
Unraveling the Mitochondrial Blueprint: Genome Characterization and Phylogenetic Insights of the Endemic Fish Onychostoma virgulatum (Teleostei: Cyprinidae)


Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling, DNA Extraction, PCR Amplification, and Sequencing
2.2. Genome Assembly, Annotation, and Selection Pressure Analysis
2.3. Phylogenetic Analysis
3. Results
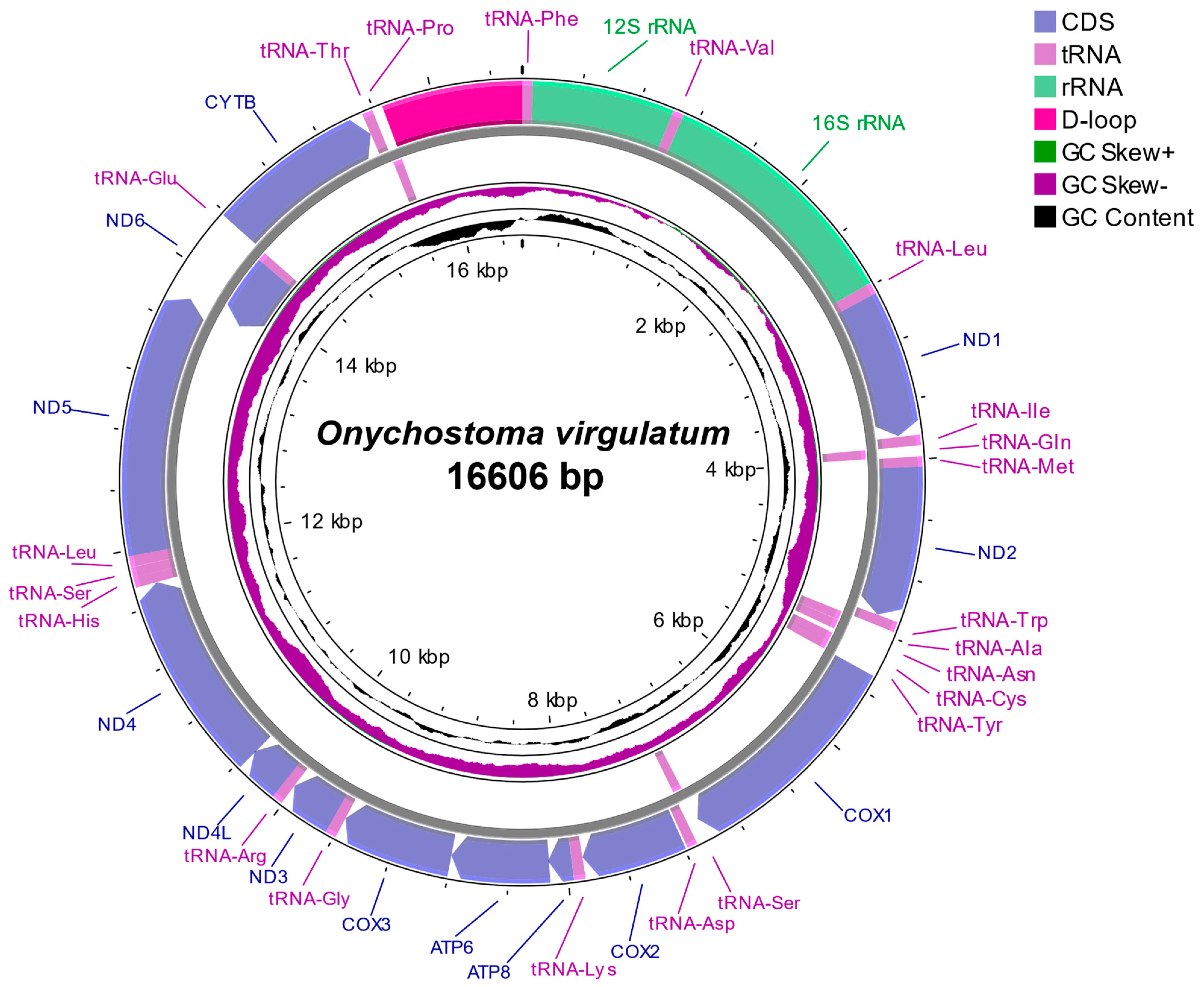
3.1. Mitochondrial Genomic Structure and Composition
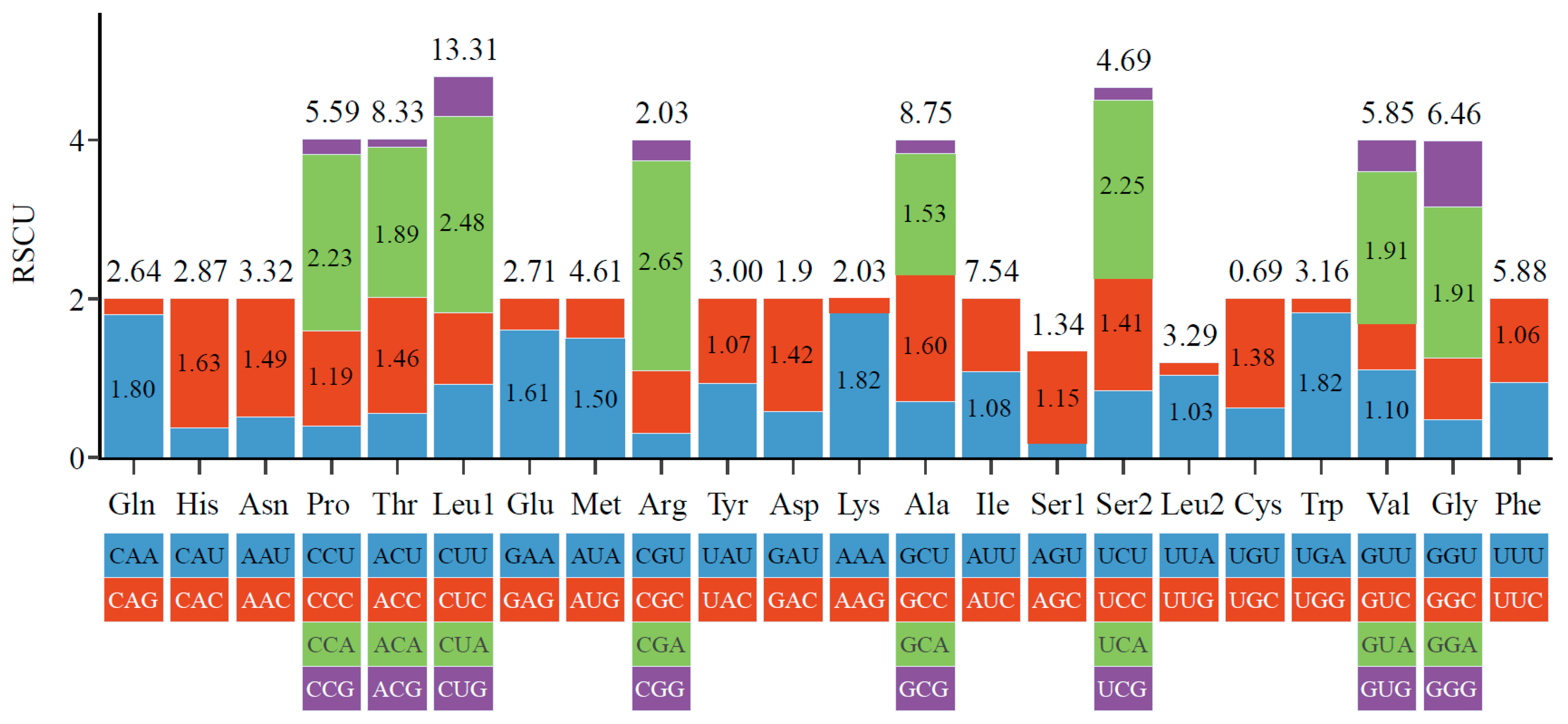
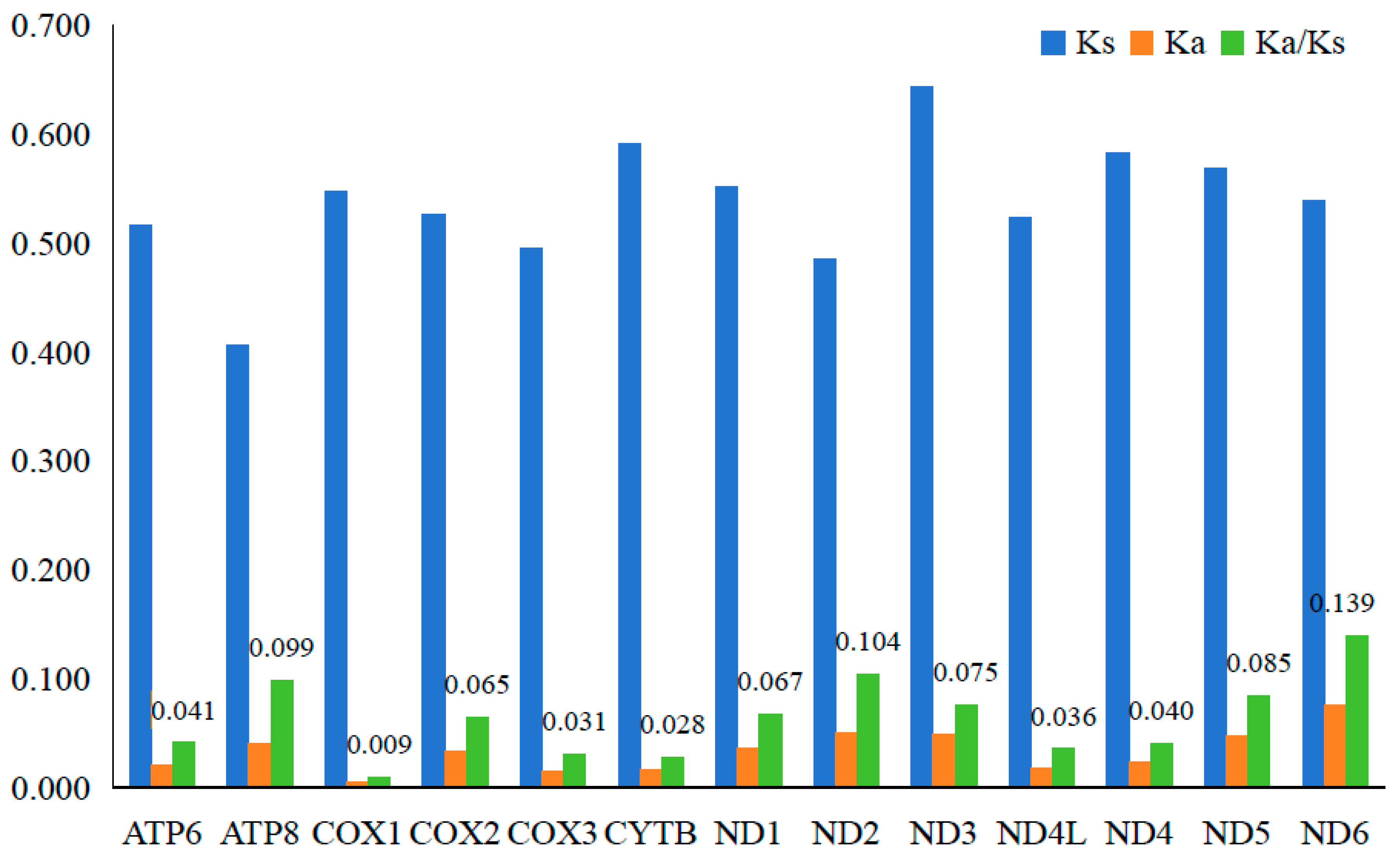
3.2. Characteristics of Codon Usage and Selection Pressure
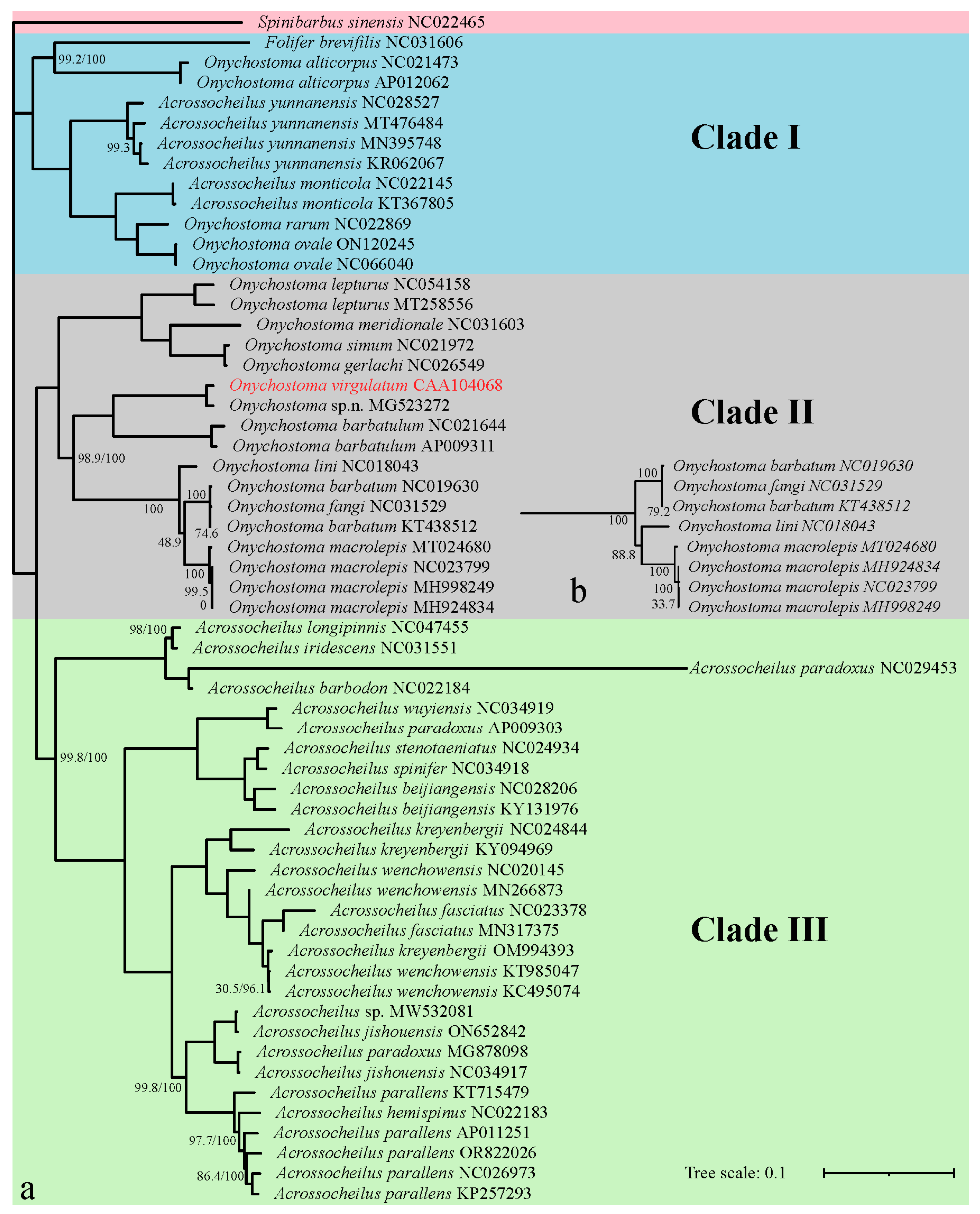
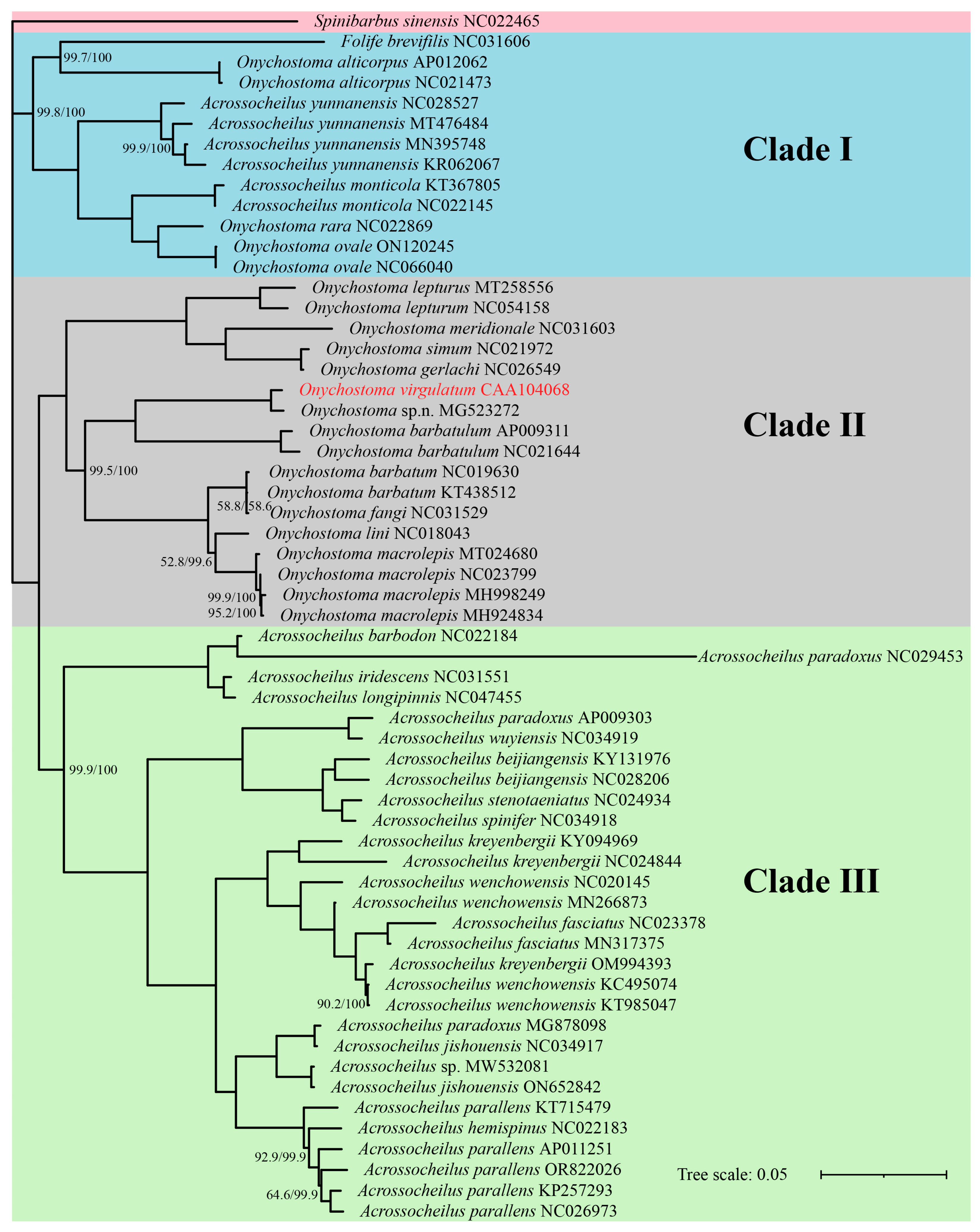
3.3. Phylogenetic Relationship
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bǎnǎrescu, P. A review of the species of the subgenus Onychostoma s. str. with description of a new species (Pisces, Cyprinidae). Rev. Roum. Biol. Ser. Zool. 1971, 16, 241–248. [Google Scholar]
- Bǎnǎrescu, P. Revision of the Onychostoma subgenus Scaphesthes (Pisces, Cyprinidae). Rev. Roum. Biol. Ser. Zool. 1971, 16, 357–364. [Google Scholar]
- Tan, M.; Armbruster, J.W. Phylogenetic classification of extant genera of fishes of the order Cypriniformes (Teleostei: Ostariophysi). Zootaxa 2018, 4476, 6–39. [Google Scholar] [CrossRef] [PubMed]
- Shan, X.H.; Lin, R.R.; Yue, P.Q.; Chu, X.L. Barbinae. In Fauna Sinica Osteichthyes: Cypriniformes (III); Yue, P.Q., Ed.; Science Press: Beijing, China, 2000; pp. 126–130. [Google Scholar]
- Rainboth, W.J. Fishes of the Cambodian Mekong; Food and Agriculture Organization of the United Nations: Rome, Italy, 1996; p. 265. [Google Scholar]
- Kottelat, M. Fishes of the Nam Theun and Xe Bangfai basin, Loas, with diagnosis of twenty-two new species (Teleostei: Cypribidae, Baltoridae, Cobitidae, Coiidae and Odontobutidae). Ichthyol. Explor. Freshw. 1998, 9, 1–128. [Google Scholar]
- Kottelat, M. Fishes of Laos; Wildlife Heritage Trust Publications: Colombo, Sri Lanka, 2001; p. 198. [Google Scholar]
- Kottelat, M. Freshwater Fishes of Northern Vietnam; World Bank: Washington, DC, USA, 2001; p. 123. [Google Scholar]
- Nguyen, V.H.; Ngo, S.V. Freshwater Fishes of Vietnam; Volume I: Family Cyprinidae; Agriculture Publish House: Hanoi, Vietnam, 2001. [Google Scholar]
- Xin, Q.; Zhang, E.; Cao, W.X. Onychostoma virgulatum, a new species of cyprinid fish (Pisces: Teleostei) from southern Anhui Province, South China. Ichthyol. Explor. Freshw. 2009, 20, 255–266. [Google Scholar]
- Jang-Liaw, N.H.; Chen, I.S. Onychostoma minnanensis, a new cyprinid species (Teleostei: Cyprinidae) from Fujian, southern mainland China with comments on the mitogenetic differentiation among related species. Ichthyol. Res. 2013, 60, 62–74. [Google Scholar] [CrossRef]
- Hoang, H.D.; Pham, H.M.; Tran, N.T. Two new species of shovel-jaw carp Onychostoma (Teleostei: Cyprinidae) from southern Vietnam. Zootaxa 2015, 3962, 123–138. [Google Scholar] [CrossRef]
- Song, X.L.; Cao, L.; Zhang, E. Onychostoma brevibarba, a new cyprinine fish (Pisces: Teleostei) from the middle Chang Jiang basin in Hunan province, south China. Zootaxa 2018, 4410, 147–163. [Google Scholar] [CrossRef]
- Wang, X.Z.; Li, J.B.; He, S.P. Molecular evidence for the monophyly of East Asian groups of Cyprinidae (Teleostei: Cypriniformes) derived from the nuclear recombination activating gene 2 sequences. Mol. Phylogenet. Evol. 2007, 42, 157–170. [Google Scholar] [CrossRef]
- Li, J.B.; Wang, X.Z.; Kong, X.H.; Zhao, K.; He, S.P.; Richard, L.M. Variation patterns of the mitochondrial 16S rRNA gene with secondary structure constraints and their application to phylogeny of cyprinine fishes (Teleostei: Cypriniformes). Mol. Phylogenet. Evol. 2008, 47, 472–487. [Google Scholar] [CrossRef]
- Xin, Q. Taxonomic Revision of Species and Phylogenetic Analysis of Interspecific Relationships Within the Cyprinid Genus Onychostoma Sensu Lato Günther, 1896. Master’s Thesis, Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan, China, 2008. [Google Scholar]
- Yuan, L.Y. Monophyly, Affinity and Taxonomic Revision of the Cyprinid Genus Acrossocheilus Oshima, 1919. Ph.D. Thesis, Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan, China, 2009. [Google Scholar]
- Yang, L.; Sado, T.; Hirt, M.V.; Pasco-Viel, E.; Arunachalam, M.; Li, J.B.; Wang, X.Z.; Freyhof, J.; Saitoh, K.; Simons, A.M.; et al. Phylogeny and polyploidy: Resolving the classification of cyprinine fishes (Teleostei: Cypriniformes). Mol. Phylogenet. Evol. 2015, 85, 97–116. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.Y.; Liu, X.X.; Zhang, E. Mitochondrial phylogeny of Chinese barred species of the cyprinid genus Acrossocheilus Oshima, 1919 (Teleostei: Cypriniformes) and its taxonomic implications. Zootaxa 2015, 4059, 151–168. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Cheng, Q.; Geng, H.; Lin, A.H.; Wang, H.Y. Molecular phylogeny of Onychostoma (Cyprinidae) based on mitochondrial genomes. Acta Hydrobiol. Sin. 2018, 42, 512–516. [Google Scholar] [CrossRef]
- Wang, Z.T.; Zhang, E. An updated species checklist of freshwater fishes from the Ganjiang River. Biodivers. Sci. 2021, 29, 1256–1264. Available online: https://www.biodiversity-science.net/CN/10.17520/biods.2021119 (accessed on 27 April 2025). [CrossRef]
- Cheng, W.J.; Fu, H.Y. The Fishes of Jiangxi; Science Press: Beijing, China, 2024; p. 78. [Google Scholar]
- Guo, Z.Z.; Liu, R.L. Study on fishes from Jiangxi province. J. Nanchang Univ. (Nat. Sci.) 1995, 19, 222–232. [Google Scholar]
- Zou, D.L. Fish resources in Xunwushui, Jiangxi province. Chin. J. Zool. 1988, 23, 15–17. [Google Scholar]
- Cheng, C. Complete Mitochondrial Genome Sequencing and Analysis of Onychostoma gerlachi and Phylogenetic Research of Genus Onychostoma. Master’s Thesis, South-Central University for Nationalities, Wuhan, China, 2013. [Google Scholar]
- Xiao, W.H.; Zhang, Y.P.; Liu, H.Z. Molecular systematics of Xenocyprinae (teleostei: Cyprinidae): Taxonomy, biogeography, and coevolution of a special group restricted in East Asia. Mol. Phylogenet. Evol. 2001, 18, 163–173. [Google Scholar] [CrossRef]
- Liu, H.Z.; Tzeng, C.S.; Teng, H.Y. Sequence variations in the mitochondrial DNA control region and their implications for the phylogeny of the Cypriniformes. Can. J. Zool. 2002, 80, 569–581. [Google Scholar] [CrossRef]
- Galtier, N.; Gouy, M.; Gautier, C. SEAVIEW and PHY-LO_WIN: Two graphic tools for sequence alignment and molecular phylogeny. Bioinformatics 1996, 12, 543–548. [Google Scholar] [CrossRef]
- Perna, N.T.; Kocher, T.D. Patterns of nucleotide composition at fourfold degenerate sites of animal mitochondrial genomes. J. Mol. Evol. 1995, 41, 353–358. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Suleski, M.; Sanderford, M.; Sharma, M.; Tamura, K. MEGA 12: Molecular Evolutionary Genetic Analysis Version 12 for Adaptive and Green Computing. Mol. Biol. Evol. 2024, 41, msae263. [Google Scholar] [CrossRef]
- Zhang, D.; Gao, F.L.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Misawa, K.; Kuma, K.-I.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed]
- Vincent, L.; Jean-Emmanuel, L.; Olivier, G. SMS: Smart model selection in PhyML. Mol. Biol. Evol. 2017, 34, 2422–2424. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Xiang, C.Y.; Gao, F.L.; Jakovlić, I.; Lei, H.P.; Hu, Y.; Zhang, H.; Zou, H.; Wang, G.T.; Zhang, D. Using PhyloSuite for molecular phylogeny and tree-based analyses. iMeta 2023, 2, e87. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Wang, I.C.; Lin, H.D.; Liang, C.M.; Huang, C.C.; Wang, R.D.; Yang, J.Q.; Wang, W.K. Complete mitochondrial genome of the freshwater fish Onychostoma lepturum (Teleostei, Cyprinidae): Genome characterization and phylogenetic analysis. ZooKeys 2020, 1005, 57–72. [Google Scholar] [CrossRef]
- Chen, S.W.; Tang, Q.Y. The complete mitochondrial genome of a vulnerable cyprinid fish Onychostoma macrolepis (teleostei: Cypriniformes) from Qinling-Bashan mountain area in China. Mitochondrial DNA Part B 2020, 5, 1640–1641. [Google Scholar] [CrossRef]
- Zhang, R.Y.; Zhu, T.T.; Luo, Q. The complete mitochondrial genome of the freshwater fish Onychostoma ovale (Cypriniformes, Cyprinidae): Genome characterization and phylogenetic analysis. Genes 2023, 14, 1227. [Google Scholar] [CrossRef]
- Lan, X.Y.; Wang, J.X.; Zhang, M.Y.; Zhou, Q.; Xiang, H.M.; Jiang, W.S. Molecular identification of Acrossocheilus jishouensis (Teleostei: Cyprinidae) and its complete mitochondrial genome. Biochem. Genet. 2024, 62, 1396–1412. [Google Scholar] [CrossRef] [PubMed]
- Saccone, C.; Gissi, C.; Lanave, C.; Larizza, A.; Pesole, G.; Reyes, A. Evolution of the mitochondrial genetic system: An overview. Gene 2000, 261, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Pesini, E.; Lott, M.T.; Procaccio, V.; Poole, J.C.; Brandon, M.C.; Mishmar, D.; Yi, C.; Kreuziger, J.; Baldi, P.; Wallace, D.C. An enhanced MITOMAP with a global mtDNA mutational phylogeny. Nucleic Acids Res. 2007, 35, D823–D828. [Google Scholar] [CrossRef]
- Nabholz, B.; Glémin, S.; Galtier, N. Extreme variation of mtDNA neutral substitution rate across mammalian species—The longevity hypothesis. Mol. Biol. Evol. 2008, 25, 795. [Google Scholar] [CrossRef]
- Shen, S.C. Fishes of Taiwan; Department of Zoology, National Taiwan University: Taipei, China, 1993; p. 960. [Google Scholar]
- Yuan, L.Y.; Wu, Z.Q.; Zhang, E. Acrossocheilus spinifer, a new species of barred cyprinid fish from south China (Pisces: Teleostei). J. Fish. Biol. 2006, 68, 163–173. [Google Scholar] [CrossRef]
- Dayrat, B. Towards integrative taxonomy. Biol. J. Linn. Soc. 2005, 85, 407–417. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NO. | Valid Species | Author | Accession NO. | Size(bp) | AT(%) | AT-Skew | GC-Skew |
|---|---|---|---|---|---|---|---|
| 1 | Acrossocheilus beijiangensis | Wu and Lin, 1977 | NC028206 | 16,600 | 56.2 | 0.11 | −0.26 |
| KY131976 | 16,596 | 55.9 | 0.11 | −0.27 | |||
| 2 | Acrossocheilus fasciatus | Steindachner, 1892 | NC023378 | 16,589 | 55.7 | 0.11 | −0.26 |
| MN317375 | 16,589 | 55.7 | 0.11 | −0.26 | |||
| 3 | Acrossocheilus hemispinus | Nichols, 1925 | NC022183 | 16,590 | 55.9 | 0.12 | −0.27 |
| 4 | Acrossocheilus iridescens (Acrossocheilus barbodon) | Nichols and Pope, 1927 | NC031551 | 16,596 | 55.9 | 0.13 | −0.28 |
| NC022184 | 16,596 | 55.9 | 0.13 | −0.28 | |||
| 5 | Acrossocheilus jishouensis | Zhao, Chen, and Li, 1997 | NC034917 | 16,587 | 56.3 | 0.11 | −0.26 |
| ON652842 | 16,594 | 56.3 | 0.11 | −0.26 | |||
| 6 | Acrossocheilus kreyenbergii | Regan, 1908 | NC024844 | 16,849 | 56.6 | 0.10 | −0.25 |
| KY094969 | 16,596 | 56.0 | 0.10 | −0.25 | |||
| OM994393 | 16,592 | 55.8 | 0.11 | −0.26 | |||
| 7 | Acrossocheilus longipinnis (Acrossocheilus stenotaeniatus) | Wu, 1939 | NC047455 | 16,593 | 55.9 | 0.13 | −0.27 |
| NC024934 | 16,594 | 55.9 | 0.12 | −0.27 | |||
| 8 | Acrossocheilus monticola | Günther, 1888 | NC022145 | 16,599 | 55.9 | 0.12 | −0.28 |
| KT367805 | 16,605 | 55.9 | 0.12 | −0.28 | |||
| 9 | Acrossocheilus paradoxus | Günther, 1868 | NC029453 | 16,597 | 54.8 | 0.07 | −0.24 |
| AP009303 | 16,595 | 55.6 | 0.11 | −0.26 | |||
| MG878098 | 16,586 | 56.2 | 0.11 | −0.26 | |||
| 10 | Acrossocheilus parallens | Nichols, 1931 | NC026973 | 16,592 | 55.7 | 0.11 | −0.26 |
| AP011251 | 16,590 | 55.6 | 0.11 | −0.26 | |||
| KP257293 | 16,588 | 55.7 | 0.11 | −0.26 | |||
| KT715479 | 16,591 | 55.5 | 0.11 | −0.26 | |||
| OR822026 | 16,589 | 55.5 | 0.11 | −0.26 | |||
| 11 | Acrossocheilus spinifer | Yuan, Wu, and Zhang, 2006 | NC034918 | 16,591 | 55.9 | 0.12 | −0.27 |
| 12 | Acrossocheilus wenchowensis | Wang, 1935 | NC020145 | 16,591 | 55.8 | 0.11 | −0.26 |
| KC495074 | 16,591 | 55.8 | 0.11 | −0.26 | |||
| KT985047 | 16,591 | 55.8 | 0.11 | −0.26 | |||
| MN266873 | 16,593 | 56.0 | 0.11 | −0.26 | |||
| 13 | Acrossocheilus wuyiensis | Wu and Chen, 1981 | NC034919 | 16,594 | 55.8 | 0.11 | −0.27 |
| 14 | Acrossocheilus yunnanensis | Regan, 1904 | NC028527 | 16,588 | 56.1 | 0.12 | −0.27 |
| MN395748 | 16,590 | 56.1 | 0.12 | −0.27 | |||
| MT476484 | 16,587 | 56.0 | 0.12 | −0.27 | |||
| KR062067 * | 16,596 | 56.2 | 0.12 | −0.26 | |||
| 15 | Acrossocheilus sp. | MW532081 * | 16,594 | 56.3 | 0.11 | −0.26 | |
| 16 | Onychostoma alticorpus | Oshima, 1920 | AP012062 | 16,604 | 54.5 | 0.13 | −0.27 |
| NC021473 | 16,607 | 54.5 | 0.13 | −0.27 | |||
| 17 | Onychostoma barbatulum | Pellegrin, 1908 | AP009311 | 16,612 | 56.7 | 0.11 | −0.27 |
| NC021644 | 16,597 | 56.6 | 0.11 | −0.26 | |||
| 18 | Onychostoma barbatum | Lin,1931 | KT438512 | 16,589 | 56.0 | 0.13 | −0.28 |
| NC019630 | 16,592 | 56.0 | 0.13 | −0.28 | |||
| 19 | Onychostoma fangi | Kottelat, 2000 | NC031529 | 16,597 | 56.0 | 0.13 | −0.28 |
| 20 | Onychostoma gerlachi | Peters, 1881 | NC026549 | 16,601 | 55.6 | 0.13 | −0.27 |
| 21 | Onychostoma lepturus | Boulenger, 1900 | MT258556 | 16,598 | 55.3 | 0.13 | −0.28 |
| NC054158 | 16,601 | 55.2 | 0.13 | −0.28 | |||
| 22 | Onychostoma lini | Wu, 1939 | NC018043 | 16,595 | 56.2 | 0.12 | −0.27 |
| 23 | Onychostoma macrolepis | Bleeker, 1871 | MH924834 | 16,594 | 55.7 | 0.12 | −0.27 |
| MH998249 | 16,621 | 55.8 | 0.12 | −0.27 | |||
| MT024680 | 16,597 | 55.8 | 0.12 | −0.27 | |||
| NC023799 | 16,595 | 55.8 | 0.12 | −0.27 | |||
| 24 | Onychostoma meridionale | Kottelat, 1998 | NC031603 | 16,595 | 55.5 | 0.12 | −0.27 |
| 25 | Onychostoma ovale | Pellegrin and Chevey, 1936 | NC066040 | 16,602 | 55.6 | 0.13 | −0.28 |
| ON120245 | 16,600 | 55.6 | 0.13 | −0.28 | |||
| 26 | Onychostoma rarum | Lin, 1933 | NC022869 | 16,590 | 55.7 | 0.13 | −0.28 |
| 27 | Onychostoma simum | Sauvage and Dabry de Thiersant, 1874 | NC021972 | 16,601 | 55.6 | 0.13 | −0.27 |
| 28 | Onychostoma sp. n. | MG523272 * | 16,602 | 56.5 | 0.11 | −0.26 | |
| 29 | Onychostoma virgulatum | Xin, Zhang, and Cao, 2009 | CAA104068 a | 16,606 | 56.5 | 0.11 | −0.26 |
| 30 | Folifer brevifilis | Peters, 1881 | NC031606 | 16,707 | 55.7 | 0.12 | −0.26 |
| 31 | Spinibarbus sinensis | Bleeker, 1871 | NC022465 | 16,591 | 57.4 | 0.11 | −0.25 |
| Number | Genes | Location (bp) | Size (bp) | Intergenic Spacer (bp) | Coding Strand | Condon | |
|---|---|---|---|---|---|---|---|
| Start | End | ||||||
| 1 | tRNA-Phe | 1–69 | 69 | 0 | H | ||
| 2 | 12S rRNA | 70–1025 | 956 | 0 | H | ||
| 3 | tRNA-Val | 1026–1097 | 72 | 0 | H | ||
| 4 | 16S rRNA | 1098–2781 | 1684 | 0 | H | ||
| 5 | tRNA-Leu | 2782–2857 | 76 | 0 | H | ||
| 6 | ND1 | 2858–3832 | 975 | 6 | H | ATG | TAA |
| 7 | tRNA-Ile | 3839–3909 | 71 | −2 | H | ||
| 8 | tRNA-Gln | 3978–3908 | 71 | 3 | L | ||
| 9 | tRNA-Met | 3982–4050 | 69 | 0 | H | ||
| 10 | ND2 | 4051–5095 | 1045 | 0 | H | ATG | T-- |
| 11 | tRNA-Trp | 5096–5165 | 70 | 2 | H | ||
| 12 | tRNA-Ala | 5237–5168 | 70 | 1 | L | ||
| 13 | tRNA-Asn | 5311–5239 | 73 | 33 | L | ||
| 14 | tRNA-Cys | 5411–5345 | 67 | −1 | L | ||
| 15 | tRNA-Tyr | 5481–5411 | 71 | 1 | L | ||
| 16 | COX1 | 5483–7033 | 1551 | 0 | H | GTG | TAA |
| 17 | tRNA-Ser | 7104–7034 | 71 | 5 | L | ||
| 18 | tRNA-Asp | 7110–7179 | 70 | 13 | H | ||
| 19 | COX2 | 7193–7883 | 691 | 0 | H | ATG | T-- |
| 20 | tRNA-Lys | 7884–7959 | 76 | 1 | H | ||
| 21 | ATP8 | 7961–8125 | 165 | −7 | H | ATG | TAG |
| 22 | ATP6 | 8119–8802 | 684 | −1 | H | ATG | TAA |
| 23 | COX3 | 8802–9586 | 785 | 0 | H | ATG | TA- |
| 24 | tRNA-Gly | 9587–9659 | 73 | 0 | H | ||
| 25 | ND3 | 9660–10,008 | 349 | 0 | H | ATG | T-- |
| 26 | tRNA-Arg | 10,009–10,079 | 71 | 0 | H | ||
| 27 | ND4L | 10,080–10,376 | 297 | −7 | H | ATG | TAA |
| 28 | ND4 | 10,370–11,750 | 1381 | 0 | H | ATG | T-- |
| 29 | tRNA-His | 11,751–11,819 | 69 | 0 | H | ||
| 30 | tRNA-Ser | 11,820–11,888 | 69 | 1 | H | ||
| 31 | tRNA-Leu | 11,890–11,962 | 73 | 0 | H | ||
| 32 | ND5 | 11,963–13,786 | 1824 | −4 | H | ATG | TAA |
| 33 | ND6 | 14,304–13,783 | 522 | 0 | L | ATG | TAA |
| 34 | tRNA-Glu | 14,373–14,305 | 69 | 5 | L | ||
| 35 | CYTB | 14,379–15,519 | 1141 | 0 | H | ATG | T-- |
| 36 | tRNA-Thr | 15,520–15,591 | 72 | −1 | H | ||
| 37 | tRNA-Pro | 15,661–15,591 | 71 | 0 | L | ||
| 38 | D-loop | 15,662–16,606 | 945 | 0 | H | ||
| Regions | Size (bp) | T (U) | C | A | G | AT (%) | AT-Skew | GC-Skew |
|---|---|---|---|---|---|---|---|---|
| Full genome | 16,606 | 25.1 | 27.4 | 31.4 | 16.1 | 56.5 | 0.112 | −0.262 |
| D-loop | 945 | 32.6 | 21.1 | 33.5 | 12.8 | 66.1 | 0.014 | −0.245 |
| PCGs | 11,403 | 27.0 | 28.1 | 29.4 | 15.5 | 56.4 | 0.042 | −0.288 |
| tRNAs | 1563 | 26.8 | 21.4 | 28.0 | 23.8 | 54.8 | 0.022 | 0.054 |
| rRNAs | 2640 | 20.1 | 24.5 | 34.7 | 20.7 | 54.8 | 0.266 | −0.085 |
| PCGs-1st | 3801 | 20.9 | 26.4 | 27.1 | 25.6 | 48.0 | 0.130 | −0.015 |
| PCGs-2nd | 3801 | 40.4 | 27.3 | 18.6 | 13.7 | 59.0 | −0.369 | −0.333 |
| PCGs-3rd | 3801 | 19.7 | 30.6 | 42.4 | 7.3 | 62.1 | 0.365 | −0.613 |
| ATP6 | 684 | 27.2 | 28.4 | 30.6 | 13.9 | 57.8 | 0.058 | −0.343 |
| ATP8 | 165 | 23.6 | 28.5 | 36.4 | 11.5 | 60.0 | 0.212 | −0.424 |
| COX1 | 1551 | 28.9 | 26.4 | 27.2 | 17.5 | 56.1 | −0.030 | −0.201 |
| COX2 | 691 | 26.5 | 26.6 | 30.7 | 16.2 | 57.2 | 0.073 | −0.243 |
| COX3 | 785 | 26.5 | 28.9 | 27.5 | 17.1 | 54.0 | 0.019 | −0.258 |
| CYTB | 1141 | 28.1 | 28.4 | 28.3 | 15.2 | 56.4 | 0.003 | −0.304 |
| ND1 | 975 | 26.1 | 29.6 | 29.1 | 15.2 | 55.2 | 0.056 | −0.323 |
| ND2 | 1045 | 23.2 | 30.4 | 34.1 | 12.3 | 57.3 | 0.191 | −0.423 |
| ND3 | 349 | 28.4 | 29.2 | 28.4 | 14.0 | 56.8 | 0.000 | −0.351 |
| ND4 | 1381 | 25.6 | 29.2 | 31.2 | 14.0 | 56.8 | 0.098 | −0.352 |
| ND4L | 297 | 29.0 | 28.3 | 26.9 | 15.8 | 55.9 | −0.036 | −0.282 |
| ND5 | 1824 | 24.7 | 30.3 | 32.4 | 12.6 | 57.1 | 0.135 | −0.413 |
| ND6 | 522 | 41.0 | 13.2 | 13.0 | 32.8 | 54.0 | −0.518 | 0.425 |
| 16S rRNA | 1684 | 20.6 | 23.1 | 36.3 | 20.0 | 56.9 | 0.276 | −0.072 |
| 12S rRNA | 956 | 19.2 | 27.0 | 31.9 | 21.9 | 51.1 | 0.247 | −0.105 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, Y.; Duan, G.; Zhou, H.; Wang, H.; Liu, A. Unraveling the Mitochondrial Blueprint: Genome Characterization and Phylogenetic Insights of the Endemic Fish Onychostoma virgulatum (Teleostei: Cyprinidae). Genes 2025, 16, 541. https://doi.org/10.3390/genes16050541
Hu Y, Duan G, Zhou H, Wang H, Liu A. Unraveling the Mitochondrial Blueprint: Genome Characterization and Phylogenetic Insights of the Endemic Fish Onychostoma virgulatum (Teleostei: Cyprinidae). Genes. 2025; 16(5):541. https://doi.org/10.3390/genes16050541
Chicago/Turabian StyleHu, Yuting, Guoqing Duan, Huaxing Zhou, Huan Wang, and Amei Liu. 2025. "Unraveling the Mitochondrial Blueprint: Genome Characterization and Phylogenetic Insights of the Endemic Fish Onychostoma virgulatum (Teleostei: Cyprinidae)" Genes 16, no. 5: 541. https://doi.org/10.3390/genes16050541
APA StyleHu, Y., Duan, G., Zhou, H., Wang, H., & Liu, A. (2025). Unraveling the Mitochondrial Blueprint: Genome Characterization and Phylogenetic Insights of the Endemic Fish Onychostoma virgulatum (Teleostei: Cyprinidae). Genes, 16(5), 541. https://doi.org/10.3390/genes16050541