
Systematic Analysis of UFMylation Family Genes in Tissues of Mice with Metabolic Dysfunction-Associated Steatotic Liver Disease


Highlights
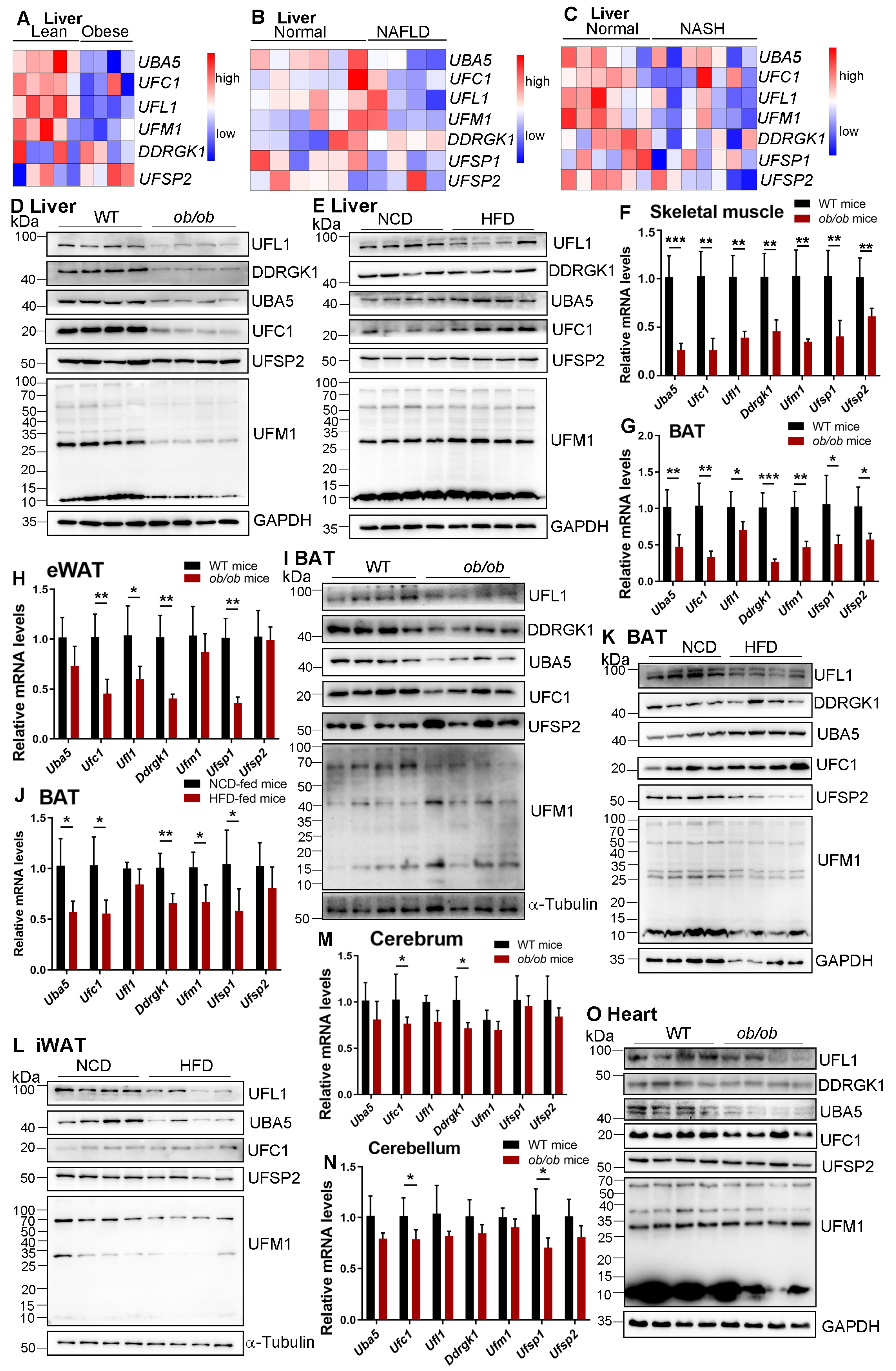
- Our findings established the expressed profiles of UFMylation in multiple tissues of mice mimicking MASLD.
- We found that the expression of UFMylation components, especially the UFM1-specific ligase UFL1, was decreased in multiple metabolic tissues of the mouse models.
- Our findings suggest novel physiological functions for UFMylation in MASLD development and progression.
- Our study clarifies that PTM regulation of UFMylation family proteins themselves is also important in understanding the biological functions and regulation of UFMylation.
- Our findings imply promising therapeutic strategies in the treatment of MASLD and other metabolic diseases by targeting UFMylation.
Abstract
1. Introduction
2. Materials and Methods
2.1. Antibodies
2.2. Animals
2.3. Western Blot Analysis
2.4. Real-Time PCR Assays
2.5. Statistical Analysis
3. Results
4. Conclusions
Limitations of This Study
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rinella, M.E.; Sookoian, S. From NAFLD to MASLD: Updated naming and diagnosis criteria for fatty liver disease. J. Lipid Res. 2024, 65, 100485. [Google Scholar] [CrossRef] [PubMed]
- Lonardo, A.; Nascimbeni, F.; Maurantonio, M.; Marrazzo, A.; Rinaldi, L.; Adinolfi, L.E. Nonalcoholic fatty liver disease: Evolving paradigms. World J. Gastroenterol. 2017, 23, 6571–6592. [Google Scholar] [CrossRef] [PubMed]
- Sacco, F.; Seelig, A.; Humphrey, S.J.; Krahmer, N.; Volta, F.; Reggio, A.; Marchetti, P.; Gerdes, J.; Mann, M. Phosphoproteomics Reveals the GSK3-PDX1 Axis as a Key Pathogenic Signaling Node in Diabetic Islets. Cell Metab. 2019, 29, 1422–1432.e3. [Google Scholar] [CrossRef]
- Banner, B.F.; Savas, L.; Zivny, J.; Tortorelli, K.; Bonkovsky, H.L. Ubiquitin as a marker of cell injury in nonalcoholic steatohepatitis. Am. J. Clin. Pathol. 2000, 114, 860–866. [Google Scholar] [CrossRef]
- Wang, Y.; Wen, H.; Fu, J.; Cai, L.; Li, P.L.; Zhao, C.L.; Dong, Z.F.; Ma, J.P.; Wang, X.; Tian, H.; et al. Hepatocyte TNF Receptor-Associated Factor 6 Aggravates Hepatic Inflammation and Fibrosis by Promoting Lysine 6-Linked Polyubiquitination of Apoptosis Signal-Regulating Kinase 1. Hepatology 2020, 71, 93–111. [Google Scholar] [CrossRef]
- Zubiete-Franco, I.; Fernandez-Tussy, P.; Barbier-Torres, L.; Simon, J.; Fernandez-Ramos, D.; Lopitz-Otsoa, F.; Gutierrez-de Juan, V.; de Davalillo, S.L.; Duce, A.M.; Iruzubieta, P.; et al. Deregulated neddylation in liver fibrosis. Hepatology 2017, 65, 694–709. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, M.; Chiba, T.; Tatsumi, K.; Iemura, S.; Tanida, I.; Okazaki, N.; Ueno, T.; Kominami, E.; Natsume, T.; Tanaka, K. A novel protein-conjugating system for Ufm1, a ubiquitin-fold modifier. EMBO J. 2004, 23, 1977–1986. [Google Scholar] [CrossRef]
- Tatsumi, K.; Sou, Y.S.; Tada, N.; Nakamura, E.; Iemura, S.; Natsume, T.; Kang, S.H.; Chung, C.H.; Kasahara, M.; Kominami, E.; et al. A novel type of E3 ligase for the Ufm1 conjugation system. J. Biol. Chem. 2010, 285, 5417–5427. [Google Scholar] [CrossRef]
- Millrine, D.; Cummings, T.; Matthews, S.P.; Peter, J.J.; Magnussen, H.M.; Lange, S.M.; Macartney, T.; Lamoliatte, F.; Knebel, A.; Kulathu, Y. Human UFSP1 is an active protease that regulates UFM1 maturation and UFMylation. Cell Rep. 2022, 40, 111168. [Google Scholar] [CrossRef] [PubMed]
- Yoo, H.M.; Kang, S.H.; Kim, J.Y.; Lee, J.E.; Seong, M.W.; Lee, S.W.; Ka, S.H.; Sou, Y.S.; Komatsu, M.; Tanaka, K.; et al. Modification of ASC1 by UFM1 is crucial for ERalpha transactivation and breast cancer development. Mol. Cell 2014, 56, 261–274. [Google Scholar] [CrossRef]
- Wang, Z.; Gong, Y.; Peng, B.; Shi, R.; Fan, D.; Zhao, H.; Zhu, M.; Zhang, H.; Lou, Z.; Zhou, J.; et al. MRE11 UFMylation promotes ATM activation. Nucleic Acids Res. 2019, 47, 4124–4135. [Google Scholar] [CrossRef] [PubMed]
- Qin, B.; Yu, J.; Nowsheen, S.; Wang, M.; Tu, X.; Liu, T.; Li, H.; Wang, L.; Lou, Z. UFL1 promotes histone H4 ufmylation and ATM activation. Nat. Commun. 2019, 10, 1242. [Google Scholar] [CrossRef]
- Liu, J.; Guan, D.; Dong, M.; Yang, J.; Wei, H.; Liang, Q.; Song, L.; Xu, L.; Bai, J.; Liu, C.; et al. UFMylation maintains tumour suppressor p53 stability by antagonizing its ubiquitination. Nat. Cell Biol. 2020, 22, 1056–1063. [Google Scholar] [CrossRef] [PubMed]
- Snider, D.L.; Park, M.; Murphy, K.A.; Beachboard, D.C.; Horner, S.M. Signaling from the RNA sensor RIG-I is regulated by ufmylation. Proc. Natl. Acad. Sci. USA 2022, 119, e2119531119. [Google Scholar] [CrossRef] [PubMed]
- Ishimura, R.; El-Gowily, A.H.; Noshiro, D.; Komatsu-Hirota, S.; Ono, Y.; Shindo, M.; Hatta, T.; Abe, M.; Uemura, T.; Lee-Okada, H.C.; et al. The UFM1 system regulates ER-phagy through the ufmylation of CYB5R3. Nat. Commun. 2022, 13, 7857. [Google Scholar] [CrossRef]
- Zhou, J.; Ma, X.; He, X.; Chen, B.; Yuan, J.; Jin, Z.; Li, L.; Wang, Z.; Xiao, Q.; Cai, Y.; et al. Dysregulation of PD-L1 by UFMylation imparts tumor immune evasion and identified as a potential therapeutic target. Proc. Natl. Acad. Sci. USA 2023, 120, e2215732120. [Google Scholar] [CrossRef] [PubMed]
- Egunsola, A.T.; Bae, Y.; Jiang, M.M.; Liu, D.S.; Chen-Evenson, Y.; Bertin, T.; Chen, S.; Lu, J.T.; Nevarez, L.; Magal, N.; et al. Loss of DDRGK1 modulates SOX9 ubiquitination in spondyloepimetaphyseal dysplasia. J. Clin. Investig. 2017, 127, 1475–1484. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Ye, X.; Zhang, C.; Wang, J.; Guan, Z.; Yan, J.; Xu, L.; Wang, K.; Guan, D.; Liang, Q.; et al. Ufl1 deficiency causes kidney atrophy associated with disruption of endoplasmic reticulum homeostasis. J. Genet. Genom. 2021, 48, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yue, G.; Ma, W.; Zhang, A.; Zou, J.; Cai, Y.; Tang, X.; Wang, J.; Liu, J.; Li, H.; et al. Ufm1-Specific Ligase Ufl1 Regulates Endoplasmic Reticulum Homeostasis and Protects Against Heart Failure. Circ. Heart Fail. 2018, 11, e004917. [Google Scholar] [CrossRef] [PubMed]
- Muona, M.; Ishimura, R.; Laari, A.; Ichimura, Y.; Linnankivi, T.; Keski-Filppula, R.; Herva, R.; Rantala, H.; Paetau, A.; Poyhonen, M.; et al. Biallelic Variants in UBA5 Link Dysfunctional UFM1 Ubiquitin-like Modifier Pathway to Severe Infantile-Onset Encephalopathy. Am. J. Hum. Genet. 2016, 99, 683–694. [Google Scholar] [CrossRef]
- Colin, E.; Daniel, J.; Ziegler, A.; Wakim, J.; Scrivo, A.; Haack, T.B.; Khiati, S.; Denomme, A.S.; Amati-Bonneau, P.; Charif, M.; et al. Biallelic Variants in UBA5 Reveal that Disruption of the UFM1 Cascade Can Result in Early-Onset Encephalopathy. Am. J. Hum. Genet. 2016, 99, 695–703. [Google Scholar] [CrossRef] [PubMed]
- Nahorski, M.S.; Maddirevula, S.; Ishimura, R.; Alsahli, S.; Brady, A.F.; Begemann, A.; Mizushima, T.; Guzman-Vega, F.J.; Obata, M.; Ichimura, Y.; et al. Biallelic UFM1 and UFC1 mutations expand the essential role of ufmylation in brain development. Brain 2018, 141, 1934–1945. [Google Scholar] [CrossRef]
- Duan, R.; Shi, Y.; Yu, L.; Zhang, G.; Li, J.; Lin, Y.; Guo, J.; Wang, J.; Shen, L.; Jiang, H.; et al. UBA5 Mutations Cause a New Form of Autosomal Recessive Cerebellar Ataxia. PLoS ONE 2016, 11, e0149039. [Google Scholar] [CrossRef] [PubMed]
- Jing, Y.; Mao, Z.; Chen, F. UFMylation System: An Emerging Player in Tumorigenesis. Cancers 2022, 14, 3501. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Li, J.; Tillman, B.; French, B.A.; French, S.W. Ufmylation and FATylation pathways are downregulated in human alcoholic and nonalcoholic steatohepatitis, and mice fed DDC, where Mallory-Denk bodies (MDBs) form. Exp. Mol. Pathol. 2014, 97, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Gong, M.; French, B.A.; Li, J.; Tillman, B.; French, S.W. Mallory-Denk Body (MDB) formation modulates Ufmylation expression epigenetically in alcoholic hepatitis (AH) and non-alcoholic steatohepatitis (NASH). Exp. Mol. Pathol. 2014, 97, 477–483. [Google Scholar] [CrossRef]
- Mao, Z.; Ma, X.; Jing, Y.; Shen, M.; Ma, X.; Zhu, J.; Liu, H.; Zhang, G.; Chen, F. Ufmylation on UFBP1 alleviates non-alcoholic fatty liver disease by modulating hepatic endoplasmic reticulum stress. Cell Death Dis. 2023, 14, 584. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, H. Multiple organs involved in the pathogenesis of non-alcoholic fatty liver disease. Cell Biosci. 2020, 10, 140. [Google Scholar] [CrossRef]
- He, C.; Xing, X.; Chen, H.Y.; Gao, M.; Shi, J.; Xiang, B.; Xiao, X.; Sun, Y.; Yu, H.; Xu, G.; et al. UFL1 ablation in T cells suppresses PD-1 UFMylation to enhance anti-tumor immunity. Mol. Cell 2024, 84, 1120–1138.e8. [Google Scholar] [CrossRef]
- Gwinn, D.M.; Shackelford, D.B.; Egan, D.F.; Mihaylova, M.M.; Mery, A.; Vasquez, D.S.; Turk, B.E.; Shaw, R.J. AMPK phosphorylation of raptor mediates a metabolic checkpoint. Mol. Cell 2008, 30, 214–226. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Sheng, L.; Zhou, T.; Yan, L.; Loveless, R.; Li, H.; Teng, Y.; Cai, Y. Loss of Ufl1/Ufbp1 in hepatocytes promotes liver pathological damage and carcinogenesis through activating mTOR signaling. J. Exp. Clin. Cancer Res. 2023, 42, 110. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Xu, X.; Wang, Z. The Post-Translational Role of UFMylation in Physiology and Disease. Cells 2023, 12, 2543. [Google Scholar] [CrossRef] [PubMed]
- Fang, B.; Li, Z.; Qiu, Y.; Cho, N.; Yoo, H.M. Inhibition of UBA5 Expression and Induction of Autophagy in Breast Cancer Cells by Usenamine A. Biomolecules 2021, 11, 1348. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Gene (Mus Musculus) | Primer Sequence (5′-3′) |
|---|---|
| Ufl1-Forward | CTGGGACAACTGATTGATGAGAA |
| Ufl1-Reverse | AGGAAGGTCATAGGCTTTACACA |
| Ufm1-Forward | GCTGCCGTACAAAGTTCTCAG |
| Ufm1-Reverse | GTGCTTCAGGAAAACATTCCCA |
| Uba5-Forward | GAGATGAGCGACGAGGTGTTG |
| Uba5-Reverse | ACAGCGTAGGTACGGATTTTCT |
| Ufc1-Forward | CGGGTCGTGTCTGAGATCC |
| Ufc1-Reverse | GGTCCCTTCCTTGTTGGACT |
| Ddrgk1-Forward | CCCTGGGTGTATCTGGTGG |
| Ddrgk1-Reverse | CATTGTGCAGTGGTTCTCCGT |
| Ufsp1-Forward | ATCACTATGGTTGCGATGGACT |
| Ufsp1-Reverse | GCCGATCCAGTTACGGGAG |
| Ufsp2-Forward | TATCAAGAACGCACTGCGACA |
| Ufsp2-Reverse | CCACAGGTACACGGAACTGTT |
| β-actin-Forward | CTCAGGAGGAGCAATGATCTTGAT |
| β-actin-Reverse | TACCACCATGTACCCAGGCA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, M.; Zhang, C.; Zhang, Z.; Duan, Y.; Qi, S.; Zeng, Q.; Wang, J.; Zhang, J.; Jiang, Y.; Wang, Y.; et al. Systematic Analysis of UFMylation Family Genes in Tissues of Mice with Metabolic Dysfunction-Associated Steatotic Liver Disease. Genes 2025, 16, 31. https://doi.org/10.3390/genes16010031
Jiang M, Zhang C, Zhang Z, Duan Y, Qi S, Zeng Q, Wang J, Zhang J, Jiang Y, Wang Y, et al. Systematic Analysis of UFMylation Family Genes in Tissues of Mice with Metabolic Dysfunction-Associated Steatotic Liver Disease. Genes. 2025; 16(1):31. https://doi.org/10.3390/genes16010031
Chicago/Turabian StyleJiang, Mingdi, Chenlu Zhang, Zhengyao Zhang, Yingying Duan, Shuaiyong Qi, Qingyu Zeng, Jiabao Wang, Jiawen Zhang, Yu Jiang, Ying Wang, and et al. 2025. "Systematic Analysis of UFMylation Family Genes in Tissues of Mice with Metabolic Dysfunction-Associated Steatotic Liver Disease" Genes 16, no. 1: 31. https://doi.org/10.3390/genes16010031
APA StyleJiang, M., Zhang, C., Zhang, Z., Duan, Y., Qi, S., Zeng, Q., Wang, J., Zhang, J., Jiang, Y., Wang, Y., Chen, Y., & Liu, J. (2025). Systematic Analysis of UFMylation Family Genes in Tissues of Mice with Metabolic Dysfunction-Associated Steatotic Liver Disease. Genes, 16(1), 31. https://doi.org/10.3390/genes16010031