
Gene–Diet Interactions in Diabetes Mellitus: Current Insights and the Potential of Personalized Nutrition

 , , , and
, , , and 
Abstract
1. Introduction
2. Genetic Basis of T2DM
3. Gene–Diet Interactions in T2DM
4. Benefits of G × D Interactions in T2DM
4.1. Identification of High-Risk Individuals
4.2. Motivation and Adherence
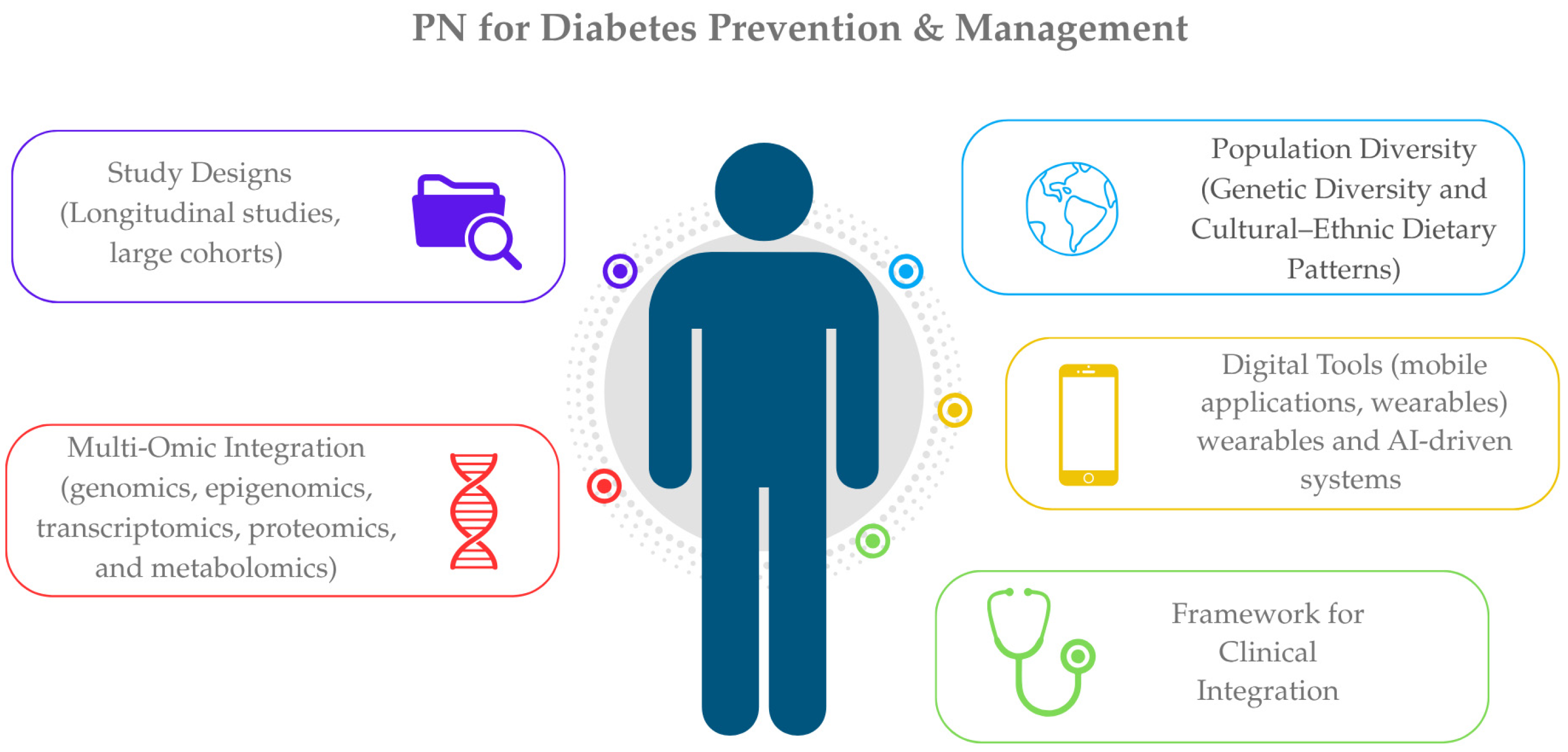
5. Research Gaps and Future Directions
5.1. Study Designs
5.2. Population Diversity
5.3. Multi-Omic Integration
5.4. Digital Tools and PN for T2DM
5.5. Framework for Clinical Integration
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Dilworth, L.; Facey, A.; Omoruyi, F. Diabetes Mellitus and Its Metabolic Complications: The Role of Adipose Tissues. Int. J. Mol. Sci. 2021, 22, 7644. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Colclough, K.; Gloyn, A.L.; Pollin, T.I. Monogenic diabetes: A gateway to precision medicine in diabetes. J. Clin. Investig. 2021, 131, e142244. [Google Scholar] [CrossRef]
- American Diabetes Association 2. Classification and Diagnosis of Diabetes: Standards of Medical Care in Diabetes-2020. Diabetes Care 2020, 43, S14–S31. [Google Scholar] [CrossRef] [PubMed]
- Riddle, M.C.; Philipson, L.H.; Rich, S.S.; Carlsson, A.; Franks, P.W.; Greeley, S.A.W.; Nolan, J.J.; Pearson, E.R.; Zeitler, P.S.; Hattersley, A.T. Monogenic Diabetes: From Genetic Insights to Population-Based Precision in Care. Reflections from a Diabetes Care Editors’ Expert Forum. Diabetes Care 2020, 43, 3117–3128. [Google Scholar] [CrossRef]
- Silva, A.; Silva, L.; Lopes, I.; Francisco, A.; Neto, A.; Monteiro, M.; Muela, H.; Silva, A.; Silva, L.; Lopes, I.; et al. Association of Dietary Pattern and Physical Inactivity with Hypertension, Obesity, Diabetes and Metabolic Syndrome. In Metabolic Syndrome—And Biological Risk Factors; IntechOpen: London, UK, 2024; ISBN 978-0-85014-433-8. [Google Scholar]
- Kim, M.S.; Shim, I.; Fahed, A.C.; Do, R.; Park, W.-Y.; Natarajan, P.; Khera, A.V.; Won, H.-H. Association of genetic risk, lifestyle, and their interaction with obesity and obesity-related morbidities. Cell Metab. 2024, 36, 1494–1503.e3. [Google Scholar] [CrossRef] [PubMed]
- Cole, J.B.; Florez, J.C. Genetics of diabetes and diabetes complications. Nat. Rev. Nephrol. 2020, 16, 377–390. [Google Scholar] [CrossRef]
- Goyal, S.; Rani, J.; Bhat, M.A.; Vanita, V. Genetics of diabetes. World J. Diabetes 2023, 14, 656–679. [Google Scholar] [CrossRef]
- Laakso, M.; Fernandes Silva, L. Genetics of Type 2 Diabetes: Past, Present, and Future. Nutrients 2022, 14, 3201. [Google Scholar] [CrossRef]
- Sami, W.; Ansari, T.; Butt, N.S.; Hamid, M.R.A. Effect of diet on type 2 diabetes mellitus: A review. Int. J. Health Sci. 2017, 11, 65–71. [Google Scholar]
- Corella, D.; Ordovás, J.M. Biomarkers: Background, classification and guidelines for applications in nutritional epidemiology. Nutr. Hosp. 2015, 31 (Suppl. 3), 177–188. [Google Scholar] [CrossRef]
- Westerman, K.E.; Walker, M.E.; Gaynor, S.M.; Wessel, J.; DiCorpo, D.; Ma, J.; Alonso, A.; Aslibekyan, S.; Baldridge, A.S.; Bertoni, A.G.; et al. Investigating Gene–Diet Interactions Impacting the Association Between Macronutrient Intake and Glycemic Traits. Diabetes 2023, 72, 653–665. [Google Scholar] [CrossRef]
- Ortega, Á.; Berná, G.; Rojas, A.; Martín, F.; Soria, B. Gene-Diet Interactions in Type 2 Diabetes: The Chicken and Egg Debate. Int. J. Mol. Sci. 2017, 18, 1188. [Google Scholar] [CrossRef]
- Zeevi, D.; Korem, T.; Zmora, N.; Israeli, D.; Rothschild, D.; Weinberger, A.; Ben-Yacov, O.; Lador, D.; Avnit-Sagi, T.; Lotan-Pompan, M.; et al. Personalized Nutrition by Prediction of Glycemic Responses. Cell 2015, 163, 1079–1094. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, S.; Jacobs, S.; Zheng, J.; Meidtner, K.; Schwingshackl, L.; Schulze, M.B. Gene-lifestyle interaction on risk of type 2 diabetes: A systematic review. Obes. Rev. 2019, 20, 1557–1571. [Google Scholar] [CrossRef] [PubMed]
- Almgren, P.; Lehtovirta, M.; Isomaa, B.; Sarelin, L.; Taskinen, M.R.; Lyssenko, V.; Tuomi, T.; Groop, L.; Botnia Study Group. Heritability and familiality of type 2 diabetes and related quantitative traits in the Botnia Study. Diabetologia 2011, 54, 2811–2819. [Google Scholar] [CrossRef] [PubMed]
- Bazzazzadehgan, S.; Shariat-Madar, Z.; Mahdi, F. Distinct Roles of Common Genetic Variants and Their Contributions to Diabetes: MODY and Uncontrolled T2DM. Biomolecules 2025, 15, 414. [Google Scholar] [CrossRef]
- Scott, R.A.; Scott, L.J.; Mägi, R.; Marullo, L.; Gaulton, K.J.; Kaakinen, M.; Pervjakova, N.; Pers, T.H.; Johnson, A.D.; Eicher, J.D.; et al. An Expanded Genome-Wide Association Study of Type 2 Diabetes in Europeans. Diabetes 2017, 66, 2888–2902. [Google Scholar] [CrossRef]
- Dupuis, J.; Langenberg, C.; Prokopenko, I.; Saxena, R.; Soranzo, N.; Jackson, A.U.; Wheeler, E.; Glazer, N.L.; Bouatia-Naji, N.; Gloyn, A.L.; et al. New genetic loci implicated in fasting glucose homeostasis and their impact on type 2 diabetes risk. Nat. Genet. 2010, 42, 105–116. [Google Scholar] [CrossRef]
- DeForest, N.; Majithia, A.R. Genetics of Type 2 Diabetes: Implications from Large-Scale Studies. Curr. Diab Rep. 2022, 22, 227–235. [Google Scholar] [CrossRef]
- Vujkovic, M.; Keaton, J.M.; Lynch, J.A.; Miller, D.R.; Zhou, J.; Tcheandjieu, C.; Huffman, J.E.; Assimes, T.L.; Lorenz, K.; Zhu, X.; et al. Discovery of 318 new risk loci for type 2 diabetes and related vascular outcomes among 1.4 million participants in a multi-ancestry meta-analysis. Nat. Genet. 2020, 52, 680–691. [Google Scholar] [CrossRef]
- Lyssenko, V.; Jonsson, A.; Almgren, P.; Pulizzi, N.; Isomaa, B.; Tuomi, T.; Berglund, G.; Altshuler, D.; Nilsson, P.; Groop, L. Clinical risk factors, DNA variants, and the development of type 2 diabetes. N. Engl. J. Med. 2008, 359, 2220–2232. [Google Scholar] [CrossRef] [PubMed]
- Meigs, J.B.; Shrader, P.; Sullivan, L.M.; McAteer, J.B.; Fox, C.S.; Dupuis, J.; Manning, A.K.; Florez, J.C.; Wilson, P.W.F.; D’Agostino, R.B.; et al. Genotype Score in Addition to Common Risk Factors for Prediction of Type 2 Diabetes. N. Engl. J. Med. 2008, 359, 2208–2219. [Google Scholar] [CrossRef] [PubMed]
- Polfus, L.M.; Darst, B.F.; Highland, H.; Sheng, X.; Ng, M.C.Y.; Below, J.E.; Petty, L.; Bien, S.; Sim, X.; Wang, W.; et al. Genetic discovery and risk characterization in type 2 diabetes across diverse populations. Hum. Genet. Genom. Adv. 2021, 2, 100029. [Google Scholar] [CrossRef] [PubMed]
- Turley, P.; Meyer, M.N.; Wang, N.; Cesarini, D.; Hammonds, E.; Martin, A.R.; Neale, B.M.; Rehm, H.L.; Wilkins-Haug, L.; Benjamin, D.J.; et al. Problems with Using Polygenic Scores to Select Embryos. N. Engl. J. Med. 2021, 385, 78–86. [Google Scholar] [CrossRef]
- Du, Z.; Zhang, F.; Ge, Y.; Liu, Y.; Yu, H.; Wang, Y.; Dalan, R.; Shen, X. Application of Wearable Devices in Diabetes Management. Health Metab. 2025, 2, 7. [Google Scholar] [CrossRef]
- Alemu, R.; Sharew, N.T.; Arsano, Y.Y.; Ahmed, M.; Tekola-Ayele, F.; Mersha, T.B.; Amare, A.T. Multi-omics approaches for understanding gene-environment interactions in noncommunicable diseases: Techniques, translation, and equity issues. Hum. Genom. 2025, 19, 8. [Google Scholar] [CrossRef]
- Farhud, D.; Zarif Yeganeh, M.; Zarif Yeganeh, M. Nutrigenomics and nutrigenetics. Iran. J. Public Health 2010, 39, 1–14. [Google Scholar]
- Merino, J.; Guasch-Ferré, M.; Li, J.; Chung, W.; Hu, Y.; Ma, B.; Li, Y.; Kang, J.H.; Kraft, P.; Liang, L.; et al. Polygenic scores, diet quality, and type 2 diabetes risk: An observational study among 35,759 adults from 3 US cohorts. PLoS Med. 2022, 19, e1003972. [Google Scholar] [CrossRef]
- Karvela, M.; Golden, C.T.; Bell, N.; Martin-Li, S.; Bedzo-Nutakor, J.; Bosnic, N.; DeBeaudrap, P.; de Mateo-Lopez, S.; Alajrami, A.; Qin, Y.; et al. Assessment of the impact of a personalised nutrition intervention in impaired glucose regulation over 26 weeks: A randomised controlled trial. Sci. Rep. 2024, 14, 5428. [Google Scholar] [CrossRef]
- Westerman, K.E.; MEIGS, J.B.; MANNING, A. 1469-P: Gene-Level Gene–Mediterranean Diet Interaction Detection Influencing Inflammatory and Glycemic Biomarkers in 137,000 UK Biobank Participants. Diabetes 2023, 72, 1469-P. [Google Scholar] [CrossRef]
- Yang, R.; Lin, J.; Yang, H.; Dunk, M.M.; Wang, J.; Xu, W.; Wang, Y. A low-inflammatory diet is associated with a lower incidence of diabetes: Role of diabetes-related genetic risk. BMC Med. 2023, 21, 483. [Google Scholar] [CrossRef]
- Apio, C.; Chung, W.; Moon, M.K.; Kwon, O.; Park, T. Gene–diet interaction analysis using novel weighted food scores discovers the adipocytokine signaling pathway associated with the development of type 2 diabetes. Front. Endocrinol. 2023, 14, 1165744. [Google Scholar] [CrossRef] [PubMed]
- Nagel, Z.D.; Engelward, B.P.; Brenner, D.J.; Begley, T.J.; Sobol, R.W.; Bielas, J.H.; Stambrook, P.J.; Wei, Q.; Hu, J.J.; Terry, M.B.; et al. Towards precision prevention: Technologies for identifying healthy individuals with high risk of disease. Mutat. Res. 2017, 800–802, 14–28. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.-L.; Chuang, C.-K.; Chiu, H.-C.; Chang, Y.-H.; Tu, Y.-R.; Lo, Y.-T.; Lin, H.-Y.; Lin, S.-P. Understanding Genetic Screening: Harnessing Health Information to Prevent Disease Risks. Int. J. Med. Sci. 2025, 22, 903–919. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, A.; Taliun, D.; Thurner, M.; Robertson, N.R.; Torres, J.M.; Rayner, N.W.; Payne, A.J.; Steinthorsdottir, V.; Scott, R.A.; Grarup, N.; et al. Fine-mapping type 2 diabetes loci to single-variant resolution using high-density imputation and islet-specific epigenome maps. Nat. Genet. 2018, 50, 1505–1513. [Google Scholar] [CrossRef]
- Zeggini, E.; Scott, L.J.; Saxena, R.; Voight, B.F.; Marchini, J.L.; Hu, T.; de Bakker, P.I.W.; Abecasis, G.R.; Almgren, P.; Andersen, G.; et al. Meta-analysis of genome-wide association data and large-scale replication identifies additional susceptibility loci for type 2 diabetes. Nat. Genet. 2008, 40, 638–645. [Google Scholar] [CrossRef]
- Zhang, C.; Qi, L.; Hunter, D.J.; Meigs, J.B.; Manson, J.E.; van Dam, R.M.; Hu, F.B. Variant of transcription factor 7-like 2 (TCF7L2) gene and the risk of type 2 diabetes in large cohorts of U.S. women and men. Diabetes 2006, 55, 2645–2648. [Google Scholar] [CrossRef]
- Grant, S.F.A.; Thorleifsson, G.; Reynisdottir, I.; Benediktsson, R.; Manolescu, A.; Sainz, J.; Helgason, A.; Stefansson, H.; Emilsson, V.; Helgadottir, A.; et al. Variant of transcription factor 7-like 2 (TCF7L2) gene confers risk of type 2 diabetes. Nat. Genet. 2006, 38, 320–323. [Google Scholar] [CrossRef]
- Dimas, A.S.; Lagou, V.; Barker, A.; Knowles, J.W.; Mägi, R.; Hivert, M.-F.; Benazzo, A.; Rybin, D.; Jackson, A.U.; Stringham, H.M.; et al. Impact of type 2 diabetes susceptibility variants on quantitative glycemic traits reveals mechanistic heterogeneity. Diabetes 2014, 63, 2158–2171. [Google Scholar] [CrossRef]
- Läll, K.; Mägi, R.; Morris, A.; Metspalu, A.; Fischer, K. Personalized risk prediction for type 2 diabetes: The potential of genetic risk scores. Genet. Med. 2017, 19, 322–329. [Google Scholar] [CrossRef]
- Grant, R.W.; O’Brien, K.E.; Waxler, J.L.; Vassy, J.L.; Delahanty, L.M.; Bissett, L.G.; Green, R.C.; Stember, K.G.; Guiducci, C.; Park, E.R.; et al. Personalized Genetic Risk Counseling to Motivate Diabetes Prevention. Diabetes Care 2013, 36, 13–19. [Google Scholar] [CrossRef]
- Hollands, G.J.; French, D.P.; Griffin, S.J.; Prevost, A.T.; Sutton, S.; King, S.; Marteau, T.M. The impact of communicating genetic risks of disease on risk-reducing health behaviour: Systematic review with meta-analysis. BMJ 2016, 352, i1102. [Google Scholar] [CrossRef] [PubMed]
- Jinnette, R.; Narita, A.; Manning, B.; McNaughton, S.A.; Mathers, J.C.; Livingstone, K.M. Does Personalized Nutrition Advice Improve Dietary Intake in Healthy Adults? A Systematic Review of Randomized Controlled Trials. Adv. Nutr. 2020, 12, 657–669. [Google Scholar] [CrossRef] [PubMed]
- Michie, S.; van Stralen, M.M.; West, R. The behaviour change wheel: A new method for characterising and designing behaviour change interventions. Implement. Sci. 2011, 6, 42. [Google Scholar] [CrossRef]
- Driver, M.N.; Kuo, S.I.-C.; Dick, D.M. Returning complex genetic risk information to promote better health-related behaviors: A commentary of the literature and suggested next steps. Transl. Behav. Med. 2022, 13, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Celis-Morales, C.; Livingstone, K.M.; Marsaux, C.F.; Macready, A.L.; Fallaize, R.; O’Donovan, C.B.; Woolhead, C.; Forster, H.; Walsh, M.C.; Navas-Carretero, S.; et al. Effect of personalized nutrition on health-related behaviour change: Evidence from the Food4Me European randomized controlled trial. Int. J. Epidemiol. 2017, 46, 578–588. [Google Scholar] [CrossRef]
- Livingstone, K.M.; Celis-Morales, C.; Navas-Carretero, S.; San-Cristobal, R.; Macready, A.L.; Fallaize, R.; Forster, H.; Woolhead, C.; O’Donovan, C.B.; Marsaux, C.F.; et al. Effect of an Internet-based, personalized nutrition randomized trial on dietary changes associated with the Mediterranean diet: The Food4Me Study. Am. J. Clin. Nutr. 2016, 104, 288–297. [Google Scholar] [CrossRef]
- Nielsen, D.E.; Shih, S.; El-Sohemy, A. Perceptions of genetic testing for personalized nutrition: A randomized trial of DNA-based dietary advice. J. Nutr. Nutr. 2014, 7, 94–104. [Google Scholar] [CrossRef]
- Ryan, R.M.; Deci, E.L. Self-Determination Theory: Basic Psychological Needs in Motivation, Development, and Wellness; The Guilford Press: New York, NY, USA, 2017; Volumes 123–178, p. 756. ISBN 978-1-4625-2876-9. [Google Scholar]
- Horne, J.R.; Nielsen, D.E.; Madill, J.; Robitaille, J.; Vohl, M.-C.; Mutch, D.M. Guiding Global Best Practice in Personalized Nutrition Based on Genetics: The Development of a Nutrigenomics Care Map. J. Acad. Nutr. Diet. 2022, 122, 259–268. [Google Scholar] [CrossRef]
- Savitz, D.A.; Wellenius, G.A. Can Cross-Sectional Studies Contribute to Causal Inference? It Depends. Am. J. Epidemiol. 2023, 192, 514–516. [Google Scholar] [CrossRef]
- Oh, J.; Cha, J.; Choi, S. Identification of Novel Genetic Variants and Food Intake Factors Associated with Type 2 Diabetes in South Korean Adults, Using an Illness–Death Model. Int. J. Mol. Sci. 2025, 26, 2597. [Google Scholar] [CrossRef] [PubMed]
- Barbaresko, J.; Rienks, J.; Nöthlings, U. Lifestyle Indices and Cardiovascular Disease Risk: A Meta-analysis. Am. J. Prev. Med. 2018, 55, 555–564. [Google Scholar] [CrossRef] [PubMed]
- Corella, D.; Coltell, O.; Macian, F.; Ordovás, J.M. Advances in Understanding the Molecular Basis of the Mediterranean Diet Effect. Annu. Rev. Food Sci. Technol. 2018, 9, 227–249. [Google Scholar] [CrossRef]
- Allen, N.E.; Lacey, B.; Lawlor, D.A.; Pell, J.P.; Gallacher, J.; Smeeth, L.; Elliott, P.; Matthews, P.M.; Lyons, R.A.; Whetton, A.D.; et al. Prospective study design and data analysis in UK Biobank. Sci. Transl. Med. 2024, 16, eadf4428. [Google Scholar] [CrossRef] [PubMed]
- Forouhi, N.G.; Wareham, N.J. The EPIC-InterAct Study: A Study of the Interplay between Genetic and Lifestyle Behavioral Factors on the Risk of Type 2 Diabetes in European Populations. Curr. Nutr. Rep. 2014, 3, 355–363. [Google Scholar] [CrossRef]
- Ioannidis, J.P.A. The Challenge of Reforming Nutritional Epidemiologic Research. JAMA 2018, 320, 969–970. [Google Scholar] [CrossRef]
- Touvier, M.; Kesse-Guyot, E.; Méjean, C.; Pollet, C.; Malon, A.; Castetbon, K.; Hercberg, S. Comparison between an interactive web-based self-administered 24 h dietary record and an interview by a dietitian for large-scale epidemiological studies. Br. J. Nutr. 2011, 105, 1055–1064. [Google Scholar] [CrossRef]
- Popejoy, A.B.; Fullerton, S.M. Genomics is failing on diversity. Nature 2016, 538, 161–164. [Google Scholar] [CrossRef]
- Morales, J.; Welter, D.; Bowler, E.H.; Cerezo, M.; Harris, L.W.; McMahon, A.C.; Hall, P.; Junkins, H.A.; Milano, A.; Hastings, E.; et al. A standardized framework for representation of ancestry data in genomics studies, with application to the NHGRI-EBI GWAS Catalog. Genome Biol. 2018, 19, 21. [Google Scholar] [CrossRef]
- Bentley, A.R.; Callier, S.; Rotimi, C.N. Diversity and inclusion in genomic research: Why the uneven progress? J. Community Genet. 2017, 8, 255–266. [Google Scholar] [CrossRef]
- Martin, A.R.; Kanai, M.; Kamatani, Y.; Okada, Y.; Neale, B.M.; Daly, M.J. Clinical use of current polygenic risk scores may exacerbate health disparities. Nat. Genet. 2019, 51, 584–591. [Google Scholar] [CrossRef]
- Wojcik, G.L.; Graff, M.; Nishimura, K.K.; Tao, R.; Haessler, J.; Gignoux, C.R.; Highland, H.M.; Patel, Y.M.; Sorokin, E.P.; Avery, C.L.; et al. Genetic analyses of diverse populations improves discovery for complex traits. Nature 2019, 570, 514–518. [Google Scholar] [CrossRef] [PubMed]
- Fuchsberger, C.; Flannick, J.; Teslovich, T.M.; Mahajan, A.; Agarwala, V.; Gaulton, K.J.; Ma, C.; Fontanillas, P.; Moutsianas, L.; McCarthy, D.J.; et al. The genetic architecture of type 2 diabetes. Nature 2016, 536, 41–47. [Google Scholar] [CrossRef]
- Wuni, R.; Vimaleswaran, K.S. Barriers in Translating Existing Nutrigenetics Insights to Precision Nutrition for Cardiometabolic Health in Ethnically Diverse Populations. Lifestyle Genom. 2024, 17, 122–135. [Google Scholar] [CrossRef]
- H3Africa Consortium; Rotimi, C.; Abayomi, A.; Abimiku, A.; Adabayeri, V.M.; Adebamowo, C.; Adebiyi, E.; Ademola, A.D.; Adeyemo, A.; Adu, D.; et al. Research capacity. Enabling the genomic revolution in Africa. Science 2014, 344, 1346–1348. [Google Scholar] [CrossRef]
- All of Us Research Program Investigators; Denny, J.C.; Rutter, J.L.; Goldstein, D.B.; Philippakis, A.; Smoller, J.W.; Jenkins, G.; Dishman, E. The “All of Us” Research Program. N. Engl. J. Med. 2019, 381, 668–676. [Google Scholar] [CrossRef] [PubMed]
- Sirugo, G.; Williams, S.M.; Tishkoff, S.A. The Missing Diversity in Human Genetic Studies. Cell 2019, 177, 26–31. [Google Scholar] [CrossRef]
- Eisenstein, M. Diversity: Of beans and genes. Nature 2010, 468, S13–S15. [Google Scholar] [CrossRef] [PubMed]
- Perry, G.H.; Dominy, N.J.; Claw, K.G.; Lee, A.S.; Fiegler, H.; Redon, R.; Werner, J.; Villanea, F.A.; Mountain, J.L.; Misra, R.; et al. Diet and the evolution of human amylase gene copy number variation. Nat. Genet. 2007, 39, 1256–1260. [Google Scholar] [CrossRef]
- Kuchay, R.A.H. New insights into the molecular basis of lactase non-persistence/persistence: A brief review. Drug Discov. Ther. 2020, 14, 1–7. [Google Scholar] [CrossRef]
- Troelsen, J.T. Adult-type hypolactasia and regulation of lactase expression. Biochim. Biophys. Acta 2005, 1723, 19–32. [Google Scholar] [CrossRef] [PubMed]
- Itan, Y.; Powell, A.; Beaumont, M.A.; Burger, J.; Thomas, M.G. The Origins of Lactase Persistence in Europe. PLOS Comput. Biol. 2009, 5, e1000491. [Google Scholar] [CrossRef] [PubMed]
- Labrie, V.; Buske, O.J.; Oh, E.; Jeremian, R.; Ptak, C.; Gasiūnas, G.; Maleckas, A.; Petereit, R.; Žvirbliene, A.; Adamonis, K.; et al. Lactase nonpersistence is directed by DNA-variation-dependent epigenetic aging. Nat. Struct. Mol. Biol. 2016, 23, 566–573. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Ye, K. Recent advances in understanding the adaptive evolution of metabolic genes and traits. Curr. Opin. Clin. Nutr. Metab. Care 2021, 24, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; Zheng, L.; Almeida, F.A. Epigenetic reprogramming in metabolic disorders: Nutritional factors and beyond. J. Nutr. Biochem. 2018, 54, 1–10. [Google Scholar] [CrossRef]
- Parrillo, L.; Spinelli, R.; Nicolò, A.; Longo, M.; Mirra, P.; Raciti, G.A.; Miele, C.; Beguinot, F. Nutritional Factors, DNA Methylation, and Risk of Type 2 Diabetes and Obesity: Perspectives and Challenges. Int. J. Mol. Sci. 2019, 20, 2983. [Google Scholar] [CrossRef]
- Lillycrop, K.A.; Burdge, G.C. Maternal diet as a modifier of offspring epigenetics. J. Dev. Orig. Health Dis. 2015, 6, 88–95. [Google Scholar] [CrossRef]
- Vanamala, J.K.P.; Sivaramakrishnan, V.; Mummidi, S. Editorial: Integrated multi-omic studies of metabolic syndrome, diabetes and insulin-related disorders: Mechanisms, biomarkers, and therapeutic targets. Front. Endocrinol. 2025, 15, 1537554. [Google Scholar] [CrossRef]
- Liu, J.; Liu, S.; Yu, Z.; Qiu, X.; Jiang, R.; Li, W. Uncovering the gene regulatory network of type 2 diabetes through multi-omic data integration. J. Transl. Med. 2022, 20, 604. [Google Scholar] [CrossRef]
- Rönn, T.; Perfilyev, A.; Oskolkov, N.; Ling, C. Predicting type 2 diabetes via machine learning integration of multiple omics from human pancreatic islets. Sci. Rep. 2024, 14, 14637. [Google Scholar] [CrossRef]
- Carrasco-Zanini, J.; Pietzner, M.; Wheeler, E.; Kerrison, N.D.; Langenberg, C.; Wareham, N.J. Multi-omic prediction of incident type 2 diabetes. Diabetologia 2024, 67, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Birhanu, T.E.; Guracho, Y.D.; Asmare, S.W.; Olana, D.D. A mobile health application use among diabetes mellitus patients: A systematic review and meta-analysis. Front. Endocrinol. 2024, 15, 1481410. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhang, H.; Zhu, R.; Yang, H.; Chen, M.; Wang, X.; Li, Z.; Xiong, Z. Factors affecting the willingness of patients with type 2 diabetes to use digital disease management applications: A cross-sectional study. Front. Public Health 2023, 11, 1259158. [Google Scholar] [CrossRef] [PubMed]
- Salinari, A.; Machì, M.; Armas Diaz, Y.; Cianciosi, D.; Qi, Z.; Yang, B.; Ferreiro Cotorruelo, M.S.; Villar, S.G.; Dzul Lopez, L.A.; Battino, M.; et al. The Application of Digital Technologies and Artificial Intelligence in Healthcare: An Overview on Nutrition Assessment. Diseases 2023, 11, 97. [Google Scholar] [CrossRef]
- Mir, R.R.; Ul Haq, N.; Ishaq, K.; Safie, N.; Dogar, A.B. Impact of machine learning on dietary and exercise behaviors in type 2 diabetes self-management: A systematic literature review. PeerJ Comput. Sci. 2025, 11, e2568. [Google Scholar] [CrossRef]
- Mortazavi, B.J.; Gutierrez-Osuna, R. A Review of Digital Innovations for Diet Monitoring and Precision Nutrition. J. Diabetes Sci. Technol. 2021, 17, 217–223. [Google Scholar] [CrossRef]
- Ordovas, J.M.; Ferguson, L.R.; Tai, E.S.; Mathers, J.C. Personalised nutrition and health. BMJ 2018, 361, bmj.k2173. [Google Scholar] [CrossRef]
- Phillips, C.M. Nutrigenetics and Metabolic Disease: Current Status and Implications for Personalised Nutrition. Nutrients 2013, 5, 32–57. [Google Scholar] [CrossRef]
- Kohlmeier, M.; De Caterina, R.; Ferguson, L.R.; Görman, U.; Allayee, H.; Prasad, C.; Kang, J.X.; Nicoletti, C.F.; Martinez, J.A. Guide and Position of the International Society of Nutrigenetics/Nutrigenomics on Personalized Nutrition: Part 2—Ethics, Challenges and Endeavors of Precision Nutrition. J. Nutr. Nutr. 2016, 9, 28–46. [Google Scholar] [CrossRef]

{kind=link}
| Study/Authors (Year) | Population | Sample Size (n) | Study Design | Genes/SNPs | Dietary Factors | Key Findings |
|---|---|---|---|---|---|---|
| Apio et al., 2023 [33] | Korean | 10,038 | Cohort | GWAS 1 scan | Food scores based on 46 food items | Food scores were positively associated with T2D risk (OR = 1.11, 95% CI: 1.03–1.20, p = 0.009 and OR = 1.10, 95% CI: 1.02–1.19, p = 0.011). Interaction analyses between food scores and SNPs identified 12 candidate genes (e.g., CACNA2D3, RELN, DOCK2) and implicated the adipocytokine signaling pathway as the most strongly associated with T2D, involving 32 key genes including STAT3, IRS1, AKT1–3, and ADIPOR2. Individuals at high genetic risk were more susceptible to the adverse effects of lower diet quality. |
| Karvela et al., 2024 [30] | UK | 148 | RCT 1 | 12 SNPs | Standard care vs. DNA-based diet based on participants’ three highest risk results | DNA-based dietary intervention significantly reduced fasting plasma glucose (−0.019 mmol/L, p = 0.01) and HbA1c (−0.038, p = 0.04) compared to standard care at 26 weeks but not earlier. |
| Merino et al., 2022 [29] | US | 35,759 | Cohort | PRS based on 67 SNPs | AHEI and DASH | Each 1 SD increase in the PRS was associated with a 29% higher T2DM risk (RR = 1.29, 95% CI: 1.25–1.32, p < 0.001), and each 10-unit decrease in the AHEI score conferred a 13% higher risk (RR = 1.13, 95% CI: 1.09–1.17, p < 0.001). Poor diet quality increased T2DM risk by ~30% regardless of genetic risk (P interaction = 0.69), with no significant interaction between diet and genetics (P interaction = 0.30). |
| Westerman, Meigs and Manning, 2023 [31] | UK | 136,880 | Cohort | GWAS scan | MDS | G × D interactions influencing HbA1c were primarily driven by alcohol and fish intake, with a significant interaction between nut intake and LIN9 variants (p = 8.8 × 10−8). The top SNP (rs9729447) showed that the inverse association between nut intake and HbA1c was attenuated in major allele carriers. |
| Westerman et al., 2023 [12] | African American, American Indian, Asian, White, and Hispanic/Latino | 33,178 | Cohort | GWAS scan | Isocaloric substitution of carbohydrates for fats | A higher carbohydrate intake was modestly associated with lower glycemic traits (−0.013% HbA1c per 250 kcal substitution). A significant G × D interaction was identified for rs79762542, enriched in African ancestry, where the inverse carbohydrate–HbA1c association was observed only in major allele homozygotes and replicated in the UK Biobank. |
| Yang et al., 2023 [32] | UK | 142,271 | Cohort | PRS based on 424 SNPs | IDI 1 score based on the sum of 34 food items (16 anti-inflammatory and 18 pro-inflammatory) | Over a median 8.4-year follow-up, low IDI scores were associated with reduced T2DM risk in both normoglycemic (HR = 0.71) and prediabetic (HR = 0.81) individuals. A low-inflammatory diet delayed T2DM onset by ~2.2 years in normoglycemia and ~1.1 years in prediabetes. Joint analysis showed up to a 74% reduced risk among those with both low genetic risk and low IDI scores. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kapellou, A.; Salata, E.; Vrachnos, D.M.; Papailia, S.; Vittas, S. Gene–Diet Interactions in Diabetes Mellitus: Current Insights and the Potential of Personalized Nutrition. Genes 2025, 16, 578. https://doi.org/10.3390/genes16050578
Kapellou A, Salata E, Vrachnos DM, Papailia S, Vittas S. Gene–Diet Interactions in Diabetes Mellitus: Current Insights and the Potential of Personalized Nutrition. Genes. 2025; 16(5):578. https://doi.org/10.3390/genes16050578
Chicago/Turabian StyleKapellou, Angeliki, Effie Salata, Dimitrios Miltiadis Vrachnos, Sevastiani Papailia, and Spiros Vittas. 2025. "Gene–Diet Interactions in Diabetes Mellitus: Current Insights and the Potential of Personalized Nutrition" Genes 16, no. 5: 578. https://doi.org/10.3390/genes16050578
APA StyleKapellou, A., Salata, E., Vrachnos, D. M., Papailia, S., & Vittas, S. (2025). Gene–Diet Interactions in Diabetes Mellitus: Current Insights and the Potential of Personalized Nutrition. Genes, 16(5), 578. https://doi.org/10.3390/genes16050578