Abstract
Tarsiger indicus (Vieillot, 1817), the White-browed Bush Robin, is a small passerine bird widely distributed in Asian countries. Here, we successfully sequenced its mitogenome using the Illumina Novaseq 6000 platform (Illumina, San Diego, CA, USA) for PE 2 × 150 bp sequencing. Combined with other published mitogenomes, we conducted the first comprehensive comparative mitogenome analysis of Muscicapidae birds and reconstructed the phylogenetic relationships between Muscicapidae and related groups. The T. indicus mitogenome was 16,723 bp in size, and it possessed the typical avian mitogenome structure and organization. Most PCGs of T. indicus were initiated strictly with the typical start codon ATG, while COX1 and ND2 were started with GTG. RSCU statistics showed that CUA, CGA, and GCC were relatively high frequency in the T. indicus mitogenome. T. cyanurus and T. indicus shared very similar mitogenomic features. All 13 PCGs of Muscicapidae mitogenomes had experienced purifying selection. Specifically, ATP8 had the highest rate of evolution (0.13296), whereas COX1 had the lowest (0.01373). The monophylies of Muscicapidae, Turdidae, and Paradoxornithidae were strongly supported. The clade of ((Muscicapidae + Turdidae) + Sturnidae) in Passeriformes was supported by both Bayesian Inference and Maximum likelihood analyses. The latest taxonomic status of many passerine birds with complex taxonomic histories were also supported. For example, Monticola gularis, T. indicus, and T. cyanurus were allocated to Turdidae in other literature; our phylogenetic topologies clearly supported their membership in Muscicapidae; Paradoxornis heudei, Suthora webbiana, S. nipalensis, and S. fulvifrons were formerly classified into Muscicapidae; we supported their membership in Paradoxornithidae; Culicicapa ceylonensis was originally classified as a member of Muscicapidae; our results are consistent with a position in Stenostiridae. Our study enriches the genetic data of T. indicus and provides new insights into the molecular phylogeny and evolution of passerine birds.
1. Introduction
Passerines (Aves: Passeriformes) include a large number of species and are adapted to various ecological environments. The latest data show that the group has 145 families and 6695 species, accounting for 60% of all bird species; moreover, Muscicapidae is the third-largest family after Tyrannidae and Thraupidae, with 351 species from 53 genera [1]. Tarsiger indicus (Vieillot, 1817) (Figure 1), the White-browed Bush Robin, is a small Muscicapidae bird widely distributed in Asian countries, including India, Nepal, Bhutan, Myanmar, Vietnam, and China [2]. In China, T. indicus is found in Sichuan, Gansu, Shanxi, Hubei, Yunnan, Tibet, and Taiwan [3,4,5]. It generally inhabits the coniferous forests and the mixed broadleaf–conifer forests between alpine rock valleys at altitudes of 2440–4270 m above sea level in western China; in addition, it also inhabits the bottom shrubland of dense forests at altitudes of 2300–3200 m above sea level in Taiwan Island of China. In the past, the White-browed Bush Robin has been divided into three subspecies, including T. indicus indicus, T. i. yunnanensis, and T. i. formosanus [3]. Recently, an integrative taxonomic investigation found the Taiwan endemic T. i. formosanus to be distinctive in genetics, song, and morphology from T. i. indicus and T. i. yunnanensis of the Sino-Himalayan mountains [6]. In view of this, the T. i. formosanus subspecies has been suggested to be upgraded to the species T. formosanus, named the Taiwan Bush Robin [6,7]. In addition, T. indicus has been included in the updated List of Terrestrial Wild Animals of Important Ecological, Scientific, and Social Value in China [8]. Due to its wide geographical distribution and large population size, the conservation status of T. indicus is Least Concern in both the IUCN Red List of Threatened Species [2] and the Red List of China’s Vertebrates [9].

Figure 1.
Reference image of adult T. indicus. The photo was taken by Taihu Hu on 20 February 2022 in Yingjing County, Ya’an City, Sichuan Province, China.
Vertebrate mitochondrial genomes (mitogenomes) are circular, typically 14,000–20,000 bp, and contain 13 protein-coding genes (PCGs), two ribosomal RNA (rRNAs), 22 transfer RNA genes (tRNAs), and one large non-coding D-loop region [10,11]. The mitogenome has been extensively used in population genetics, population dynamics, and adaptive evolution studies of various animal groups [12,13,14,15,16], particularly in phylogenetic reconstruction among animal species [14,16,17,18,19]. It is worth emphasizing that mitochondrial genomes are more reliable in phylogenetic reconstruction than a single mitochondrial gene [20,21,22]. However, the mitogenomes of the Muscicapidae family, a complex lineage of passerines, has been studied very little. So far, complete mitochondrial genomes of only 24 species (ca. 7% of the overall clade) from 15 genera (ca. 28%) within Muscicapidae family have published in the GenBank database (Table 1), mainly focusing on simple mitogenomic descriptions [23,24,25,26,27,28,29].

Table 1.
List of 41 species used for the comparative mitogenomic analyses and the mitogenomic phylogenetic analyses in this study.
Genetic data on T. indicus are currently rare. In the GenBank database, only 39 nucleotide sequences have been uploaded as of August 2023, including 16 sequences of mitochondrial Cytb and ND2 genes. An accurate understanding of phylogeny is an important prerequisite for many studies of ecology and evolution [6,48]. However, in terms of phylogenetic status, T. indicus was previously placed into the genus Luscinia [49] and is now still placed into the Turdidae family in some publications [50].
In order to better understand the mitogenome characteristics and the phylogenetic relationship of T. indicus, we sequenced its mitochondrial genome through high-throughput sequencing technology here. Combined with other published data, we conduct the first comprehensive comparative mitogenome analysis of Muscicapidae birds and reconstruct the phylogenetic relationships between Muscicapidae and related groups using a mitogenomic approach.
2. Materials and Methods
2.1. Materials
A subadult window victim, which was found dead, was collected from Yingjing Area of the Giant Panda National Park, Scihuan Province, China (29°33′39.50″ N, 102°51′4.10″ E, 2428 m above sea level) on 30 July 2022, and it was identified as T. indicus by morphological characters and mitochondrial Cytb blast. The extraction of genomic DNA from a pectoral muscle was carried out using the Rapid Animal Genomic DNA Isolation Kit (Sangon Biotech Co., Ltd., Shanghai, China), according to the manufacturer’s protocol. The specimen and its DNA were deposited at the Chengdu Research Base of Giant Panda Breeding (Dr. Jiabin Liu, jiabin_liu2013@126.com) with the voucher number PB2022027.
2.2. Mitogenome Sequencing, Assembly, and Annotation
With the assistance of Sangon Biotech Co., Ltd. (Shanghai, China), we sequenced the mitochondrial genome through a high-throughput sequencing technique. Library preparation, mitogenome sequencing, and mitogenome assembly were performed as previously described [51]. Mitogenome annotations were implemented using MITOS WebServer (http://mitos2.bioinf.uni-leipzig.de/index.py, accessed on 15 August 2023) [52] and MitoAnnotator (http://mitofish.aori.u-tokyo.ac.jp/annotation/input/, accessed on 15 August 2023) [53]. Based on their proposed cloverleaf secondary structures and anticodon sequences, the tRNAs were rechecked using ARWEN online services (http://130.235.244.92/ARWEN/, accessed on 15 August 2023) [54]. The mitogenome visualization map was generated using Chloroplot (https://irscope.shinyapps.io/Chloroplot/, accessed on 18 August 2023) [55].
2.3. Comparative Mitogenomic Analyses
The complete mitogenome of T. indicus and 24 other Muscicapidae birds belonging to 15 genera were used for comparative mitogenomic analyses (Table 1). The 13 PCGs, two rRNAs, and whole mitogenomes were aligned in batches with MAFFT v7.505 [56]. Nucleotide composition and relative synonymous codon usage (RSCU) were calculated using MEGA v11.0.9 [57]. Nucleotide composition biases were determined from the formulas AT-skew = (A − T)/(A + T) and GC-skew = (G − C)/(G + C). The nucleotide diversity (Pi), the non-synonymous substitution rate (Ka), and the synonymous substitution rate (Ks) were calculated using DnaSP v6.12.03 [58].
Data visualization was performed using OmicStudio tools (https://www.omicstudio.cn/tool, accessed on 25 August 2023) [59].
2.4. Mitogenomic Phylogenetic Analyses
Two rRNAs and 13 PCGs of T. indicus and 40 other Passeriformes birds belonging to 26 genera and seven families were used for mitogenomic phylogenetic analyses (Table 1). The taxonomy of all birds is based on the IOC World Bird List v13.2 [1]. Pitta sordida (Passeriformes: Pittidae) was used as an outgroup based on its well-documented distant phylogenetic position from the ingroup [60,61,62]. Two rRNA sequences were aligned in batches with MAFFT v7.505 [56] using ‘–auto’ strategy and normal alignment mode, and 13 PCGs sequences were aligned in batches using the codon-aware program MACSE v2.06 [63], which preserves reading frame and allows incorporation of sequencing errors or sequences with frameshifts. Ambiguously aligned fragments of these 15 alignments were removed in batches using Gblocks v0.91b [64] with the following parameter settings: minimum number of sequences for a conserved/flank position (22/22), maximum number of contiguous non-conserved positions (8), minimum length of a block (10), allowed gap positions (with half). The 15 alignments were eventually concatenated into one multi-gene dataset consisting of a 13,893 bp sequence using PhyloSuite v1.2.3 [65]. The concatenated multi-gene dataset was used to clarify the phylogeny using Bayesian Inference (BI) and Maximum Likelihood (ML) methods. A best-fit partition model (edge-linked) was selected by ModelFinder v2.2.0 [66] using a BIC criterion, and the results are shown in Table S1. BI phylogenies were inferred using MrBayes v3.2.6 [67] under a partition model (2 parallel runs, ten million generations, sampling every one thousand generations), in which the initial 25% of sampled data were discarded as burn-in. ML phylogenies were inferred using IQ-TREE v2.2.0 [68] under an edge-linked partition model for one hundred thousand ultrafast [69] bootstraps.
High-quality figures of phylogenetic trees were produced using FigTree v.1.4.4 (http://tree.bio.ed.ac.uk/software/figtree/, accessed on 31 August 2023).
3. Results and Discussion
3.1. Structure and Organization of the T. indicus Mitogenome
Herein, the complete mitogenome of T. indicus (GenBank accession number: OR459825) was successfully sequenced and annotated. It was a circular and double-stranded DNA molecule, consisting of a typical structure with 13 PCGs, 2 rRNAs, 22 tRNAs, and a major non-coding D-loop region (Table 2; Figure 2). Among these 37 genes, 28 were located on the heavy strand, while the remaining nine genes, including eight tRNAs (trnQ, trnA, trnN, trnC, trnY, trnS2, trnE and trnP) and one PCG (ND6), were located on the light strand (Table 2; Figure 2). T. indicus showed the typical avian mitogenome order [21,70], which was also the ancestral avian arrangement found in many lineages of Passeriformes [21]. The mitogenome structure and organization of T. indicus was consistent with those of T. cyanurus, but the T. indicus mitogenome (16,723 bp) was smaller in size than the T. cyanurus mitogenome (16,803 bp), and the interspecific difference was mainly caused by the size difference in the D-loop region located between trnE and trnF (Table 2).
Table 2.
The mitochondrial genome comparison between T. indicus and T. cyanurus.
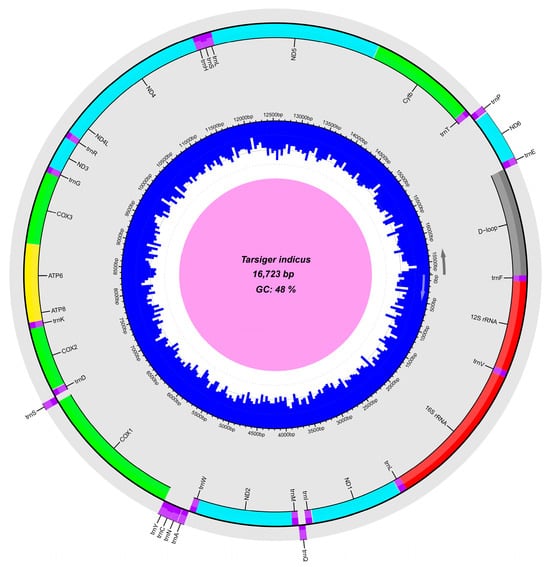
Figure 2.
Graphical representation of Tarsiger indicus mitogenome. Genes outside the outer multicolored circle are located on the light strand counterclockwise, and those inside the outer circle are located on the heavy strand clockwise. Different colors indicate different types of genes and regions. The inner blue circle represents the local GC content.
3.2. Codon Usage
Among the 13 PCGs, the smallest one was ATP8, and the largest one was ND5, ranging from 168 bp to 1818 bp (Table 2). Most PCGs of T. indicus were initiated with the typical start codon ATG, while COX1 and ND2 were started with GTG (Table 2). The unusual start codon GTG was also observed in COX1 from other bird groups, such as Sittidae [71,72], Accipitridae [73,74], Phasianidae [75], Columbidae [76], and other Passeriformes species [24,25,30,36,45]. The stop codons of 13 PCGs were quite varied in T. indicus. ATP6, ATP8, COX2, Cytb, ND3, ND4L, and ND6 were terminated with the representative stop codon TAA or TAG, COX1, ND1, and ND5 ended with AGA or AGG, while COX3, ND2, and ND4 were occasionally terminated with the truncated stop codon TA or T (Table 2). The incomplete stop codons TA and T are common in metazoan mitogenomes [19,20,51,72], and they can be converted to TAA by post-transcriptional modifications during the mRNA maturation process [77]. The start and stop codons of the 13 PCGs were very similar in the mitogenomes of T. indicus and T. cyanurus, and the only difference was the stop codon of the ND6 gene: the former was TAG, while the latter was AGG (Table 2).
The T. indicus mitogenome contained a total of 3797 codons in its protein-coding regions (Table S2). The three most frequently used codons were CUA (Leu1), AUC (Ile), and UUC (Phe), which were used 347, 217, and 181 times, respectively, and the five least-used codons were UGU (Cys), AGU (Ser1), ACG (Thr), CGG (Arg), and AAG (Lys), which were used 6, 6, 6, 4, and 4 times, respectively (Table S2). As in other birds [76,78,79], amino acids with high frequency encoded by PCGs were Leu (664), Thr (327), and Ala (323) (Table S2).
In addition, RSCU is a reference value to evaluate the frequency of codons encoding the same amino acid [80]. When the RSCU ratio was greater than 1, it indicated that the codon occurred many times [80]. Statistics on the RSCU showed that CUA (3.14), CGA (2.34), and GCC (2.18) were relatively high-frequency in T. indicus mitogenome (Figure 3; Table S2). RSCU values of T. cyanurus mitogenome was also summarized and compared with T. indicus, and these two mitogenomes had very similar characteristics of utilization rate of synonymous codon of single amino acids (Figure 3; Table S2).
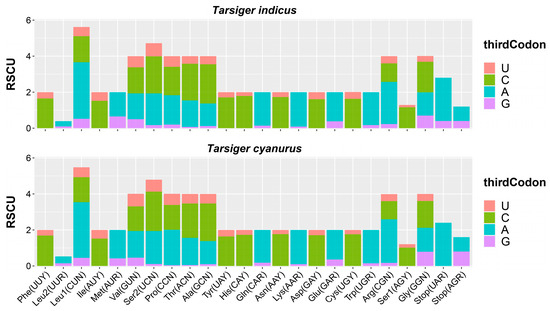
Figure 3.
The relative synonymous codon usage (RSCU) in mitogenomes of T. indicus and T. cyanurus.
3.3. Nucleotide Composition, Diversity, and Evolution
The overall nucleotide composition of the T. indicus mitogenome was 32.88% C, 29.63% A, 22.75% T, and 14.73% G, indicating that the mitogenomes were biased towards C and A bases, which had also been the case in previous studies of avian mitochondrial genomes [18,81]. Its overall G + C content was 47.62%, which was similar to the 47.03% of the T. cyanurus mitogenome (Figure 4). Similar to most other birds [18,37,72], overall G + C content of the whole mitogenomes of all 25 Muscicapidae birds was slightly lower than their overall A + T content (Table S3). In terms of a single mitochondrial gene of Muscicapidae species including T. indicus, the individual G + C contents were very close to 50% (Table S3; Figure 4 and Figure 5). Although T. indicus and T. cyanurus were closely related species, their individual G + C content had an inconsistent trend among all genes (Figure 4).
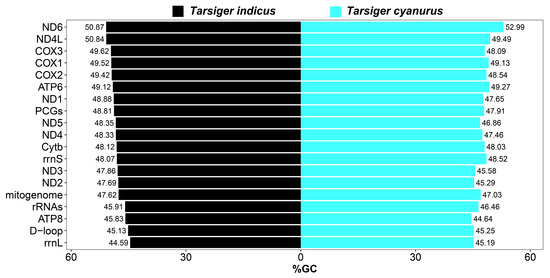
Figure 4.
The G + C content (%GC) of T. indicus and T. cyanurus mitogenomes.
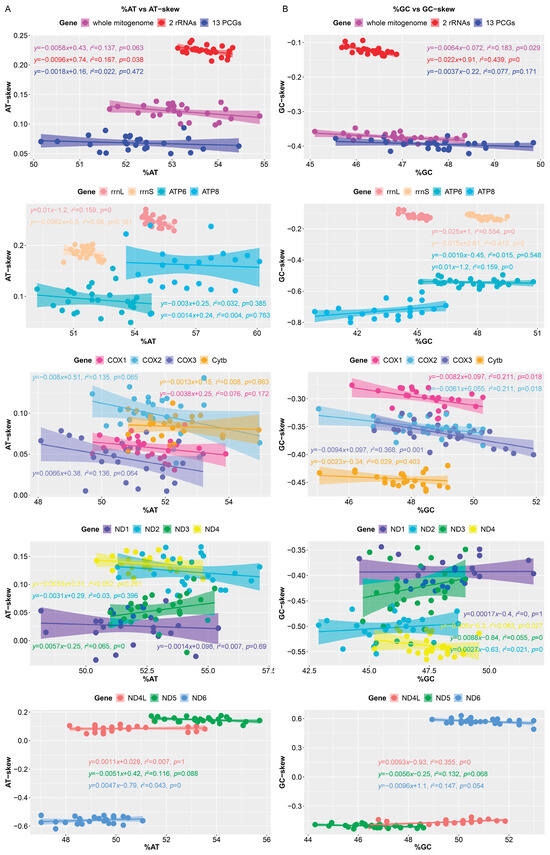
Figure 5.
Correlation between nucleotide content and corresponding skew in the mitogenomes of 26 species of Muscicapidae. (A) A + T content (%AT) vs. AT-skew; (B) G + C content (%GC) vs. GC-skew. Each dot represents a mitogenome.
We also calculated the nucleotide skew of mitochondrial gene in 25 Muscicapidae species. The AT-skew values of the entire genome, concatenated rRNAs, concatenated PCGs, and single rRNA and PCG (except ND6) were positive, while the GC-skew values were negative (Figure 5), as was common in mitogenomes of Strigiformes [18] and Accipitriformes [74], indicating that Cs were more abundant than Gs, and As were more abundant than Ts. AT-skew and GC-skew were due to the different distribution of nucleotides between the two DNA strands, which further led to an asymmetry in the DNA strands [51,80]. We also analyzed the correlation between nucleotide content and corresponding skew of all mitogenomes of Muscicapidae (Figure 5), but the correlation was weak and further confirmation was needed with more data.
The nucleotides varied greatly among different genes (Figure 6). The average nucleotide diversity values for individual genes ranged from 0.04264 (rrnS) to 0.16538 (ND2), and the percentage of nucleotide variable sites ranged from 18.05% (rrnL) to 52.93% (ND2) (Figure 6A), indicating that rrnL and rrnS were slow-evolving genes, ND2 was a fast-evolving gene.
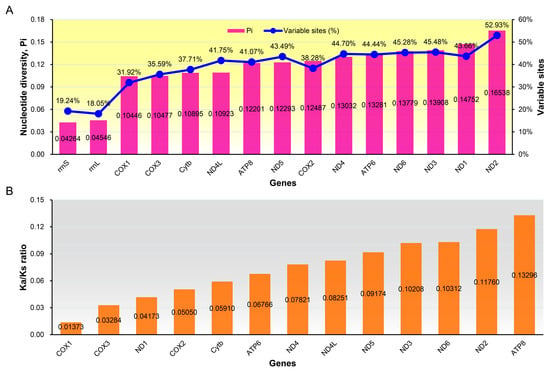
Figure 6.
Evolutionary rates of mitochondrial genes of 25 species of Muscicapidae. (A) Nucleotide diversity and percentage of variable sites; (B)The ratio of non-synonymous substitution rate and synonymous substitution rate.
To further understand the role of selective pressure on the mitochondrial PCGs among the Muscicapidae species, we calculated and compared the average Ka/Ks ratio for each PCG (Figure 6B). Ka/Ks ratio = 1 denotes neutral mutations, Ka/Ks ratio < 1 denotes negative selection, and Ka/Ks ratio > 1 denotes positive selection [82,83]. Here, the average Ka/Ks ratio for all PCGs were consistently far lower than 1, indicating that all PCGs of Muscicapidae mitogenomes had experienced purifying selection. Among the 13 PCGs, ATP8 had the highest rate of evolution (0.13296), whereas COX1 had the lowest (0.01373) (Figure 6B), which was congruent with the previous studies in Passeriformes [51,71], Piciformes [79], Strigiformes [18], and penguins [84], as well as frogs [85]. Therefore, our findings confirmed that COX1 experienced the strongest purifying selection and COX1 might play important roles in the evolution of avian mitogenomes.
3.4. Mitochondrial Phylogenomics
The ML and BI trees of the 13PCGs + 2rRNAs dataset had similar topologies, and most nodes were supported by high bootstrap percentages (BP) and Bayesian posterior probabilities (BPP) (Figure 7 and Figure S1).
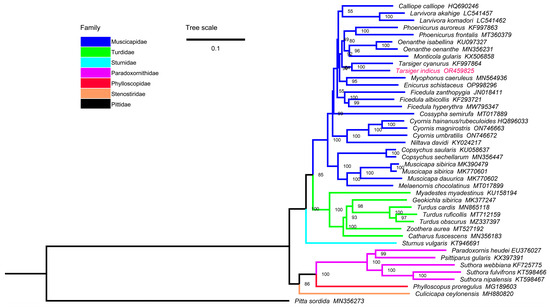
Figure 7.
The phylogenetic relationships of Passeriformes inferred by ML method based on the 13PCGs + 2rRNAs dataset. Numbers on nodes are the bootstrap percentages.
Our results showed that Muscicapidae, Turdidae, and Paradoxornithidae were clustered into two monophyletic groups, and species of the same genus were clustered together with a high degree of confidence. Muscicapidae and Turdidae were sister groups (BP = 85, BPP = 1.00), and they clustered together with Sturnidae (BP = 100, BPP = 1.00), which was consistent with a previous study [38]. T. indicus and T. cyanurus were clustered together with high confidence (BP = 100, BPP = 1.00). These two Tarsiger birds were previously placed in the genus Luscinia [49]. Although many species of Muscicapidae, such as M. gularis, T. indicus, and T. cyanurus were allocated to Turdidae in some older works [50,86] and the up-to-date NCBI taxonomy database; our phylogenetic topologies clearly supported their membership in the Muscicapidae family. It is important to note that the phylogenetic relationships between some genera within Muscicapidae are problematic between our study and a previous study [23]. The position of C. semirufa in our ML and BI trees was not consistent, and different from the ML tree based on a 13 PCGs dataset in a Yang et al. study [23], and the degree of confidence of related branches was not high (Figure 7 and Figure S1). Our ML and BI trees showed consistent topology (Calliope + Larvivora) + Ficedula (Figure 7 and Figure S1); however, the ML tree of the Yang et al. study showed the diametrical topology Calliope + (Ficedula + Larvivora) with low bootstrap percentages [23]. Complete mitogenomes may provide more accurate signals than gene fragments for phylogenetic reconstruction. Overall, the current 25 species represent only 7% of the old-world flycatchers group, so, in order to better resolve the phylogenetic relationships within Muscicapidae, it is still necessary to obtain more mitochondrial genome sequences of old-world flycatchers.
In addition, P. heudei, S. webbiana, S. nipalensis, and S. fulvifrons were classified into Muscicapidae in previous studies [57,71] and the NCBI taxonomy database, but our results showed that these species clustered into the Paradoxornithidae family [87]. The taxonomic history of C. ceylonensis was also complex [72]. C. ceylonensis was originally classified into the Muscicapidae family based on external morphology, reproductive habits, and nesting characteristics [86]. Subsequently, it was classified into the family Rhipiduridae [88]. Lately, the phylogenetic analyses based on multilocus sequence data revealed that C. ceylonensis was in fact a member of the Stenostiridae family [62]. Here, we also clarified its taxonomic validity based on mitochondrial genome approach.
4. Conclusions
In this study, we successfully sequenced the mitogenome of T. indicus using the Illumina Novaseq 6000 platform with a paired-end read length of 150 bp. We also annotated and summarized its mitogenomic characteristics in detail. Importantly, we conducted the first comprehensive mitogenome analysis of Muscicapidae. The mitogenome of T. indicus mitogenome contained the typical avian mitochondrial gene arrangement. T. cyanurus and T. indicus shared very similar mitogenomic features. All 13 PCGs of the mitogenomes of Muscicapidae had experienced purifying selection. The monophylies of Muscicapidae, Turdidae, and Paradoxornithidae were strongly supported. The clade of ((Muscicapidae + Turdidae) + Sturnidae) in Passeriformes was supported by both BI and ML analyses. The current taxonomic status of many passerine birds with complex taxonomic histories were also supported. Our study provides the first complete mitochondrial genome of T. indicus to enrich its genetic data. A large number of studies on the mitochondrial genome of Muscicapidae are still needed in the future to further solve some phylogenetic problems.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/genes15010090/s1, Table S1: The partition and best-fit partition models used in this study; Table S2: The codon usage in the mitogenomes of T. indicus and T. cyanurus; Table S3: The nucleotide composition and skew in the mitogenomes of 25 species of Muscicapidae; Figure S1: The phylogenetic relationships of Passeriformes inferred by BI method based on the 13PCGs + 2rRNAs dataset. Numbers on nodes are the Bayesian posterior probabilities.
Author Contributions
Conceptualization, J.L. (Jiabin Liu) and G.Q.; Data curation, J.Y.; Formal analysis, J.Y.; Funding acquisition, G.L. and J.L. (Jiabin Liu); Investigation, G.L., J.L. (Juan Liu) and Y.Z. (Yue Zhang); Methodology, G.L. and J.Y.; Project administration, J.L. (Jiabin Liu) and G.Q.; Resources, G.L.; Visualization, J.Y.; Writing—original draft, G.L. and J.Y.; Writing—review & editing, J.L. (Juan Liu), Y.Z. (Yue Zhang), R.M., Y.Z. (Yanshan Zhou), B.Z., W.W., J.L. (Jiabin Liu) and G.Q. All authors have read and agreed to the published version of the manuscript.
Funding
This work was financially supported by the National Natural Science Foundation of China (Grant No. U21A20193), the Sichuan Science and Technology Program (Grant No. 2023NSFSC1156), and the Capacity Building Project of Baihe National Nature Reserve in 2022 (Grant No. N5132252022000101-3).
Institutional Review Board Statement
Ethical review and approval were not required because the specimen used in this study was a subadult bird that crashed into a window and died.
Informed Consent Statement
Not applicable.
Data Availability Statement
The GenBank accession number of the newly determined Tarsiger indicus mitogenome sequence is OR459825. The BioProject, BioSample, and SRA accession numbers of metadata are PRJNA1006441, SAMN37041239, and SRR25670941, respectively.
Acknowledgments
We gratefully acknowledge Taifu Hu, a bird lover from Longcanggou Town of Yingjing County, for providing the photo of T. indicus. We truly thank HOME for Researchers (http://www.home-for-researchers.com/, accessed on 2 October 2023) for the linguistic assistance during the preparation of this manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Gill, F.; Donsker, D.; Rasmussen, P. IOC World Bird List (v13.2). Available online: https://www.worldbirdnames.org/new/ioc-lists/master-list-2/ (accessed on 8 August 2023).
- BirdLife International. Tarsiger indicus. The IUCN Red List of Threatened Species 2016: E.T22709743A94221641. Available online: https://www.iucnredlist.org/species/22709743/94221641 (accessed on 12 August 2023).
- Zheng, G. A Checklist on the Classification and Distribution of the Birds of China, 3rd ed.; Science Press: Beijing, China, 2017; p. 492. [Google Scholar]
- Liu, S.; Yu, J.; Cui, J.; Zhou, Y. White-browed Bush Robin (Tarsiger indicus) found in Shennongjia, Hubei Province. Hubei For. Sci. Technol. 2017, 46, 45–75. [Google Scholar]
- Zeng, Z.; Gong, H. White-browed Bush Robin (Tarsiger indicus) found in the southern slope of Qinling Mountains in Shaanxi Province. Chin. J. Zool. 2018, 53, 39. [Google Scholar]
- Wei, C.; Sangster, G.; Olsson, U.; Rasmussen, P.C.; Svensson, L.; Yao, C.-T.; Carey, G.J.; Leader, P.J.; Zhang, R.; Chen, G.; et al. Cryptic species in a colorful genus: Integrative taxonomy of the bush robins (Aves, Muscicapidae, Tarsiger) suggests two overlooked species. Mol. Phylogenet. Evol. 2022, 175, 107580. [Google Scholar] [CrossRef] [PubMed]
- Zheng, G. A Checklist on the Classification and Distribution of the Birds of China, 4th ed.; Science Press: Beijing, China, 2023; p. 498. [Google Scholar]
- National Forestry and Grassland Administration. List of Terrestrial Wild Animals of Important Ecological, Scientific and Social Value in China. Available online: https://www.forestry.gov.cn/c/www/gsgg/509640.jhtml (accessed on 8 August 2023).
- Jiang, Z.; Jiang, J.; Wang, Y.; Zhang, E.; Zhang, Y.; Li, L.; Xie, F.; Cai, B.; Cao, L.; Zheng, G.; et al. Red List of China’s Vertebrates. Biodivers. Sci. 2016, 24, 500–551. [Google Scholar]
- Montaña-Lozano, P.; Moreno-Carmona, M.; Ochoa-Capera, M.; Medina, N.S.; Boore, J.L.; Prada, C.F. Comparative genomic analysis of vertebrate mitochondrial reveals a differential of rearrangements rate between taxonomic class. Sci. Rep. 2022, 12, 5479. [Google Scholar] [CrossRef] [PubMed]
- Lavrov, D.V.; Pett, W. Animal mitochondrial DNA as we do not know it: Mt-genome organization and evolution in nonbilaterian lineages. Genome Biol. Evol. 2016, 8, 2896–2913. [Google Scholar] [CrossRef] [PubMed]
- Winkelmann, I.; Campos, P.F.; Strugnell, J.; Cherel, Y.; Smith, P.J.; Kubodera, T.; Allcock, L.; Kampmann, M.-L.; Schroeder, H.; Guerra, A.; et al. Mitochondrial genome diversity and population structure of the giant squid Architeuthis: Genetics sheds new light on one of the most enigmatic marine species. Proc. R. Soc. B Biol. Sci. 2013, 280, 20130273. [Google Scholar] [CrossRef] [PubMed]
- Deng, M.-X.; Xiao, B.; Yuan, J.-X.; Hu, J.-M.; Kim, K.S.; Westbury, M.V.; Lai, X.-L.; Sheng, G.-L. Ancient mitogenomes suggest stable mitochondrial clades of the Siberian roe deer. Genes 2022, 13, 114. [Google Scholar] [CrossRef]
- Wu, L.; Tong, Y.; Ayivi, S.P.; Storey, K.B.; Zhang, J.-Y.; Yu, D.-N. The complete mitochondrial genomes of three Sphenomorphinae species (Squamata: Scincidae) and the selective pressure analysis on mitochondrial genomes of limbless Isopachys Gyldenstolpei. Animals 2022, 12, 2015. [Google Scholar] [CrossRef]
- Sangster, G.; Luksenburg, J.A. Sharp increase of problematic mitogenomes of birds: Causes, consequences, and remedies. Genome Biol. Evol. 2021, 13, evab210. [Google Scholar] [CrossRef]
- Li, M.; Chen, W.-T.; Zhang, Q.-L.; Liu, M.; Xing, C.-W.; Cao, Y.; Luo, F.-Z.; Yuan, M.-L. Mitochondrial phylogenomics provides insights into the phylogeny and evolution of spiders (Arthropoda: Araneae). Zool. Res. 2022, 43, 566–584. [Google Scholar] [CrossRef] [PubMed]
- Iacolina, L.; Buzan, E.; Safner, T.; Bašić, N.; Geric, U.; Tesija, T.; Lazar, P.; Arnal, M.C.; Chen, J.; Han, J.; et al. A mother’s story, mitogenome relationships in the genus Rupicapra. Animals 2021, 11, 1065. [Google Scholar] [CrossRef]
- Yu, J.; Liu, J.; Li, C.; Wu, W.; Feng, F.; Wang, Q.; Ying, X.; Qi, D.; Qi, G. Characterization of the complete mitochondrial genome of Otus lettia: Exploring the mitochondrial evolution and phylogeny of owls (Strigiformes). Mitochondrial DNA Part B 2021, 6, 3443–3451. [Google Scholar] [CrossRef] [PubMed]
- Catanese, G.; Morey, G.; Verger, F.; Grau, A.M. The Nursehound Scyliorhinus stellaris mitochondrial genome—Phylogeny, relationships among Scyliorhinidae and variability in waters of the Balearic Islands. Int. J. Mol. Sci. 2022, 23, 10355. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Yu, J.; Zhou, M.; Yang, J. Complete mitochondrial genome of Japalura flaviceps: Deep insights into the phylogeny and gene rearrangements of Agamidae species. Int. J. Biol. Macromol. 2019, 125, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Caparroz, R.; Rocha, A.V.; Cabanne, G.S.; Tubaro, P.; Aleixo, A.; Lemmon, E.M.; Lemmon, A.R. Mitogenomes of two neotropical bird species and the multiple independent origin of mitochondrial gene orders in Passeriformes. Mol. Biol. Rep. 2018, 45, 279–285. [Google Scholar] [CrossRef] [PubMed]
- He, K.; Chen, X.; Qiu, Y.-B.; Liu, Z.; Wang, W.-Z.; Woodman, N.; Maldonado, J.E.; Pan, X. Mitogenome and phylogenetic analyses support rapid diversification among species groups of small-eared shrews genus Cryptotis (Mammalia: Eulipotyphla: Soricidae). Zool. Res. 2021, 42, 739–745. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.; Liu, W.; Yang, K. The complete mitochondrial genome of the snowy-browed flycatcher Ficedula hyperythra (Passeriformes, Muscicapidae). Mitochondrial DNA Part B 2021, 6, 2870–2871. [Google Scholar] [CrossRef]
- Li, F.-j.; Qiao, L.; Yang, Y.-j.; Yang, K.; Yang, N.; Yue, B.-s. The complete mitochondrial genome of Phoenicurus frontalis (Passeriformes: Muscicapidae). Mitochondrial DNA Part B 2020, 5, 2322–2323. [Google Scholar] [CrossRef]
- Du, C.; Liu, L.; Liu, Y.; Fu, Z.; Xu, Y. The complete mitochondrial genome of Daurian redstart Phoenicurus auroreus (Aves: Passeriformes: Muscicapidae). Mitochondrial DNA Part B 2020, 5, 2231–2232. [Google Scholar] [CrossRef]
- Zhang, N.; Zhao, Y.; Meng, D.; Zhou, X.; Liu, Z.; Teng, L. The complete mitochondrial genome of the Siberian Rubythroat (Calliope calliope) from Maorshan, China. Mitochondrial DNA Part B 2023, 8, 110–112. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.-H.; Sun, C.-H.; Hou, S.-L.; Huang, Y.-L.; Lu, C.-H. The complete mitochondrial genome of dark-sided flycatcher Muscicapa sibirica (Passeriformes: Muscicapidae). Mitochondrial DNA Part B 2019, 4, 2675–2676. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Sun, C.-H.; Wang, L.-B.; Xue, D.-D.; Xu, P.; Xie, S.-B.; Lu, C.-H. The complete mitochondrial genome of Grey-streaked Flycatcher Muscicapa griseisticta (Passeriformes: Muscicapidae). Mitochondrial DNA Part B 2019, 4, 1857–1858. [Google Scholar] [CrossRef]
- Min, X.; Lu, C.-H.; Chen, T.-Y.; Liu, B.; Lu, C.-H. The complete mitochondrial genome of Asian brown flycatcher Muscicapa latirostris (Passeriformes: Muscicapidae). Mitochondrial DNA Part B 2019, 4, 3880–3881. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Luo, A.; Li, G.; Li, W. Complete mitochondrial genome of the isabelline wheatear Oenanthe isabellina (Passeriformes, Muscicapidae). Mitochondrial DNA Part B 2016, 1, 355–356. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Stiller, J.; Deng, Y.; Armstrong, J.; Fang, Q.; Reeve, A.H.; Xie, D.; Chen, G.; Guo, C.; Faircloth, B.C.; et al. Dense sampling of bird diversity increases power of comparative genomics. Nature 2020, 587, 252–257. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.-F.; Yang, D.-C.; Lu, C.-H. Complete mitochondrial genome of oriental magpie-robin Copsychus saularis (Aves: Muscicapidae). Mitochondrial DNA Part B 2016, 1, 21–22. [Google Scholar] [CrossRef] [PubMed]
- Ekblom, R.; Smeds, L.; Ellegren, H. Patterns of sequencing coverage bias revealed by ultra-deep sequencing of vertebrate mitochondria. BMC Genom. 2014, 15, 467. [Google Scholar] [CrossRef]
- Zhang, H.; Bai, Y.; Shi, X.; Sun, L.; Wang, Z.; Wu, X. The complete mitochondrial genomes of Tarsiger cyanurus and Phoenicurus auroreus: A phylogenetic analysis of Passeriformes. Genes Genom. 2018, 40, 151–165. [Google Scholar] [CrossRef]
- Zhou, C.; Jin, J.; Chen, Y.; Hao, Y.; Zhang, X.; Meng, Y.; Yang, N.; Yue, B. Two new complete mitochondrial genomes (Paradoxornis gularis and Niltava davidi) and their phylogenetic and taxonomic implications. Mitochondrial DNA Part B 2019, 4, 820–821. [Google Scholar] [CrossRef]
- Zhang, H.; Cheng, Y.; Zhou, L.; Dong, Y. Complete mitochondrial genome of white-throated rock-thrush Monticola cinclorhynchus gularis (Passeriformes: Muscicapidae). Mitochondrial DNA Part B 2016, 1, 684–685. [Google Scholar] [CrossRef]
- Zhang, X.; Ma, X.; Ha, J.; Wei, Y.; Zhao, Y.; Wei, S.; Zhong, X.; Cai, Z. Characterization of a complete mitochondrial genome of Turdus ruficollis (Passeriformes: Turdidae). Mitochondrial DNA Part B 2021, 6, 1623–1624. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Dong, Y.; Sun, G.; Wang, Q.; Dong, Y.; Dou, H.; Wei, Q. The complete mitochondrial genome of Turdus obscurus (Passeriformes: Turdidae). Mitochondrial DNA Part B 2021, 6, 3076–3077. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.-H.; Xu, P.; Lu, C.-H.; Han, Q.; Lin, Y.-F.; Gao, Z.-J.; Lu, C.-H. Complete mitochondrial genome of the Grey thrush Turdus cardis (Aves, Turdidae). Mitochondrial DNA Part B 2020, 5, 1824–1825. [Google Scholar] [CrossRef]
- Li, J.; Miao, K.; Wu, X.; Wang, Q.; Hu, C.; Li, P. The complete mitogenome of scaly thrush Zoothera aurea (Passeriformes, Turdidae). Mitochondrial DNA Part B 2020, 5, 2936–2937. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.-H.; Liu, B.; Lu, C.-H. Complete mitochondrial genome of the Siberian thrush, Geokichla sibirica sibirica (Aves, Turdidae). Mitochondrial DNA Part B 2019, 4, 1150–1151. [Google Scholar] [CrossRef]
- Anmarkrud, J.A.; Lifjeld, J.T. Complete mitochondrial genomes of eleven extinct or possibly extinct bird species. Mol. Ecol. Resour. 2017, 17, 334–341. [Google Scholar] [CrossRef] [PubMed]
- Rollins, L.A.; Woolnough, A.P.; Fanson, B.G.; Cummins, M.L.; Crowley, T.M.; Wilton, A.N.; Sinclair, R.; Butler, A.; Sherwin, W.B. Selection on mitochondrial variants occurs between and within individuals in an expanding invasion. Mol. Biol. Evol. 2016, 33, 995–1007. [Google Scholar] [CrossRef]
- Wen, L.; Yang, X.; Liao, J.; Fu, Y.; Dai, B. The complete mitochondrial genome of the fulvous parrotbill Paradoxornis fulvifrons (Passeriformes: Muscicapidae). Mitochondrial DNA Part A 2017, 28, 143–144. [Google Scholar] [CrossRef]
- Zhang, H.; Li, Y.; Wu, X.; Xue, H.; Yan, P.; Wu, X.-B. The complete mitochondrial genome of Paradoxornis webbianus (Passeriformes, Muscicapidae). Mitochondrial DNA 2015, 26, 879–880. [Google Scholar] [CrossRef]
- Jiao, S.-Y.; Liu, Z.-X.; Yu, F.; Yao, J.-Y.; Li, Y.-M.; Yan, S.-Q. Complete mitochondrial genome of Pallas’s Leaf Warbler (Phylloscopus proregulus). Mitochondrial DNA Part B 2018, 3, 211–212. [Google Scholar] [CrossRef] [PubMed]
- Tu, H.; Gao, J.; Jing, J.; Zhou, C.; Yue, B.; Zhang, X. The complete mitochondrial genome of grey-headed canary-flycatcher (Culicicapa ceylonensis, Muscicapidae, Passeriformes). Mitochondrial DNA Part B 2019, 4, 1079–1080. [Google Scholar] [CrossRef]
- Luo, T.; Yan, S.-S.; Xiao, N.; Zhou, J.-J.; Wang, X.-L.; Chen, W.-C.; Deng, H.-Q.; Zhang, B.-W.; Zhou, J. Phylogenetic analysis of combined mitochondrial genome and 32 nuclear genes provides key insights into molecular systematics and historical biogeography of Asian warty newts of the genus Paramesotriton (Caudata: Salamandridae). Zool. Res. 2022, 43, 787–804. [Google Scholar] [CrossRef] [PubMed]
- Sangster, G.; Alström, P.; Forsmark, E.; Olsson, U. Multi-locus phylogenetic analysis of Old World chats and flycatchers reveals extensive paraphyly at family, subfamily and genus level (Aves: Muscicapidae). Mol. Phylogenetics Evol. 2010, 57, 380–392. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X. A Photographic Guide to the Birds of China; The Commercial Press: Beijing, China, 2018; p. 980. [Google Scholar]
- Liu, J.; Yu, J.; Yu, X.; Bi, W.; Yang, H.; Xue, F.; Zhang, G.; Zhang, J.; Yi, D.; Ma, R.; et al. Complete mitogenomes of ticks Ixodes acutitarsus and Ixodes ovatus parasitizing giant panda: Deep insights into the comparative mitogenomic and phylogenetic relationship of Ixodidae species. Genes 2022, 13, 2049. [Google Scholar] [CrossRef] [PubMed]
- Donath, A.; Jühling, F.; Al-Arab, M.; Bernhart, S.H.; Reinhardt, F.; Stadler, P.F.; Middendorf, M.; Bernt, M. Improved annotation of protein-coding genes boundaries in metazoan mitochondrial genomes. Nucleic Acids Res. 2019, 47, 10543–10552. [Google Scholar] [CrossRef]
- Zhu, T.; Sato, Y.; Sado, T.; Miya, M.; Iwasaki, W. MitoFish, MitoAnnotator, and MiFish Pipeline: Updates in 10 Years. Mol. Biol. Evol. 2023, 40, msad035. [Google Scholar] [CrossRef] [PubMed]
- Canbäck, B.; Laslett, D. ARWEN: A program to detect tRNA genes in metazoan mitochondrial nucleotide sequences. Bioinformatics 2008, 24, 172–175. [Google Scholar]
- Zheng, S.; Poczai, P.; Hyvönen, J.; Tang, J.; Amiryousefi, A. Chloroplot: An online program for the versatile plotting of organelle genomes. Front. Genet. 2020, 11, 576124. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef] [PubMed]
- Lyu, F.; Han, F.; Ge, C.; Mao, W.; Chen, L.; Hu, H.; Chen, G.; Lang, Q.; Fang, C. OmicStudio: A composable bioinformatics cloud platform with real-time feedback that can generate high-quality graphs for publication. iMeta 2023, 2, e85. [Google Scholar] [CrossRef]
- Jarvis, E.D.; Mirarab, S.; Aberer, A.J.; Li, B.; Houde, P.; Li, C.; Ho, S.Y.W.; Faircloth, B.C.; Nabholz, B.; Howard, J.T.; et al. Whole-genome analyses resolve early branches in the tree of life of modern birds. Science 2014, 346, 1320–1331. [Google Scholar] [CrossRef] [PubMed]
- Kimball, R.T.; Oliveros, C.H.; Wang, N.; White, N.D.; Barker, F.K.; Field, D.J.; Ksepka, D.T.; Chesser, R.T.; Moyle, R.G.; Braun, M.J.; et al. A phylogenomic supertree of birds. Diversity 2019, 11, 109. [Google Scholar] [CrossRef]
- Fuchs, J.; Pasquet, E.; Couloux, A.; Fjeldså, J.; Bowie, R.C.K. A new Indo-Malayan member of the Stenostiridae (Aves: Passeriformes) revealed by multilocus sequence data: Biogeographical implications for a morphologically diverse clade of flycatchers. Mol. Phylogenetics Evol. 2009, 53, 384–393. [Google Scholar] [CrossRef] [PubMed]
- Ranwez, V.; Douzery, E.J.P.; Cambon, C.; Chantret, N.; Delsuc, F. MACSE v2: Toolkit for the alignment of coding sequences accounting for frameshifts and stop codons. Mol. Biol. Evol. 2018, 35, 2582–2584. [Google Scholar] [CrossRef] [PubMed]
- Talavera, G.; Castresana, J. Improvement of phylogenies after removing divergent and ambiguously aligned blocks from protein sequence alignments. Syst. Biol. 2007, 56, 564–577. [Google Scholar] [CrossRef]
- Xiang, C.-Y.; Gao, F.; Jakovlić, I.; Lei, H.-P.; Hu, Y.; Zhang, H.; Zou, H.; Wang, G.-T.; Zhang, D. Using PhyloSuite for molecular phylogeny and tree-based analyses. iMeta 2023, 2, e87. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Nguyen, M.A.T.; von Haeseler, A. Ultrafast approximation for phylogenetic bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef] [PubMed]
- Urantówka, A.D.; Kroczak, A.; Mackiewicz, P. New view on the organization and evolution of Palaeognathae mitogenomes poses the question on the ancestral gene rearrangement in Aves. BMC Genom. 2020, 21, 874. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.; Bi, D.; Han, S.; Yi, R.; Zhang, S.; Ye, Y.; Gao, J.; Yang, J.; Kan, X. Mitogenomic codon usage patterns of superfamily Certhioidea (Aves, Passeriformes): Insights into asymmetrical bias and phylogenetic implications. Animals 2023, 13, 96. [Google Scholar] [CrossRef]
- Yuan, Q.; Guo, Q.; Cao, J.; Luo, X.; Duan, Y. Description of the three complete mitochondrial genomes of Sitta (S. himalayensis, S. nagaensis, and S. yunnanensis) and phylogenetic relationship (Aves: Sittidae). Genes 2023, 14, 589. [Google Scholar] [CrossRef]
- Adawaren, E.O.; Du Plessis, M.; Suleman, E.; Kindler, D.; Oosthuizen, A.O.; Mukandiwa, L.; Naidoo, V. The complete mitochondrial genome of Gyps coprotheres (Aves, Accipitridae, Accipitriformes): Phylogenetic analysis of mitogenome among raptors. PeerJ 2020, 8, e10034. [Google Scholar] [CrossRef]
- Jiang, L.; Peng, L.; Tang, M.; You, Z.; Zhang, M.; West, A.; Ruan, Q.; Chen, W.; Merilä, J. Complete mitochondrial genome sequence of the Himalayan Griffon, Gyps himalayensis (Accipitriformes: Accipitridae): Sequence, structure, and phylogenetic analyses. Ecol. Evol. 2019, 9, 8813–8828. [Google Scholar] [CrossRef]
- Li, X.; Huang, Y.; Lei, F. Comparative mitochondrial genomics and phylogenetic relationships of the Crossoptilon species (Phasianidae, Galliformes). BMC Genom. 2015, 16, 42. [Google Scholar] [CrossRef]
- Liu, H.-Y.; Sun, C.-H.; Zhu, Y.; Zhang, Q.-Z. Complete mitogenomic and phylogenetic characteristics of the speckled wood-pigeon (Columba hodgsonii). Mol. Biol. Rep. 2020, 47, 3567–3576. [Google Scholar] [CrossRef]
- Ojala, D.; Montoya, J.; Attardi, G. tRNA punctuation model of RNA processing in human mitochondria. Nature 1981, 290, 470–474. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.-H.; Liu, H.-Y.; Min, X.; Lu, C.-H. Mitogenome of the little owl Athene noctua and phylogenetic analysis of Strigidae. Int. J. Biol. Macromol. 2020, 151, 924–931. [Google Scholar] [CrossRef] [PubMed]
- Bi, D.; Ding, H.; Wang, Q.; Jiang, L.; Lu, W.; Wu, X.; Zhu, R.; Zeng, J.; Zhou, S.; Yang, X.; et al. Two new mitogenomes of Picidae (Aves, Piciformes): Sequence, structure and phylogenetic analyses. Int. J. Biol. Macromol. 2019, 133, 683–692. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zheng, Y.; Zhao, X.; Xu, X.; Xu, Z.; Cui, C. Molecular phylogeny and evolution of the Tuerkayana (Decapoda: Brachyura: Gecarcinidae) genus based on whole mitochondrial genome sequences. Biology 2023, 12, 974. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Zhou, L.; Li, B.; Zhang, L. The complete mitochondrial genome of Aix galericulata and Tadorna ferruginea: Bearings on their phylogenetic position in the Anseriformes. PLoS ONE 2014, 9, e109701. [Google Scholar] [CrossRef] [PubMed]
- Jia, C.; Zhang, X.; Xu, S.; Yang, T.; Yanagimoto, T.; Gao, T. Comparative analysis of the complete mitochondrial genomes of three rockfishes (Scorpaeniformes, Sebastiscus) and insights into the phylogenetic relationships of Sebastidae. Biosci. Rep. 2020, 40, BSR20203379. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.-H.; Huang, H.-M.; Wu, L.; Storey, K.B.; Zhang, J.-Y.; Zhang, Y.-P.; Yu, D.-N. Characterization of two mitogenomes of Hyla sanchiangensis (Anura: Hylidae), with phylogenetic relationships and selection pressure analyses of Hylidae. Animals 2023, 13, 1593. [Google Scholar] [CrossRef] [PubMed]
- Ramos, B.; González-Acuña, D.; Loyola, D.E.; Johnson, W.E.; Parker, P.G.; Massaro, M.; Dantas, G.P.M.; Miranda, M.D.; Vianna, J.A. Landscape genomics: Natural selection drives the evolution of mitogenome in penguins. BMC Genom. 2018, 19, 53. [Google Scholar] [CrossRef]
- Chen, W.; Qian, W.; Miao, K.; Qian, R.; Yuan, S.; Liu, W.; Dai, J.; Hu, C.; Chang, Q. Comparative mitogenomics of true frogs (Ranidae, Anura), and its implications for the phylogeny and evolutionary history of Rana. Animals 2022, 12, 1250. [Google Scholar] [CrossRef]
- Zheng, G. A Checklist on the Classification and Distribution of the Birds of the World; Science Press: Beijing, China, 2002; p. 400. [Google Scholar]
- Cai, T.; Cibois, A.; Alström, P.; Moyle, R.G.; Kennedy, J.D.; Shao, S.; Zhang, R.; Irestedt, M.; Ericson, P.G.P.; Gelang, M.; et al. Near-complete phylogeny and taxonomic revision of the world’s babblers (Aves: Passeriformes). Mol. Phylogenet. Evol. 2019, 130, 346–356. [Google Scholar] [CrossRef]
- Zheng, G. A Checklist on the Classification and Distribution of the Birds of China, 1st ed.; Science Press: Beijing, China, 2005; p. 444. [Google Scholar]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).