L.; Gill, M.; Williams, N.; Owen, M.J.; et al. Investigating the contribution of common genetic variants to the risk and pathogenesis of ADHD. Am. J. Psychiatry 2012, 169, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Williams, N.M.; Franke, B.; Mick, E.; Anney, R.J.L.; Freitag, C.M.; Gill, M.; Thapar, A.; O’Donovan, M.C.; Owen, M.J.; Holmans, P.; et al. Genome-wide analysis of copy number variants in attention deficit hyperactivity disorder: The role of rare variants and duplications at 15q13.3. Am. J. Psychiatry 2012, 169, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Schadt, E.E. Molecular networks as sensors and drivers of common human diseases. Nature 2009, 461, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Ruano, D.; Abecasis, G.R.; Glaser, B.; Lips, E.S.; Cornelisse, L.N.; de Jong, A.P.H.; Evans, D.M.; Davey Smith, G.; Timpson, N.J.; Smit, A.B.; et al. Functional gene group analysis reveals a role of synaptic heterotrimeric G proteins in cognitive ability. Am. J. Hum. Genet. 2010, 86, 113–125. [Google Scholar] [CrossRef] [PubMed]
- De Jong, A.P.H.; Verhage, M. Presynaptic signal transduction pathways that modulate synaptic transmission. Curr. Opin. Neurobiol. 2009, 19, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Feldman, I.; Rzhetsky, A.; Vitkup, D. Network properties of genes harboring inherited disease mutations. Proc. Natl. Acad. Sci. USA 2008, 105, 4323–4328. [Google Scholar] [CrossRef] [PubMed]
- Rossin, E.J.; Lage, K.; Raychaudhuri, S.; Xavier, R.J.; Tatar, D.; Benita, Y.; International Inflammatory Bowel Disease Genetics Constortium; Cotsapas, C.; Daly, M.J. Proteins encoded in genomic regions associated with immune-mediated disease physically interact and suggest underlying biology. PLoS Genet. 2011, 7, e1001273. [Google Scholar] [CrossRef] [PubMed]
- Ripke, S.; O’Dushlaine, C.; Chambert, K.; Moran, J.L.; Kähler, A.K.; Akterin, S.; Bergen, S.E.; Collins, A.L.; Crowley, J.J.; Fromer, M.; et al. Genome-wide association analysis identifies 13 new risk loci for schizophrenia. Nat. Genet. 2013, 45, 1150–1159. [Google Scholar] [CrossRef] [PubMed]
- Goudriaan, A.; de Leeuw, C.; Ripke, S.; Hultman, C.M.; Sklar, P.; Sullivan, P.F.; Smit, A.B.; Posthuma, D.; Verheijen, M.H.G. Specific glial functions contribute to schizophrenia susceptibility. Schizophr. Bull. 2014, 40, 925–935. [Google Scholar] [CrossRef] [PubMed]
- Complex Trait Genetics. Available online: http://ctglab.nl/software/ (accessed on 4 April 2014).
- Purcell, S.; Cherny, S.S.; Sham, P.C. Genetic power calculator: Design of linkage and association genetic mapping studies of complex traits. Bioinformatics 2003, 19, 149–150. [Google Scholar] [CrossRef] [PubMed]
- Purcell, S.; Sham, P.C. Genetic Power Calculator. Available online: http://pngu.mgh.harvard.edu/~purcell/gpc/ (accessed on 3 March 2014).
- Lips, E.S.; Cornelisse, L.N.; Toonen, R.F.; Min, J.L.; Hultman, C.M.; International Scizophrenia Consortium; Holmans, P.A.; O’Donovan, M.C.; Purcell, S.M.; Smit, A.B.; et al. Functional gene group analysis identifies synaptic gene groups as risk factor for schizophrenia. Mol. Psychiatry 2012, 17, 996–1006. [Google Scholar] [CrossRef] [PubMed]
- 1000 Genomes Project Consortium; Abecasis, G.R.; Auton, A.; Brooks, L.D.; DePristo, M.A.; Durbin, R.M.; Handsaker, R.E.; Kang, H.M.; Marth, G.T.; McVean, G.A. An integrated map of genetic variation from 1092 human genomes. Nature 2012, 491, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Bralten, J.; Franke, B.; Waldman, I.; Rommelse, N.; Hartman, C.; Asherson, P.; Banaschewski, T.; Ebstein, R.P.; Gill, M.; Miranda, A.; et al. Candidate genetic pathways for attention-deficit/hyperactivity disorder (ADHD) show association to hyperactive/impulsive symptoms in children with ADHD. J. Am. Acad. Child Adolesc. Psychiatry 2013, 52, 1204.e1–1212.e1. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, M.A.R.; O’Donovan, M.C.; Meng, Y.A.; Jones, I.R.; Ruderfer, D.M.; Jones, L.; Fan, J.; Kirov, G.; Perlis, R.H.; Green, E.K.; et al. Collaborative genome-wide association analysis supports a role for ANK3 and CACNA1C in bipolar disorder. Nat. Genet. 2008, 40, 1056–1058. [Google Scholar] [PubMed]
- Psychiatric GWAS Consortium Bipolar Disorder Working Group. Large-scale genome-wide association analysis of bipolar disorder identifies a new susceptibility locus near ODZ4. Nat. Genet. 2011, 43, 977–983. [Google Scholar]
- Cross-Disorder Group of the Psychiatric Genomics Consortium. Identification of risk loci with shared effects on five major psychiatric disorders: A genome-wide analysis. Lancet 2013, 381, 1371–1379. [Google Scholar]
- Cross-Disorder Group of the Psychiatric Genomics Consortium; Lee, S.H.; Ripke, S.; Neale, B.M.; Faraone, S.V.; Purcell, S.M.; Perlis, R.H.; Mowry, B.J.; Thapar, A.; Goddard, M.E.; et al. Genetic relationship between five psychiatric disorders estimated from genome-wide SNPs. Nat. Genet. 2013, 45, 984–994. [Google Scholar] [PubMed]
- Faraone, S.V. Genetics of childhood disorders: XX. ADHD, Part 4: is ADHD genetically heterogeneous? J. Am. Acad. Child Adolesc. Psychiatry 2000, 39, 1455–1457. [Google Scholar] [CrossRef] [PubMed]
- Psychiatric Genomics Consortium. Available online: https://pgc.unc.edu/Sharing.php#SharingOpp/ (accessed on 23 October 2013).
- Genetic Cluster Computer. Available online: http://www.geneticcluster.org/ (accessed on 4 April 2014).
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Hammerschlag, A.R.; Polderman, T.J.C.; De Leeuw, C.; Tiemeier, H.; White, T.; Smit, A.B.; Verhage, M.; Posthuma, D. Functional Gene-Set Analysis Does Not Support a Major Role for Synaptic Function in Attention Deficit/Hyperactivity Disorder (ADHD). Genes 2014, 5, 604-614. https://doi.org/10.3390/genes5030604
Hammerschlag AR, Polderman TJC, De Leeuw C, Tiemeier H, White T, Smit AB, Verhage M, Posthuma D. Functional Gene-Set Analysis Does Not Support a Major Role for Synaptic Function in Attention Deficit/Hyperactivity Disorder (ADHD). Genes. 2014; 5(3):604-614. https://doi.org/10.3390/genes5030604
Chicago/Turabian StyleHammerschlag, Anke R., Tinca J. C. Polderman, Christiaan De Leeuw, Henning Tiemeier, Tonya White, August B. Smit, Matthijs Verhage, and Danielle Posthuma. 2014. "Functional Gene-Set Analysis Does Not Support a Major Role for Synaptic Function in Attention Deficit/Hyperactivity Disorder (ADHD)" Genes 5, no. 3: 604-614. https://doi.org/10.3390/genes5030604
APA StyleHammerschlag, A. R., Polderman, T. J. C., De Leeuw, C., Tiemeier, H., White, T., Smit, A. B., Verhage, M., & Posthuma, D. (2014). Functional Gene-Set Analysis Does Not Support a Major Role for Synaptic Function in Attention Deficit/Hyperactivity Disorder (ADHD). Genes, 5(3), 604-614. https://doi.org/10.3390/genes5030604