
FOXO3/Rab7-Mediated Lipophagy and Its Role in Zn-Induced Lipid Metabolism in Yellow Catfish (Pelteobagrus fulvidraco)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals and Reagents
2.2. Promoter Cloning and Plasmid Construction
2.3. Sequence Analysis and Activities Assays of Luciferase
2.4. Hepatocyte Culture and Treatments
2.5. Site Mutation Assays of FOXO3 Binding Sites on the rab7 Promoter
2.6. Zn, LD, Autophagic Vesicles, and Triglycerides Content in Yellow Catfish Hepatocytes
2.7. Quantitative Real-Time PCR (qPCR) Assay
2.8. Statistical Analysis
3. Results
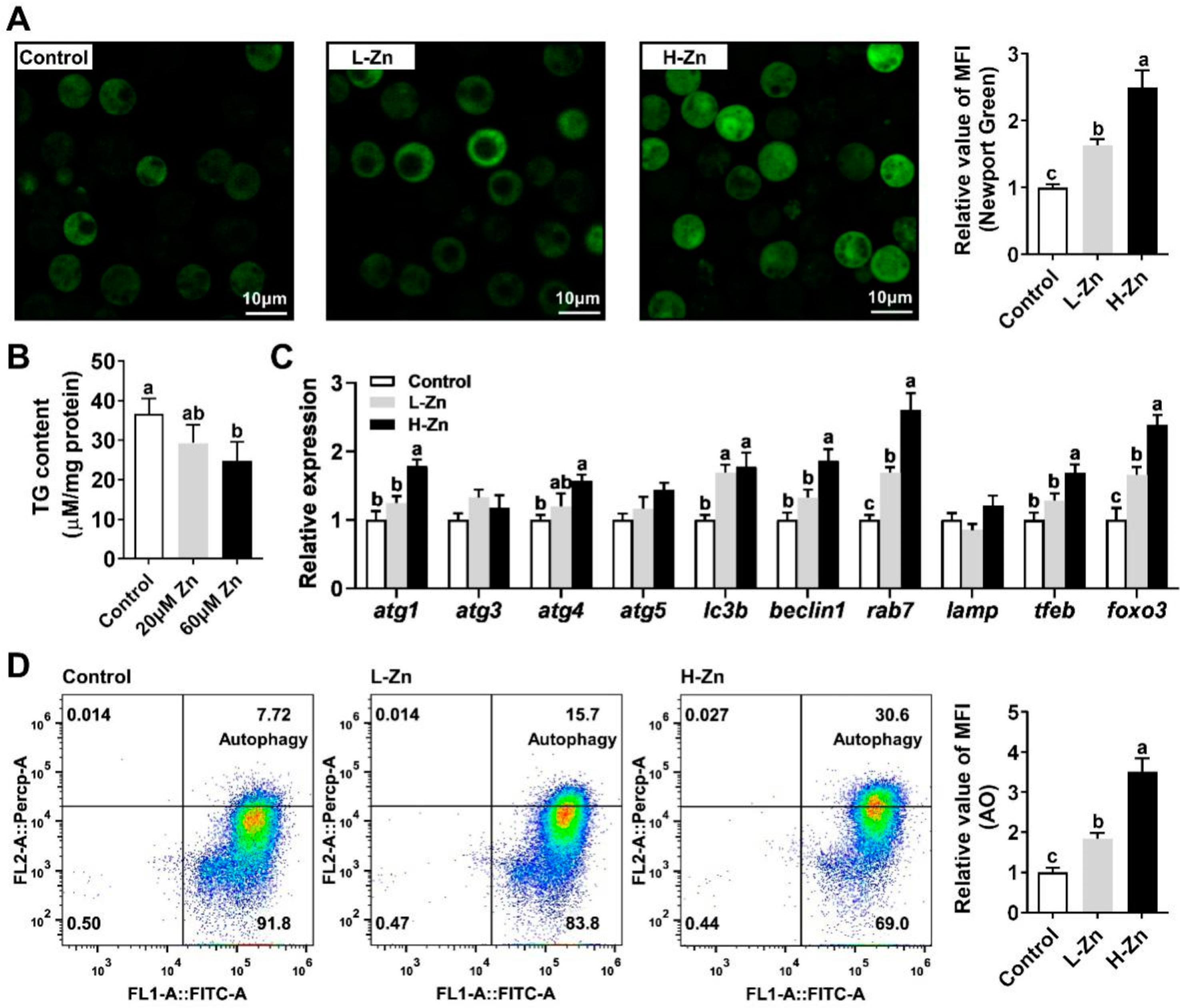
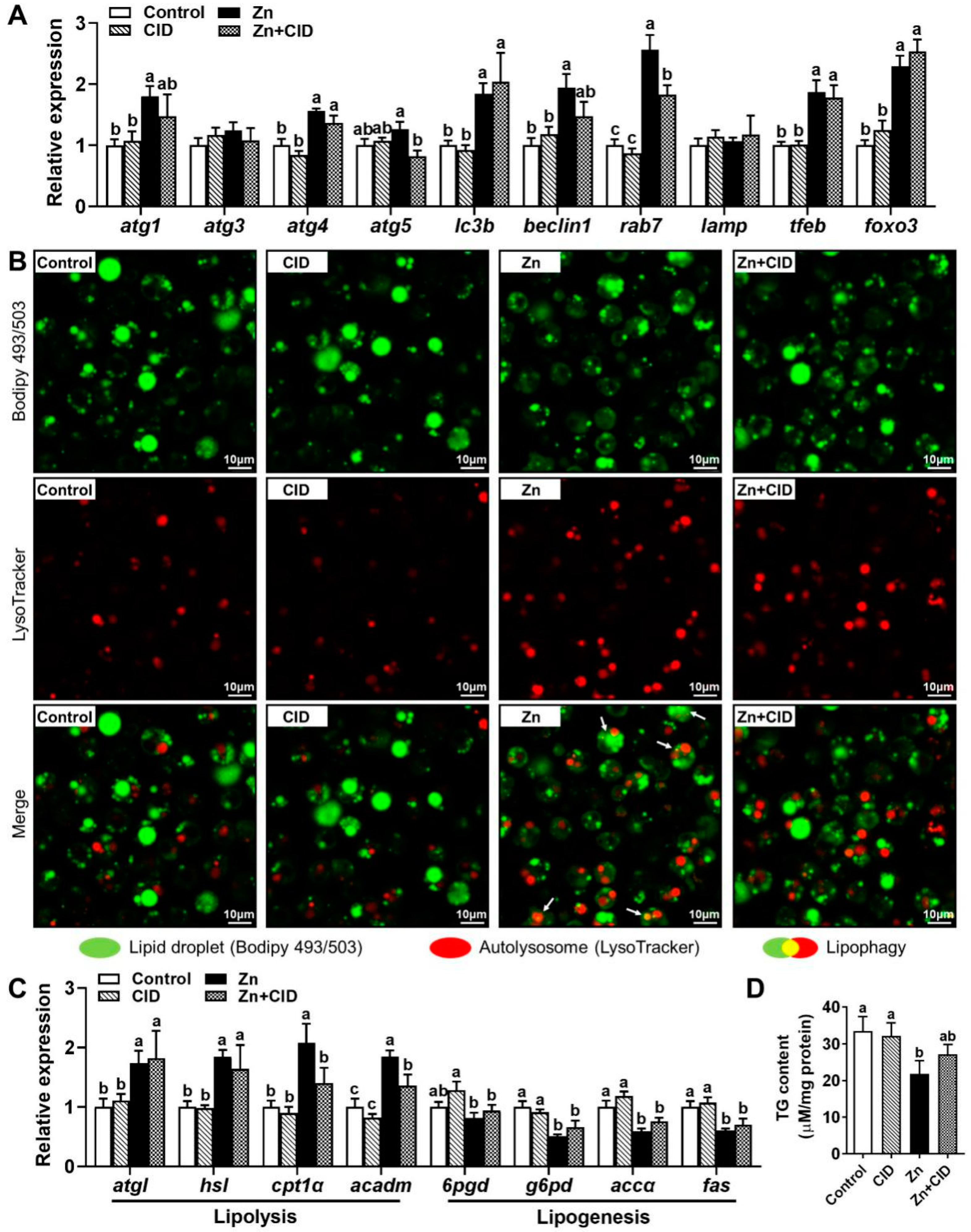
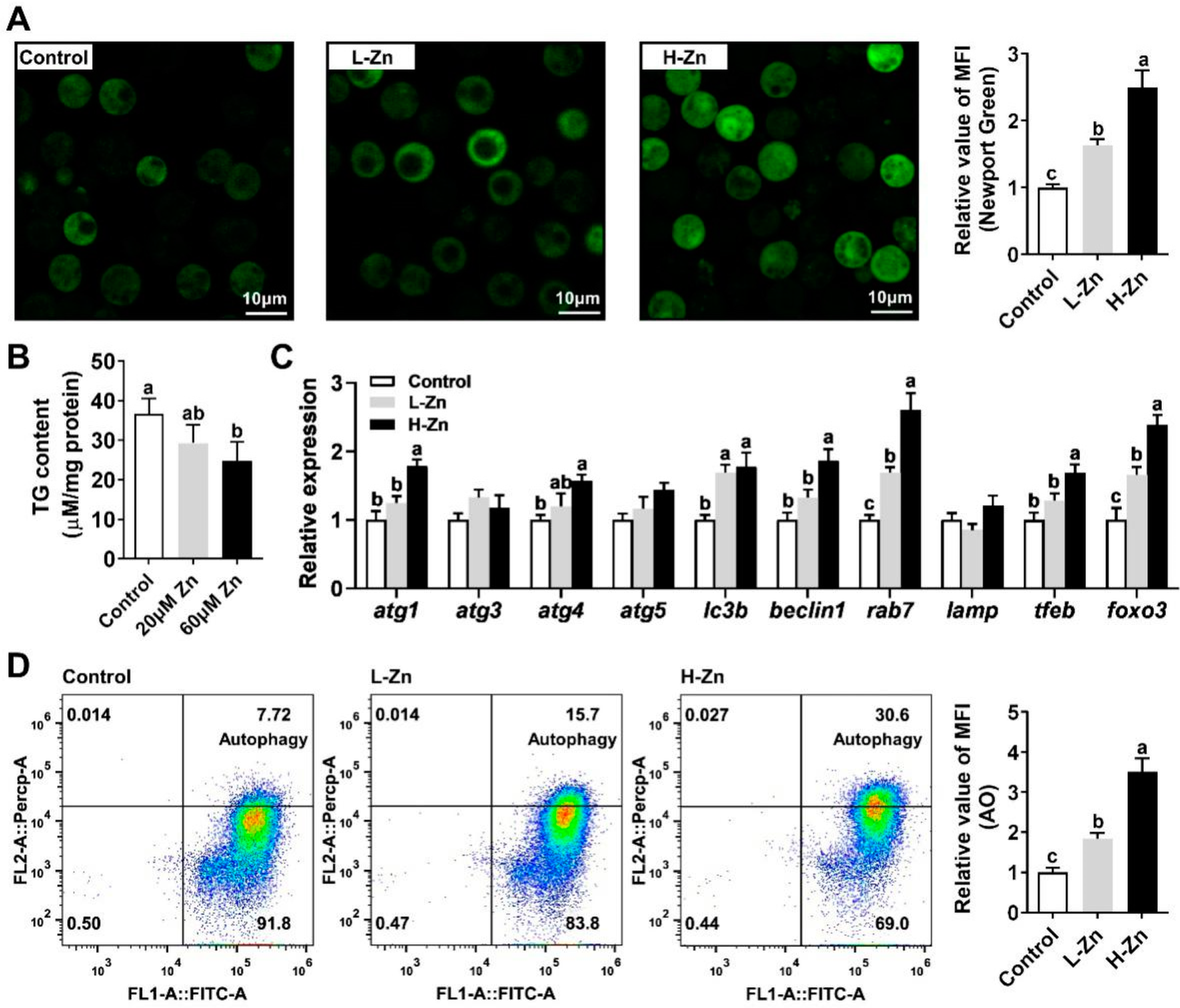
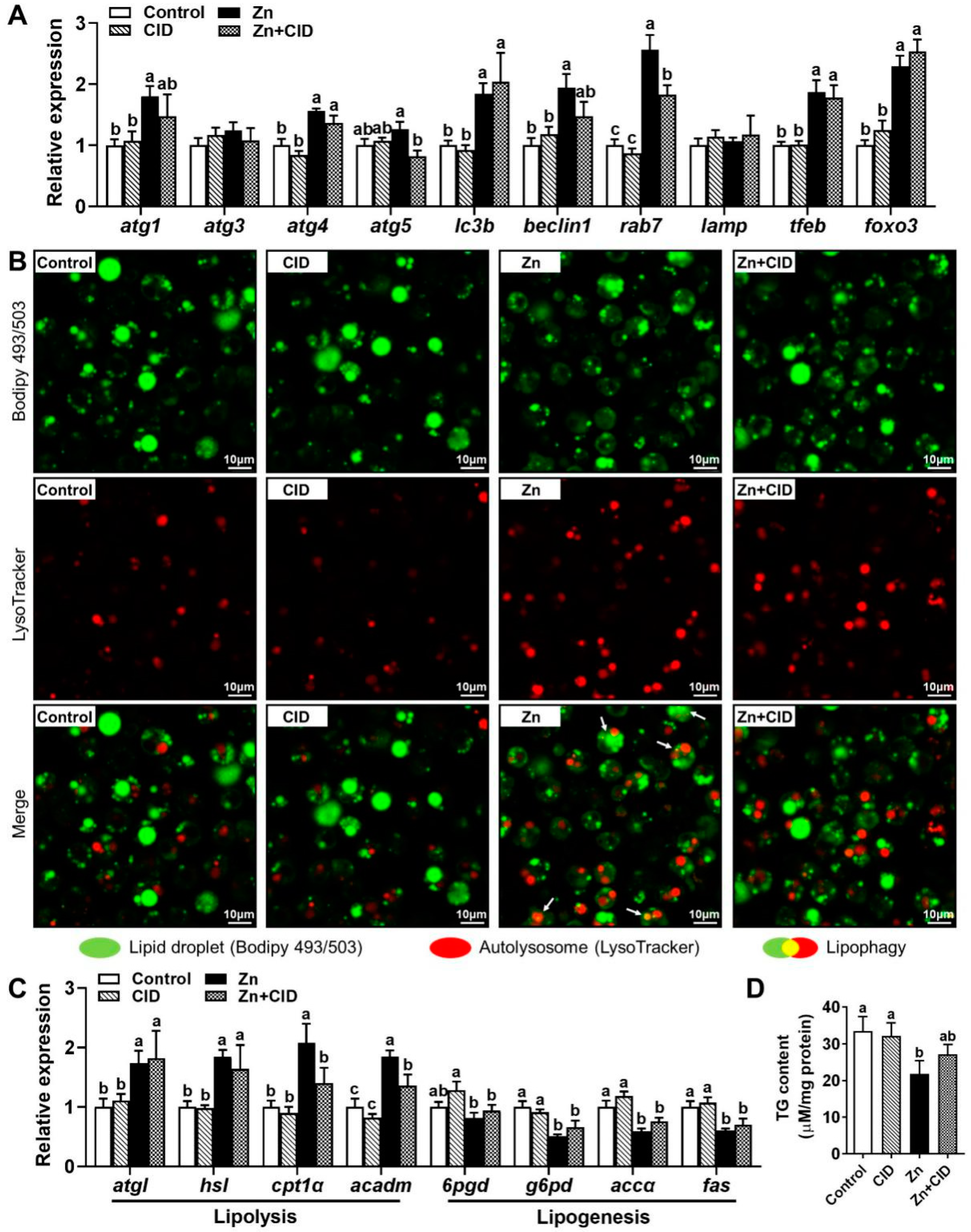
3.1. Zn Reduces TG Content in Hepatocytes by Inducing Lipophagy
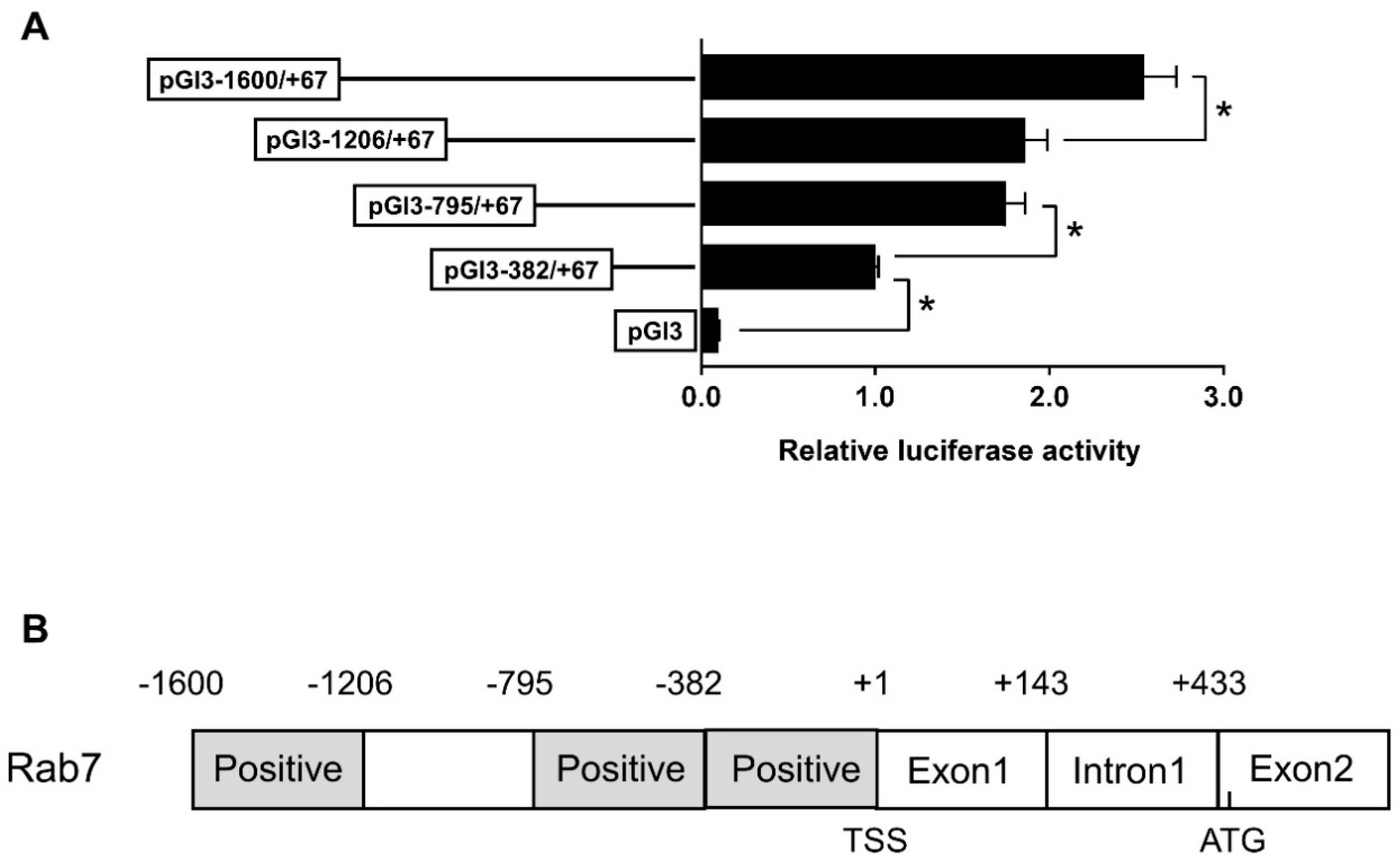
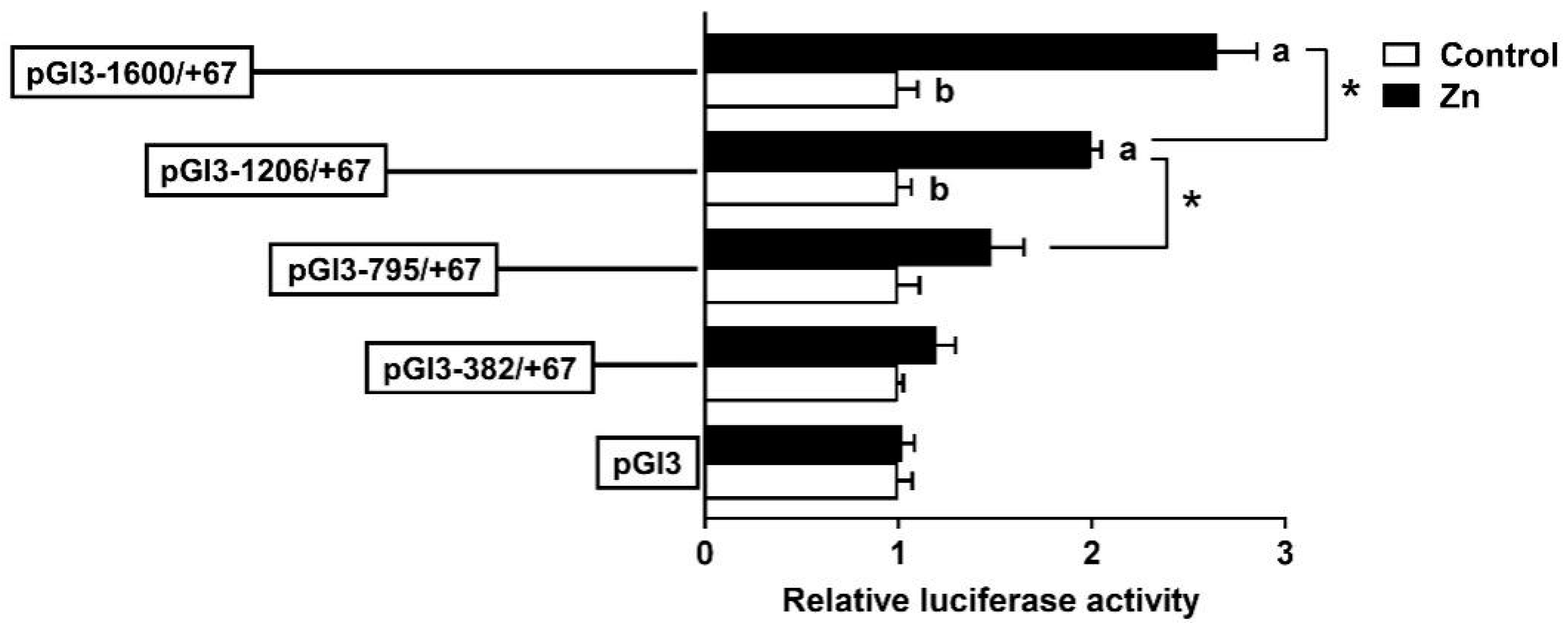
3.2. The rab7 Promoter Possesses Multiple Potential FOXO3 Binding Sites
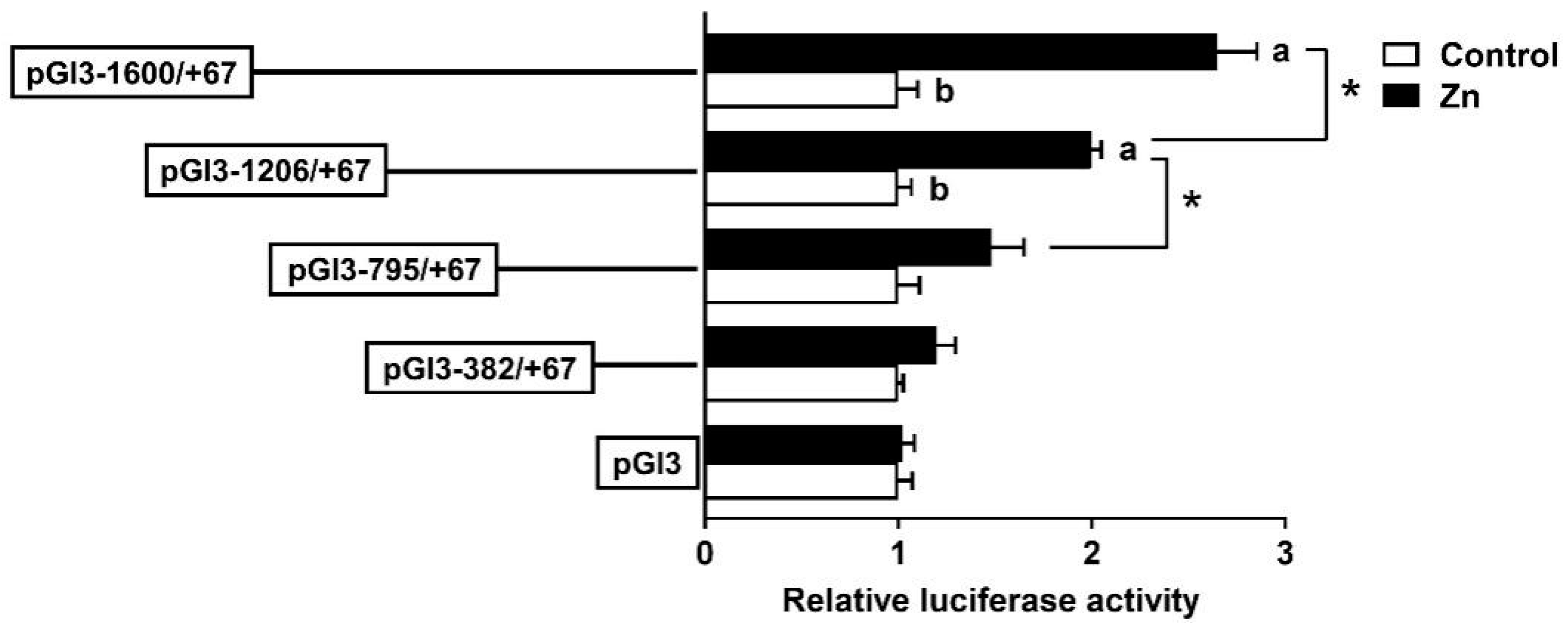
3.3. The rab7 Promoter Depends on Specific Regions to Respond to Zn Signals
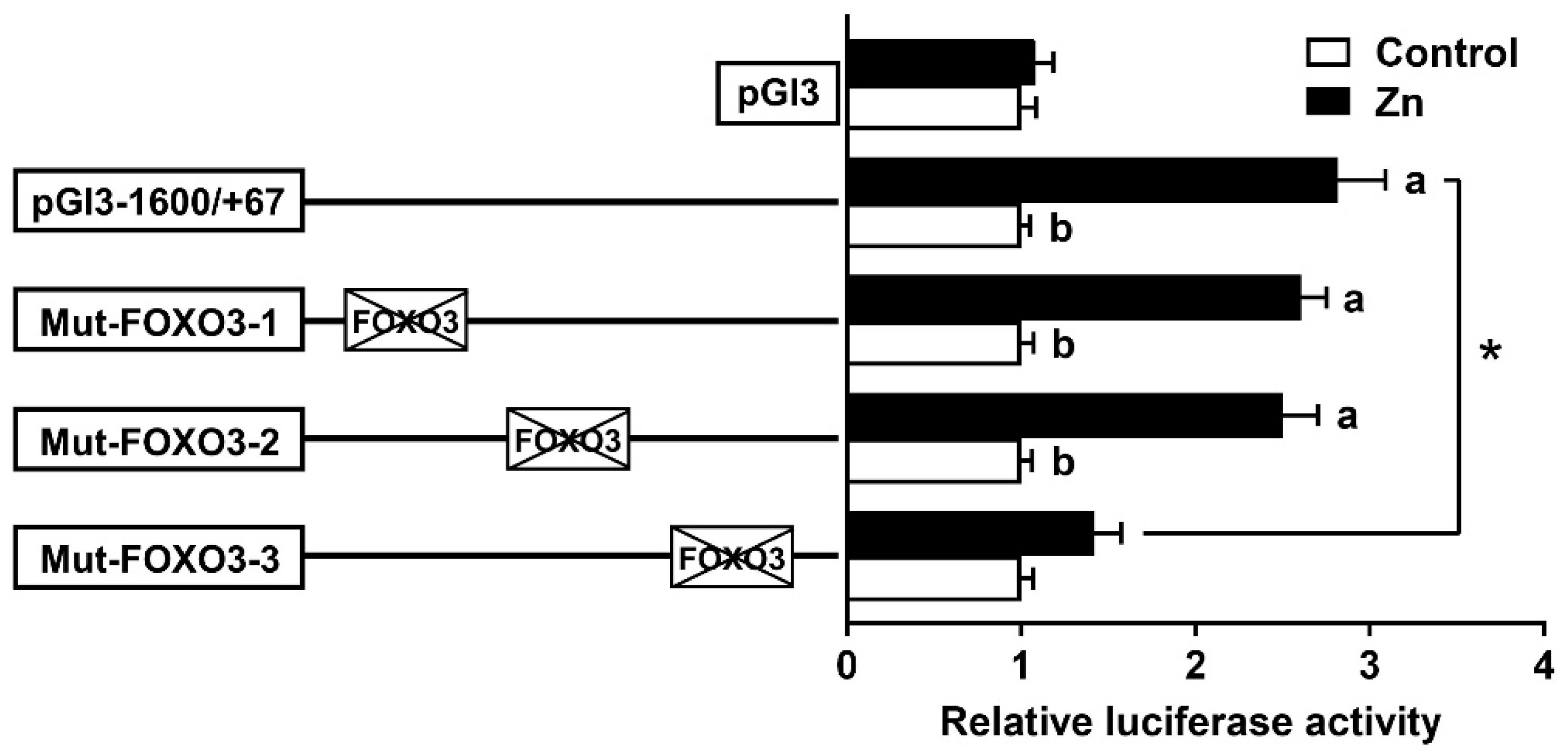
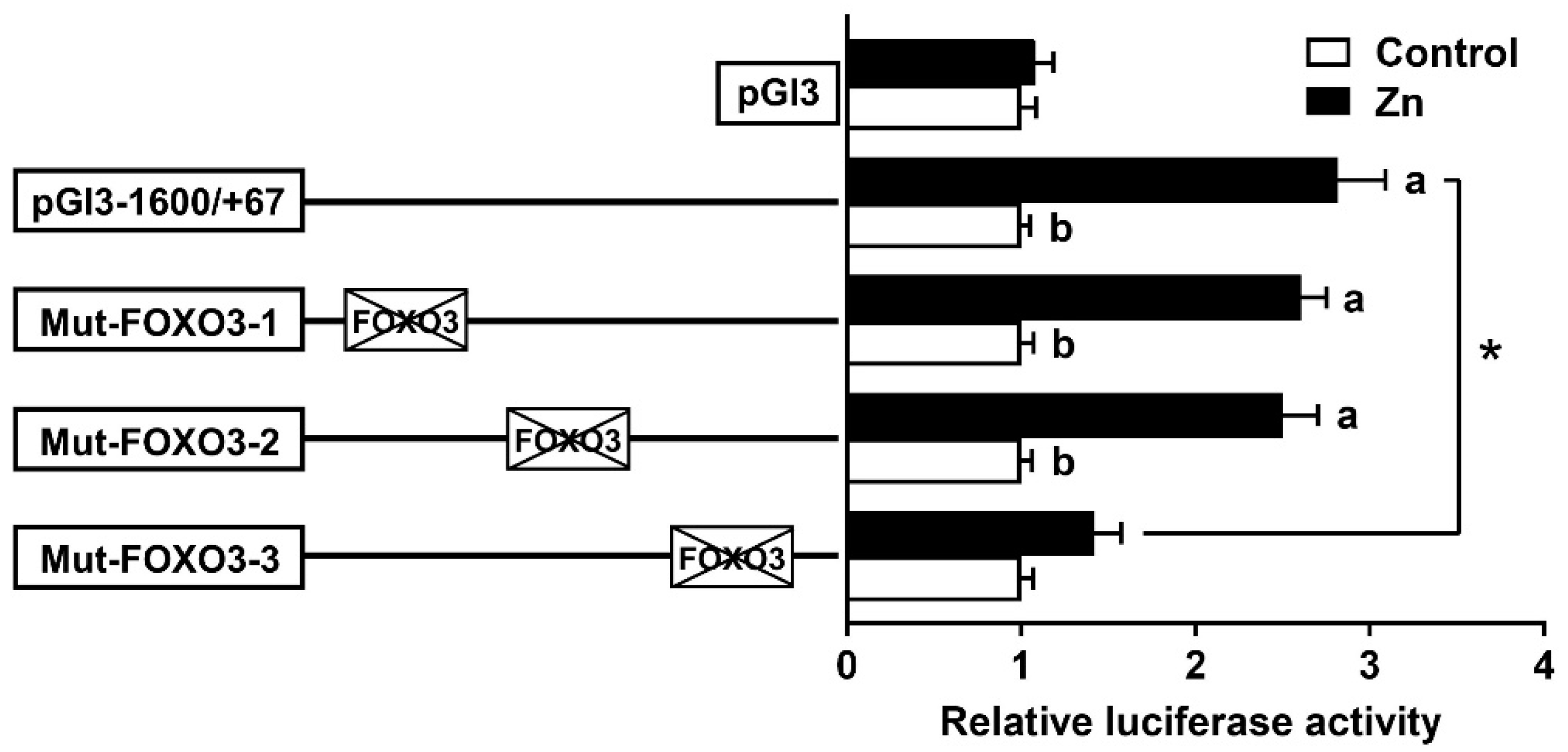
3.4. The FOXO3 Binding Site Regulated rab7 Promoter Activity
3.5. rab7 Mediated Zn-Induced Lipophagy to Reduce Lipid Accumulation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kim, K.H.; Lee, M.-S. Autophagy-a key player in cellular and body metabolism. Nat. Rev. Endocrinol. 2014, 10, 322–337. [Google Scholar] [CrossRef]
- Singh, R.; Kaushik, S.; Wang, Y.; Xiang, Y.; Novak, I.; Komatsu, M.; Tanaka, K.; Cuervo, A.M.; Czaja, M.J. Autophagy regulates lipid metabolism. Nature 2009, 458, 1131–1135. [Google Scholar] [CrossRef]
- Kaushik, S.; Cuervo, A.M. Degradation of lipid droplet-associated proteins by chaperone-mediated autophagy facilitates lipolysis. Nat. Cell Biol. 2015, 17, 759–770. [Google Scholar] [CrossRef]
- Zechner, R.; Madeo, F.; Kratky, D. Cytosolic lipolysis and lipophagy: Two sides of the same coin. Nat. Rev. Mol. Cell Biol. 2017, 18, 671–684. [Google Scholar] [CrossRef]
- Wu, K.; Zhao, T.; Hogstrand, C.; Xu, Y.-C.; Ling, S.-C.; Chen, G.-H.; Luo, Z. FXR-mediated inhibition of autophagy contributes to FA-induced TG accumulation and accordingly reduces FA-induced lipotoxicity. In Cell Communication and Signaling; Springer: Berlin/Heidelberg, Germany, 2020; Volume 18, pp. 1–16. [Google Scholar]
- Kaur, J.; Debnath, J. Autophagy at the crossroads of catabolism and anabolism. Nat. Rev. Mol. Cell Biol. 2015, 16, 461–472. [Google Scholar] [CrossRef]
- Schott, M.B.; Rozeveld, C.N.; Weller, S.G.; McNiven, M.A. Lipophagy at a glance. J. Cell Sci. 2022, 135, jcs259402. [Google Scholar] [CrossRef]
- Wu, K.; Chen, G.-H.; Hogstrand, C.; Ling, S.-C.; Wu, L.-X.; Luo, Z. Methionine-chelated Zn promotes anabolism by integrating mTOR signal and autophagy pathway in juvenile yellow catfish. J. Trace Elem. Med. Biol. 2021, 65, 126732. [Google Scholar] [CrossRef]
- Zhang, S.; Peng, X.; Yang, S.; Li, X.; Huang, M.; Wei, S.; Liu, J.; He, G.; Zheng, H.; Yang, L.; et al. The regulation, function, and role of lipophagy, a form of selective autophagy, in metabolic disorders. Cell Death Dis. 2022, 13, 132. [Google Scholar] [CrossRef]
- Lin, H.; Guo, X.; Liu, J.; Liu, P.; Mei, G.; Li, H.; Li, D.; Chen, H.; Chen, L.; Zhao, Y.; et al. Improving lipophagy by restoring Rab7 cycle: Protective effects of quercetin on ethanol-induced liver steatosis. Nutrients 2022, 14, 658. [Google Scholar] [CrossRef]
- Langemeyer, L.; Fröhlich, F.; Ungermann, C. Rab GTPase Function in endosome and lysosome biogenesis. Trends Cell Biol. 2018, 28, 957–970. [Google Scholar] [CrossRef]
- Lu, Z.; Yang, H.; Sutton, M.N.; Yang, M.; Clarke, C.H.; Liao, W.S.-L.; Bast, R.C., Jr. ARHI (DIRAS3) induces autophagy in ovarian cancer cells by downregulating the epidermal growth factor receptor, inhibiting PI3K and Ras/MAP signaling and activating the FOXO3a-mediated induction of Rab7. Cell Death Differ. 2014, 21, 1275–1289. [Google Scholar] [CrossRef]
- Niu, J.; Yan, T.; Guo, W.; Wang, W.; Ren, T.; Huang, Y.; Zhao, Z.; Yu, Y.; Chen, C.; Huang, Q.; et al. The COPS3-FOXO3 positive feedback loop regulates autophagy to promote cisplatin resistance in osteosarcoma. Autophagy 2023, 19, 1693–1710. [Google Scholar] [CrossRef]
- Wei, C.C.; Luo, Z.; Hogstrand, C.; Xu, Y.H.; Wu, L.X.; Chen, G.H.; Pan, Y.X.; Song, Y.F. Zn reduces hepatic lipid deposition and activates lipophagy via Zn2+/MTF-1/PPARα and Ca2+/CaMKKβ/AMPK pathways. FASEB J. 2018, 32, 6666–6680. [Google Scholar] [CrossRef]
- Ranasinghe, P.; Wathurapatha, W.S.; Ishara, M.H.; Jayawardana, R.; Galappatthy, P.; Katulanda, P.; Constantine, G.R. Effects of Zn supplementation on serum lipids: A systematic review and meta-analysis. Nutr. Metab. 2015, 12, 26. [Google Scholar] [CrossRef]
- Zhang, J.J.; Hao, J.J.; Zhang, Y.R.; Wang, Y.L.; Li, M.Y.; Miao, H.L.; Zou, X.J.; Liang, B. Zn Mediates the SREBP-SCD axis to regulate lipid metabolism in Caenorhabditis elegans. J. Lipid Res. 2017, 58, 1845–1854. [Google Scholar] [CrossRef]
- Olechnowicz, J.; Tinkov, A.; Skalny, A.; Suliburska, J. Zn status is associated with inflammation, oxidative stress, lipid, and glucose metabolism. J. Physiol. Sci. 2018, 68, 19–31. [Google Scholar] [CrossRef]
- Wu, K.; Luo, Z.; Hogstrand, C.; Chen, G.-H.; Wei, C.-C.; Li, D.-D. Zn stimulates the phospholipids biosynthesis via the pathways of oxidative and endoplasmic reticulum stress in the intestine of freshwater teleost yellow catfish. Environ. Sci. Technol. 2018, 52, 9206–9214. [Google Scholar] [CrossRef]
- Liu, Y.-G.; Chen, Y. High-efficiency thermal asymmetric interlaced PCR for amplification of unknown flanking sequences. Biotechniques 2007, 43, 649–656. [Google Scholar] [CrossRef]
- Chen, S.-W.; Wu, K.; Lv, W.-H.; Chen, F.; Song, C.-C.; Luo, Z. Functional analysis of two Zn (Zn) transporters (ZIP3 and ZIP8) promoters and their distinct response to MTF1 and RREB1 in the regulation of Zn metabolism. Int. J. Mol. Sci. 2020, 21, 6135. [Google Scholar] [CrossRef]
- Gao, H.; Fan, X.; Wu, Q.-C.; Chen, C.; Xiao, F.; Wu, K. Structural and functional analysis of SHP promoter and its transcriptional response to FXR in Zn-induced changes to lipid metabolism. Int. J. Mol. Sci. 2022, 23, 6523. [Google Scholar] [CrossRef]
- Wang, S.; Shi, X.; Wei, S.; Ma, D.; Oyinlade, O.; Lv, S.-Q.; Ying, M.; Zhang, Y.A.; Claypool, S.M.; Watkins, P.; et al. Krüppel-like factor 4 (KLF4) induces mitochondrial fusion and increases spare respiratory capacity of human glioblastoma cells. J. Biol. Chem. 2018, 293, 6544–6555. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Inoue, T.; Inoue, M.; Murae, M.; Fukasawa, M.; Kaneko, M.K.; Kato, Y.; Noguchi, K. SARS-CoV-2 spike protein mutation at cysteine-488 impairs its golgi localization and intracellular S1/S2 processing. Int. J. Mol. Sci. 2022, 23, 15834. [Google Scholar] [CrossRef]
- Vandesompele, J.; Preter, K.D.; Pattyn, F.; Poppe, B.; Roy, N.V.; Paepe, A.D.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034. [Google Scholar]
- Livak, K.; Schmittgen, T. Analysis of relative gene expression data using real-Time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 4022–4408. [Google Scholar] [CrossRef]
- Han, S.L.; Qian, Y.C.; Limbu, S.M.; Wang, J.; Chen, L.Q.; Zhang, M.L.; Du, Z.Y. Lipolysis and lipophagy play individual and interactive roles in regulating triacylglycerol and cholesterol homeostasis and mitochondrial form in zebrafish. BBA-Mol. Cell Biol. L 2021, 1866, 158988. [Google Scholar] [CrossRef]
- Liu, K.; Czaja, M.J. Regulation of lipid stores and metabolism by lipophagy. Cell Death Differ. 2013, 20, 3–11. [Google Scholar] [CrossRef]
- Pabon, M.A.; Ma, K.C.; Choi, A.M.K. Autophagy and obesity-related lung disease. Am. J. Respir. Cell Mol. Biol. 2016, 54, 636–646. [Google Scholar] [CrossRef]
- Xu, T.; Nicolson, S.; Denton, D.; Kumar, S. Distinct requirements of Autophagy-related genes in programmed cell death. Cell Death Differ. 2015, 22, 1792–1802. [Google Scholar] [CrossRef]
- Liuzzi, J.P.; Guo, L.; Yoo, C.; Stewart, T.S. Zn and autophagy. Biometals 2014, 27, 1084–1096. [Google Scholar] [CrossRef]
- Wei, X.; Hogstrand, C.; Chen, G.; Lv, W.; Song, Y.; Xu, Y.; Luo, Z. Zn induces lipophagy via the deacetylation of beclin1 and alleviates Cu-induced lipotoxicity at their environmentally relevant concentrations. Environ. Sci. Technol. 2021, 55, 4943–4953. [Google Scholar] [CrossRef]
- Bucci, C.; Thomsen, P.; Nicoziani, P.; McCarthy, J.; van Deurs, B.; Okamoto, Y.; Shikano, S.; Gruenberg, M.E.J.E.; Hegedűs, K.; Takáts, S.; et al. Rab7: A key to lysosome biogenesis. Mol. Biol. Cell 2000, 11, 467–480. [Google Scholar] [CrossRef]
- Schroeder, B.; Schulze, R.J.; Weller, S.G.; Sletten, A.C.; Casey, C.A.; McNiven, M.A. The small GTPase Rab7 as a central regulator of hepatocellular lipophagy. Hepatology 2015, 61, 1896–1907. [Google Scholar] [CrossRef]
- Mammucari, C.; Milan, G.; Romanello, V.; Masiero, E.; Rudolf, R.; Del Piccolo, P.; Burden, S.J.; Di Lisi, R.; Sandri, C.; Zhao, J.; et al. FOXO3 controls autophagy in skeletal muscle in vivo. Cell Metab. 2007, 6, 458–471. [Google Scholar] [CrossRef]
- Zhao, J.; Brault, J.J.; Schild, A.; Cao, P.; Sandri, M.; Schiaffino, S.; Lecker, S.H.; Goldberg, A.L. FoxO3 coordinately activates protein degradation by the autophagic/lysosomal and proteasomal pathways in atrophying muscle cells. Cell Metab. 2007, 6, 472–483. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, H.; Li, X.; Wang, Q.; Yan, M.; Zhang, H.; Zhao, T.; Zhang, N.; Zhang, P.; Peng, L.; et al. Formononetin alleviates hepatic steatosis by facilitating TFEB-mediated lysosome biogenesis and lipophagy. J. Nutr. Biochem. 2019, 73, 108214. [Google Scholar] [CrossRef]
- Roy, A.L.; Singer, D.S. Core promoters in transcription: Old problem, new insights. Trends Biochem. Sci. 2015, 40, 165–171. [Google Scholar] [CrossRef]
- Basehoar, A.D.; Zanton, S.J.; Pugh, B. Identification and distinct regulation of yeast TATA Box-containing genes. Cell 2004, 116, 699–709. [Google Scholar] [CrossRef]
- Kim, Y.S.; Lee, H.M.; Kim, J.K.; Yang, C.S.; Kim, T.S.; Jung, M.; Jin, H.S.; Kim, S.; Jang, J.; Oh, G.T.; et al. PPAR-α activation mediates innate host defense through induction of TFEB and lipid catabolism. J. Immunol. 2017, 198, 3283–3295. [Google Scholar] [CrossRef]
- Dorayappan, K.D.P.; Wanner, R.; Wallbillich, J.J.; Saini, U.; Zingarelli, R.; Suarez, A.A.; Cohn, D.E.; Selvendiran, K. Hypoxia-induced exosomes contribute to a more aggressive and chemoresistant ovarian cancer phenotype: A novel mechanism linking STAT3/Rab proteins. Oncogene 2018, 37, 3806–3821. [Google Scholar] [CrossRef]
- Tsai, M.-S.; Lee, H.-M.; Huang, S.-C.; Sun, C.-K.; Chiu, T.-C.; Chen, P.-H.; Lin, Y.-C.; Hung, T.-M.; Lee, P.-H.; Kao, Y.-H. Nerve growth factor induced farnesoid X receptor upregulation modulates autophagy flux and protects hepatocytes in cholestatic livers. Arch. Biochem. Biophys. 2020, 682, 108281. [Google Scholar] [CrossRef]
- Tang, H.; Huang, H.; Wang, D.; Li, P.; Tian, Z.; Li, D.; Wang, S.; Ma, R.; Xia, T.; Wang, A. TFEB ameliorates autophagy flux disturbance induced by PBDE-47 via up-regulating autophagy-lysosome fusion. J. Hazard. Mater. 2022, 430, 128483. [Google Scholar] [CrossRef]
- Settembre, C.; Ballabio, A. TFEB regulates autophagy: An integrated coordination of cellular degradation and recycling processes. Autophagy 2011, 7, 1379–1381. [Google Scholar] [CrossRef]
- Yan, S. Role of TFEB in Autophagy and the pathogenesis of liver diseases. Biomolecules 2022, 12, 672. [Google Scholar] [CrossRef]
- Barbosa, A.D.; Siniossoglou, S. Function of lipid droplet-organelle interactions in lipid homeostasis. BBA-Mol Cell Res. 2017, 1864, 1459–1468. [Google Scholar] [CrossRef]
- Lam, T.; Kulp, D.V.; Wang, R.; Lou, Z.; Taylor, J.; Rivera, C.E.; Yan, H.; Zhang, Q.; Wang, Z.; Zan, H.; et al. Small molecule inhibition of Rab7 impairs B cell class switching and plasma cell survival to dampen the autoantibody response in murine lupus. J. Immunol. 2016, 197, 3792–3805. [Google Scholar] [CrossRef]
- Kallenborn-Gerhardt, W.; Möser, C.V.; Lorenz, J.E.; Steger, M.; Heidler, J.; Scheving, R.; Petersen, J.; Kennel, L.; Flauaus, C.; Lu, R. Rab7-a novel redox target that modulates inflammatory pain processing. Pain 2017, 158, 1354–1365. [Google Scholar] [CrossRef]
- Ritter, J.L.; Zhu, Z.; Thai, T.C.; Mahadevan, N.R.; Mertins, P.; Knelson, E.H.; Piel, B.P.; Han, S.; Jaffe, J.D.; Carr, S.A.; et al. Phosphorylation of Rab7 by TBK1/IKKϵ regulates innate immune sgnaling in triple-negative breast cancer. Cancer Res. 2020, 80, 44–56. [Google Scholar] [CrossRef]
- Sun, M.; Zhang, W.; Bi, Y.; Xu, H.; Abudureyimu, M.; Peng, H.; Zhang, Y.; Ren, J. NDP52 protects against myocardial infarction-provoked cardiac anomalies through promoting autophagosome-lysosome fusionvia recruiting TBK1 and RAB7. Antioxid. Redox Signal. 2022, 36, 16–18. [Google Scholar] [CrossRef]
- Zhu, L.; Yuan, G.; Wang, X.; Zhao, T.; Hou, L.; Li, C.; Jiang, X.; Zhang, J.; Zhao, X.; Pei, C.; et al. Molecular characterization of Rab7 and its involvement in innate immunity in red swamp crayfish Procambarus clarkia. Fish Shellfish. Immunol. 2022, 127, 318–328. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, Q.; Guo, X.K.; Zhang, X.; Zhou, Z. FOXO3a regulates lipid accumulation and adipocyte inflammation in adipocytes through autophagy: Role of FOXO3a in obesity. Inflamm. Res. 2021, 70, 591–603. [Google Scholar] [CrossRef]
- He, C.; Klionsky, D.J. Regulation mechanisms and signaling pathways of autophagy. Annu. Rev. Genet. 2009, 43, 67–93. [Google Scholar] [CrossRef]
- Kang, X.; Zhong, W.; Liu, J.; Song, Z.; McClain, C.J.; Kang, Y.J.; Zhou, Z. Zn supplementation reverses alcohol-induced steatosis in mice through reactivating hepatocyte nuclear factor-4α and peroxisome proliferator-activated receptor-α. Hepatology 2009, 50, 1241–1250. [Google Scholar] [CrossRef]
- Wu, K.; Huang, C.; Shi, X.; Chen, F.; Xu, Y.-H.; Pan, Y.-X.; Luo, Z.; Liu, X. Role and mechanism of the AMPK pathway in waterborne Zn exposure influencing the hepatic energy metabolism of Synechogobius hasta. Sci. Rep. 2016, 6, 38716. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, F.; Chen, C.; Zhang, W.; Wang, J.; Wu, K. FOXO3/Rab7-Mediated Lipophagy and Its Role in Zn-Induced Lipid Metabolism in Yellow Catfish (Pelteobagrus fulvidraco). Genes 2024, 15, 334. https://doi.org/10.3390/genes15030334
Xiao F, Chen C, Zhang W, Wang J, Wu K. FOXO3/Rab7-Mediated Lipophagy and Its Role in Zn-Induced Lipid Metabolism in Yellow Catfish (Pelteobagrus fulvidraco). Genes. 2024; 15(3):334. https://doi.org/10.3390/genes15030334
Chicago/Turabian StyleXiao, Fei, Chuan Chen, Wuxiao Zhang, Jiawei Wang, and Kun Wu. 2024. "FOXO3/Rab7-Mediated Lipophagy and Its Role in Zn-Induced Lipid Metabolism in Yellow Catfish (Pelteobagrus fulvidraco)" Genes 15, no. 3: 334. https://doi.org/10.3390/genes15030334
APA StyleXiao, F., Chen, C., Zhang, W., Wang, J., & Wu, K. (2024). FOXO3/Rab7-Mediated Lipophagy and Its Role in Zn-Induced Lipid Metabolism in Yellow Catfish (Pelteobagrus fulvidraco). Genes, 15(3), 334. https://doi.org/10.3390/genes15030334