


Isolation and Activity Evaluation of Callus-Specific Promoters in Rice (Oryza sativa L.)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. RNA Isolation and Gene Expression Analysis
2.3. Promoter Cloning, Vector Construction, and Sequence Analysis
2.4. Generation of Transgenic Rice Plants
2.5. DNA Extraction and Identification of T1 Generation Transgenic Rice Plants
2.6. Histochemical Staining
2.7. Quantification of GUS Activity
3. Results
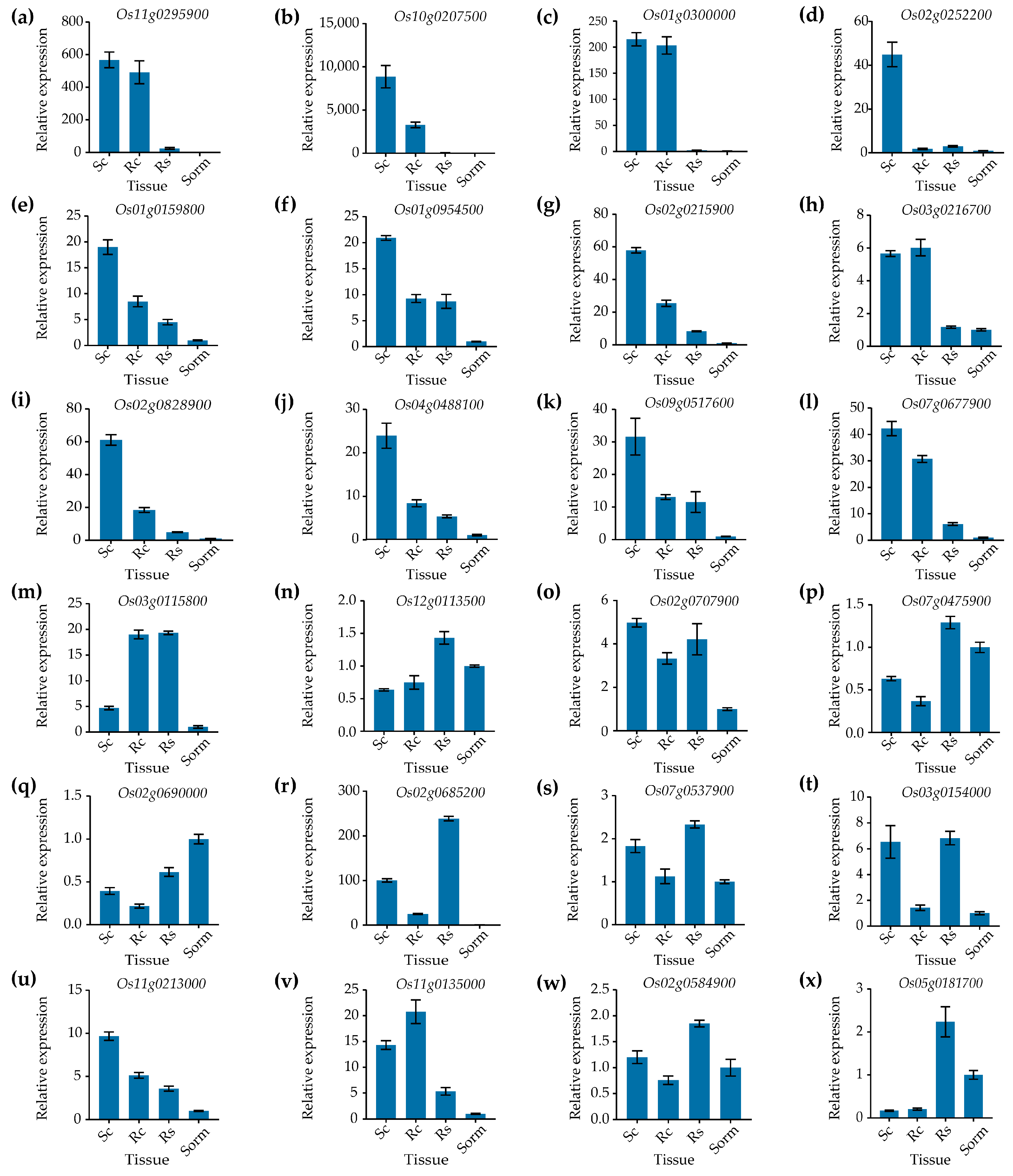
3.1. Identification of Genes Specifically Expressed in Rice Calli
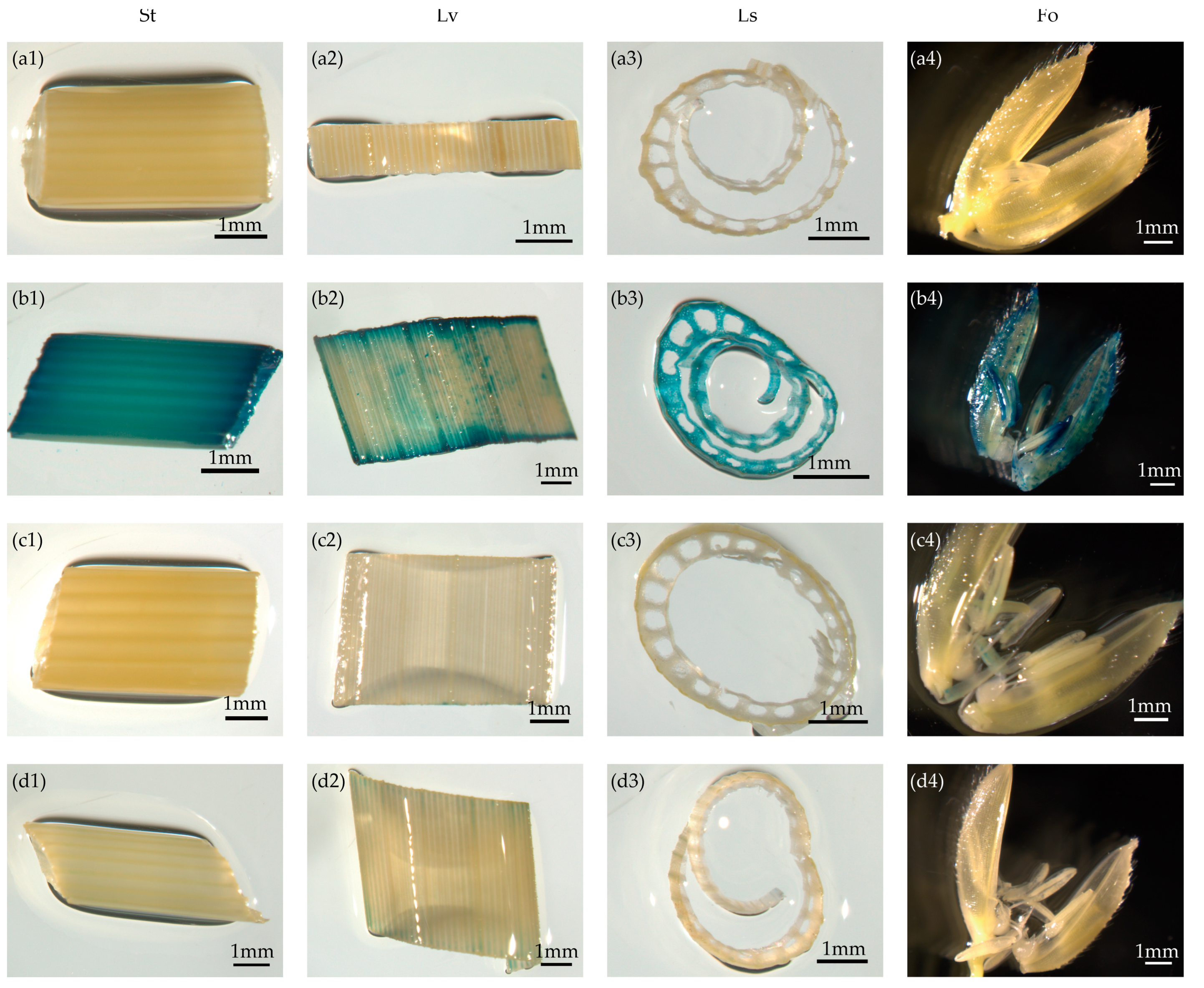
3.2. The Genetic Validation of Promoter Activities
3.3. Quantitative GUS Activity in pOsTDL1B:GUS Transgenic Rice
3.4. Analysis of Cis-Acting Elements in the pOsTDL1B Promoter
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| qPCR | quantitative real-time polymerase chain reaction |
| MTF | morphogenic transcription factor |
| CTAB | cetyltrimethylammonium bromide |
| 4-MU | 4-methylumbelliferone |
References
- Muthayya, S.; Sugimoto, J.D.; Montgomery, S.; Maberly, G.F. An overview of global rice production, supply, trade, and consumption. Ann. N. Y. Acad. Sci. 2014, 1324, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.H.; Cao, L.Y.; Zhuang, J.Y.; Chen, S.G.; Zhan, X.D.; Fan, Y.Y.; Zhu, D.F.; Min, S.K. Super hybrid rice breeding in China: Achievements and prospects. J. Integr. Plant Biol. 2007, 49, 805–810. [Google Scholar] [CrossRef]
- Yang, S.H.; Cheng, B.Y.; Wu, J.L.; Shen, W.F.; Cheng, S.H. Review and prospects on rice breeding and extension in China. Rice Sci. 2006, 13, 1–8. [Google Scholar]
- Chen, Z.; Debernardi, J.M.; Dubcovsky, J.; Gallavotti, A. Recent advances in crop transformation technologies. Nat. Plants 2022, 8, 1343–1351. [Google Scholar] [CrossRef]
- Li, B.S.; Sun, C.; Li, J.Y.; Gao, C.X. Targeted genome-modification tools and their advanced applications in crop breeding. Nat. Rev. Genet. 2024, 25, 603–622. [Google Scholar] [CrossRef]
- Gasser, C.S.; Fraley, R.T. Genetically engineering plants for crop improvement. Science 1989, 244, 1293–1299. [Google Scholar] [CrossRef]
- Wang, F.J.; Wang, C.L.; Liu, P.Q.; Lei, C.L.; Hao, W.; Gao, Y.; Liu, Y.G.; Zhao, K.J. Enhanced rice blast resistance by CRISPR/Cas9-targeted mutagenesis of the ERF transcription factor gene OsERF922. PLoS ONE 2016, 11, e0154027. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Mao, B.; Li, Y.; Lv, Q.; Zhang, L.; Chen, C.; He, H.; Wang, W.; Zeng, X.; Shao, Y.; et al. Knockout of OsNramp5 using the CRISPR/Cas9 system produces low Cd-accumulating indica rice without compromising yield. Sci. Rep. 2017, 7, 14438. [Google Scholar] [CrossRef]
- Xu, R.F.; Yang, Y.C.; Qin, R.Y.; Li, H.; Qiu, C.H.; Li, L.; Wei, P.C.; Yang, J.B. Rapid improvement of grain weight via highly efficient CRISPR/Cas9-mediated multiplex genome editing in rice. J. Genet. Genom. 2016, 43, 529–532. [Google Scholar] [CrossRef]
- Ma, X.L.; Zhang, Q.Y.; Zhu, Q.L.; Liu, W.; Chen, Y.; Qiu, R.; Wang, B.; Yang, Z.F.; Li, H.Y.; Lin, Y.R.; et al. A robust CRISPR/Cas9 system for convenient, high-efficiency multiplex genome editing in monocot and dicot plants. Mol. Plant 2015, 8, 1274–1284. [Google Scholar] [CrossRef]
- Ye, X.; Al-Babili, S.; Klöti, A.; Zhang, J.; Lucca, P.; Beyer, P.; Potrykus, I. Engineering the provitamin A (beta-carotene) biosynthetic pathway into (carotenoid-free) rice endosperm. Science 2000, 287, 303–305. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.M.; Zhang, C.Y.; Yao, B.; Xue, G.X.; Yang, W.Z.; Zhou, X.J.; Zhang, J.M.; Sun, C.; Chen, P.; Fan, Y.L. Corn seeds as bioreactors for the production of phytase in the feed industry. J. Biotechnol. 2013, 165, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.X. Genome engineering for crop improvement and future agriculture. Cell 2021, 184, 1621–1635. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.W.; Lin, Q.Q.; Jin, S.; Gao, C.X. The CRISPR-Cas toolbox and gene editing technologies. Mol. Cell 2022, 82, 333–347. [Google Scholar] [CrossRef]
- Altpeter, F.; Springer, N.M.; Bartley, L.E.; Blechl, A.E.; Brutnell, T.P.; Citovsky, V.; Conrad, L.J.; Gelvin, S.B.; Jackson, D.P.; Kausch, A.P.; et al. Advancing crop transformation in the era of genome editing. Plant Cell 2016, 28, 1510–1520. [Google Scholar] [CrossRef]
- Rashid, H.; Yokoi, S.; Toriyama, K.; Hinata, K. Transgenic plant production mediated by Agrobacterium in Indica rice. Plant Cell Rep. 1996, 15, 727–730. [Google Scholar] [CrossRef]
- Cheng, M.; Fry, J.E.; Pang, S.; Zhou, H.; Hironaka, C.M.; Duncan, D.R.; Conner, T.W.; Wan, Y. Genetic transformation of wheat mediated by Agrobacterium tumefaciens. Plant Physiol. 1997, 115, 971–980. [Google Scholar] [CrossRef]
- Ishida, Y.; Saito, H.; Ohta, S.; Hiei, Y.; Komari, T.; Kumashiro, T. High efficiency transformation of maize (Zea mays L.) mediated by Agrobacterium tumefaciens. Nat. Biotechnol. 1996, 14, 745–750. [Google Scholar] [CrossRef]
- Zhao, Z.Y.; Cai, T.; Tagliani, L.; Miller, M.; Wang, N.; Pang, H.; Rudert, M.; Schroeder, S.; Hondred, D.; Seltzer, J.; et al. Agrobacterium-mediated sorghum transformation. Plant Mol. Biol. 2000, 44, 789–798. [Google Scholar] [CrossRef]
- Anami, S.; Njuguna, E.; Coussens, G.; Aesaert, S.; Van Lijsebettens, M. Higher plant transformation: Principles and molecular tools. Int. J. Dev. Biol. 2013, 57, 483–494. [Google Scholar] [CrossRef]
- Liu, J.; Nannas, N.J.; Fu, F.-F.; Shi, J.; Aspinwall, B.; Parrott, W.A.; Dawe, R.K. Genome-scale sequence disruption following biolistic transformation in rice and maize. Plant Cell 2019, 31, 368–383. [Google Scholar] [CrossRef] [PubMed]
- Taylor, N.J.; Fauquet, C.M. Microparticle bombardment as a tool in plant science and agricultural biotechnology. DNA Cell Biol. 2002, 21, 963–977. [Google Scholar] [CrossRef]
- Hamilton, C.M.; Frary, A.; Lewis, C.; Tanksley, S.D. Stable transfer of intact high molecular weight DNA into plant chromosomes. Proc. Natl. Acad. Sci. USA. 1996, 93, 9975–9979. [Google Scholar] [CrossRef] [PubMed]
- Shrawat, A.K.; Lörz, H. Agrobacterium-mediated transformation of cereals: A promising approach crossing barriers. Plant Biotechnol. J. 2006, 4, 575–603. [Google Scholar] [CrossRef]
- Ji, Q.; Xu, X.; Wang, K. Genetic transformation of major cereal crops. Int. J. Dev. Biol. 2013, 57, 495–508. [Google Scholar] [CrossRef] [PubMed]
- Brettell, R.I.S.; Wernicke, W.; Thomas, E. Embryogenesis from cultured immature inflorescences of Sorghum bicolor. Protoplasm 1980, 104, 141–148. [Google Scholar] [CrossRef]
- Wen, F.S.; Sorensen, E.L.; Barnett, F.L.; Liang, G.H. Callus induction and plant regeneration from anther and inflorescence culture of Sorghum. Euphytica 1991, 52, 177–181. [Google Scholar] [CrossRef]
- Gless, C.; Lörz, H.; Jähne-Gärtner, A. Establishment of a highly efficient regeneration system from leaf base segments of oat (Avena sativa L.). Plant Cell Rep. 1998, 17, 441–445. [Google Scholar] [CrossRef]
- Yu, H.M.; Wang, W.C.; Wang, Y.W.; Hou, B.K. High frequency wheat regeneration from leaf tissue explants of regenerated plantlets. Adv. Biosci. Biotechnol. 2012, 3, 46–50. [Google Scholar] [CrossRef]
- Park, S.H.; Pinson, S.R.M.; Smith, R.H. T-DNA integration into genomic DNA of rice following Agrobacterium inoculation of isolated shoot apices. Plant Mol. Biol 1996, 32, 1135–1148. [Google Scholar] [CrossRef]
- Gordon-Kamm, B.; Sardesai, N.; Arling, M.; Lowe, K.; Hoerster, G.; Betts, S.; Jones, T. Using morphogenic genes to improve recovery and regeneration of transgenic plants. Plants 2019, 8, 38. [Google Scholar] [CrossRef] [PubMed]
- Lowe, K.; Wu, E.; Wang, N.; Hoerster, G.; Hastings, C.; Cho, M.-J.; Scelonge, C.; Lenderts, B.; Chamberlin, M.; Cushatt, J.; et al. Morphogenic regulators Baby boom and Wuschel improve monocot transformation. Plant Cell 2016, 28, 1998–2015. [Google Scholar] [CrossRef] [PubMed]
- Boutilier, K.; Offringa, R.; Sharma, V.K.; Kieft, H.; Ouellet, T.; Zhang, L.; Hattori, J.; Liu, C.-M.; van Lammeren, A.A.M.; Miki, B.L.A.; et al. Ectopic expression of BABY BOOM triggers a conversion from vegetative to embryonic growth. Plant Cell 2002, 14, 1737–1749. [Google Scholar] [CrossRef]
- Wang, K.; Shi, L.; Liang, X.N.; Zhao, P.; Wang, W.X.; Liu, J.X.; Chang, Y.N.; Hiei, Y.; Yanagihara, C.; Du, L.P.; et al. The gene TaWOX5 overcomes genotype dependency in wheat genetic transformation. Nat. Plants 2022, 8, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Zuo, J.; Niu, Q.-W.; Frugis, G.; Chua, N.-H. The WUSCHEL gene promotes vegetative-to-embryonic transition in Arabidopsis. Plant J. 2002, 30, 349–359. [Google Scholar] [CrossRef]
- Aregawi, K.; Shen, J.; Pierroz, G.; Sharma, M.K.; Dahlberg, J.; Owiti, J.; Lemaux, P.G. Morphogene-assisted transformation of Sorghum bicolor allows more efficient genome editing. Plant Biotechnol. J. 2022, 20, 748–760. [Google Scholar] [CrossRef]
- Luo, W.N.; Tan, J.T.; Li, T.; Feng, Z.T.; Ding, Z.; Xie, X.R.; Chen, Y.L.; Chen, L.T.; Liu, Y.G.; Zhu, Q.L.; et al. Overexpression of maize GOLDEN2 in rice and maize calli improves regeneration by activating chloroplast development. Sci. China Life Sci. 2023, 66, 340–349. [Google Scholar] [CrossRef]
- Battraw, M.J.; Hall, T.C. Histochemical analysis of CaMV 35S promoter-β-glucuronidase gene expression in transgenic rice plants. Plant Mol. Biol. 1990, 15, 527–538. [Google Scholar] [CrossRef]
- McElroy, D.; Blowers, A.D.; Jenes, B.; Wu, R. Construction of expression vectors based on the rice actin 1 (Act1) 5′ region for use in monocot transformation. Mol. Gen. Genet. 1991, 231, 150–160. [Google Scholar] [CrossRef]
- Christensen, A.H.; Sharrock, R.A.; Quail, P.H. Maize polyubiquitin genes: Structure, thermal perturbation of expression and transcript splicing, and promoter activity following transfer to protoplasts by electroporation. Plant Mol. Biol. 1992, 18, 675–689. [Google Scholar] [CrossRef]
- Wang, Q.; Zhu, Y.; Sun, L.; Li, L.B.; Jin, S.X.; Zhang, X.L. Transgenic Bt cotton driven by the green tissue-specific promoter shows strong toxicity to lepidopteran pests and lower Bt toxin accumulation in seeds. Sci. China Life Sci. 2016, 59, 172–182. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.Q.; Tian, J.; Zhou, X.J.; Chen, R.M.; Wang, L.; Zhang, C.Y.; Zhao, J.; Fan, Y.L. Identification and characterization of promoters specifically and strongly expressed in maize embryos. Plant Biotechnol. J. 2014, 12, 1286–1296. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.-Y.; Adachi, T.; Hatano, T.; Washida, H.; Suzuki, A.; Takaiwa, F. Promoters of rice seed storage protein genes direct endosperm-specific gene expression in transgenic rice. Plant Cell Physiol. 1998, 39, 885–889. [Google Scholar] [CrossRef]
- Zhu, Q.L.; Yu, S.Z.; Zeng, D.C.; Liu, H.M.; Wang, H.C.; Yang, Z.F.; Xie, X.R.; Shen, R.X.; Tan, J.T.; Li, H.Y.; et al. Development of “purple endosperm rice” by engineering anthocyanin biosynthesis in the endosperm with a high-efficiency transgene stacking system. Mol. Plant 2017, 10, 918–929. [Google Scholar] [CrossRef]
- Liu, J.-J.; Ekramoddoullah, A.K.M. Root-specific expression of a western white pine PR10 gene is mediated by different promoter regions in transgenic tobacco. Plant Mol. Biol. 2003, 52, 103–120. [Google Scholar] [CrossRef]
- Huang, N.; Wu, L.; Nandi, S.; Bowman, E.; Huang, J.; Sutliff, T.; Rodriguez, R.L. The tissue-specific activity of a rice beta-glucanase promoter (Gns9) is used to select rice transformants. Plant Sci. 2001, 161, 589–595. [Google Scholar] [CrossRef]
- Hiei, Y.; Ohta, S.; Komari, T.; Kumashiro, T. Efficient transformation of rice (Oryza sativa L.) mediated by Agrobacterium and sequence analysis of the boundaries of the T-DNA. Plant J. 1994, 6, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4325. [Google Scholar] [CrossRef]
- Jefferson, R.A.; Kavanagh, T.A.; Bevan, M.W. GUS fusions: β-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 1987, 6, 3901–3907. [Google Scholar] [CrossRef]
- Wu, C.-Y.; Suzuki, A.; Washida, H.; Takaiwa, F. The GCN4 motif in a rice glutelin gene is essential for endosperm-specific gene expression and is activated by Opaque-2 in transgenic rice plants. Plant J. 1998, 14, 673–683. [Google Scholar] [CrossRef] [PubMed]
- Yin, T.; Wu, H.Y.; Zhang, S.L.; Liu, J.M.; Lu, H.Y.; Zhang, L.X.; Xu, Y.; Chen, D.M. Two negative cis-regulatory regions involved in fruit-specific promoter activity from watermelon (Citrullus vulgaris S.). J. Exp. Bot. 2009, 60, 169–185. [Google Scholar] [CrossRef]
- Cai, M.; Wei, J.; Li, X.H.; Xu, C.G.; Wang, S.P. A rice promoter containing both novel positive and negative cis-elements for regulation of green tissue-specific gene expression in transgenic plants. Plant Biotechnol. J. 2007, 5, 664–674. [Google Scholar] [CrossRef]
- Ye, R.J.; Zhou, F.; Lin, Y.J. Two novel positive cis-regulatory elements involved in green tissue-specific promoter activity in rice (Oryza sativa L ssp.). Plant Cell Rep. 2012, 31, 1159–1172. [Google Scholar] [CrossRef]
- Qu, L.Q.; Xing, Y.P.; Liu, W.X.; Xu, X.P.; Song, Y.R. Expression pattern and activity of six glutelin gene promoters in transgenic rice. J. Exp. Bot. 2008, 59, 2417–2424. [Google Scholar] [CrossRef]
- Li, H.; Li, J.; Xu, R.F.; Qin, R.Y.; Song, F.S.; Li, L.; Wei, P.C.; Yang, J.B. Isolation of five rice nonendosperm tissue-expressed promoters and evaluation of their activities in transgenic rice. Plant Biotechnol. J. 2018, 16, 1138–1147. [Google Scholar] [CrossRef] [PubMed]
- De Wilde, C.; Van Houdt, H.; De Buck, S.; Angenon, G.; De Jaeger, G.; Depicker, A. Plants as bioreactors for protein production: Avoiding the problem of transgene silencing. Plant Mol. Biol. 2000, 43, 347–359. [Google Scholar] [CrossRef] [PubMed]
- Lowe, K.; La Rota, M.; Hoerster, G.; Hastings, C.; Wang, N.; Chamberlin, M.; Wu, E.; Jones, T.; Gordon-Kamm, W. Rapid genotype “independent” Zea mays L. (maize) transformation via direct somatic embryogenesis. In Vitro Cell. Dev. Biol. Plant 2018, 54, 240–252. [Google Scholar] [CrossRef]
- Wang, N.; Arling, M.; Hoerster, G.; Ryan, L.; Wu, E.; Lowe, K.; Gordon-Kamm, W.; Jones, T.J.; Chilcoat, N.D.; Anand, A. An efficient gene excision system in maize. Front. Plant Sci. 2020, 11, 1298. [Google Scholar] [CrossRef]
- Li, J.P.; Pan, W.B.; Zhang, S.; Ma, G.J.; Li, A.X.; Zhang, H.W.; Liu, L.J. A rapid and highly efficient sorghum transformation strategy using GRF4-GIF1/ternary vector system. Plant J. 2024, 117, 1604–1613. [Google Scholar] [CrossRef]
- Belaffif, M.B.; Brown, M.C.; Marcial, B.; Baysal, C.; Swaminathan, K. New strategies to advance plant transformation. Curr. Opin. Biotechnol. 2025, 91, 103241. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Luo, K.; Li, Z.; Yang, Y. A novel method for induction of plant regeneration via somatic embryogenesis. Plant Sci. 2009, 177, 43–48. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, Y.; Lu, M.-H.; Chai, Y.-P.; Jiang, Y.-Y.; Zhou, Y.; Wang, X.-C.; Chen, Q.-J. A novel ternary vector system united with morphogenic genes enhances CRISPR/Cas delivery in maize. Plant Physiol. 2019, 181, 1441–1448. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, X.; Wu, C. Isolation and Activity Evaluation of Callus-Specific Promoters in Rice (Oryza sativa L.). Genes 2025, 16, 610. https://doi.org/10.3390/genes16050610
Ma X, Wu C. Isolation and Activity Evaluation of Callus-Specific Promoters in Rice (Oryza sativa L.). Genes. 2025; 16(5):610. https://doi.org/10.3390/genes16050610
Chicago/Turabian StyleMa, Xiaojiao, and Chuanyin Wu. 2025. "Isolation and Activity Evaluation of Callus-Specific Promoters in Rice (Oryza sativa L.)" Genes 16, no. 5: 610. https://doi.org/10.3390/genes16050610
APA StyleMa, X., & Wu, C. (2025). Isolation and Activity Evaluation of Callus-Specific Promoters in Rice (Oryza sativa L.). Genes, 16(5), 610. https://doi.org/10.3390/genes16050610