
The Mitochondrial Genomes of Siboglinum plumosum and Oligobrachia dogieli (Annelida: Siboglinidae) and Their Phylogenetic Analysis
 , , , and
, , , and
Abstract

1. Introduction
2. Materials and Methods
2.1. Sample Collection and DNA Extraction and Sequencing
2.2. Assembly and Annotation
2.3. Phylogenetic Analysis
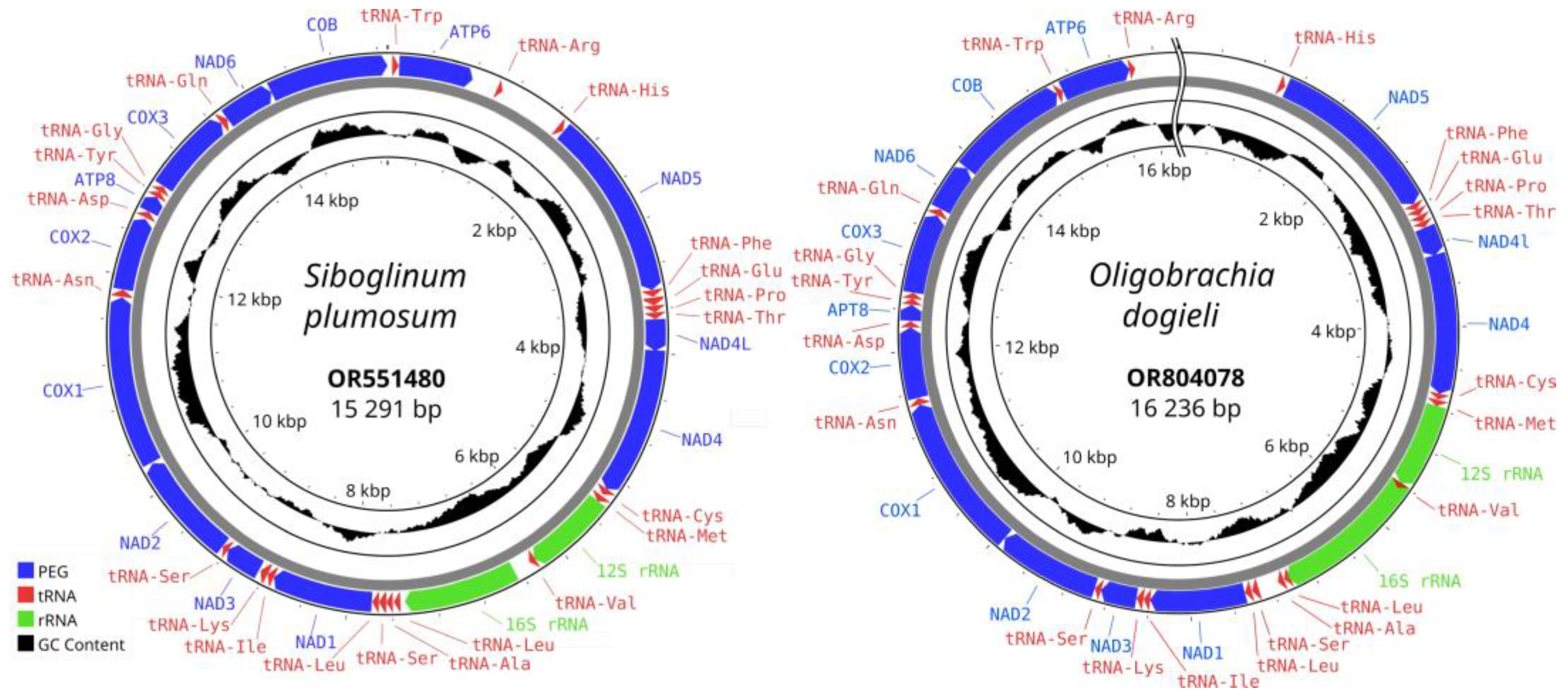
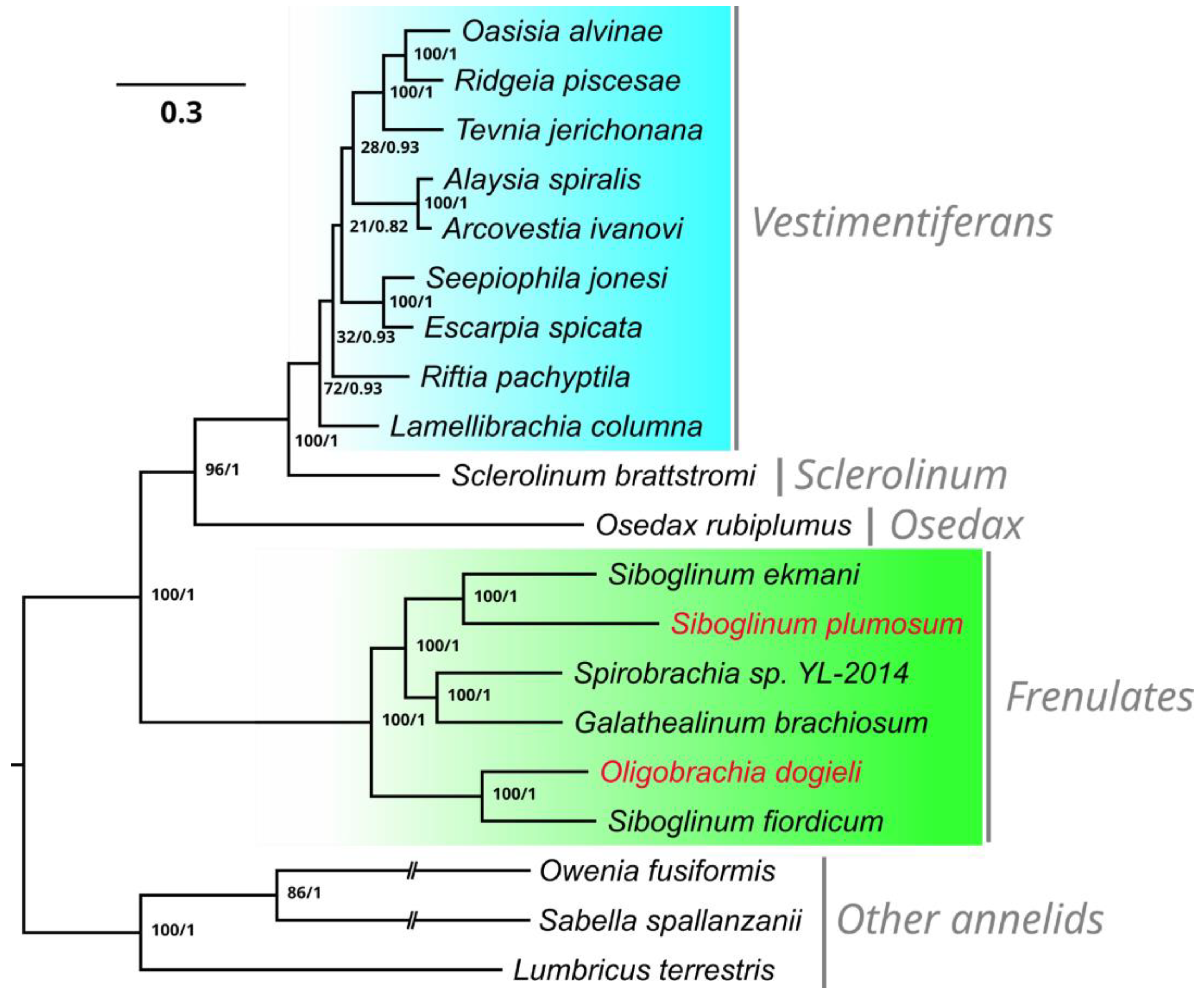
3. Results
4. Discussion
4.1. Structure of the Assembled Mitochondrial Genomes
4.2. The Phylogenetic Relationships of Siboglinidae
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rousset, V.; Rouse, G.W.; Siddall, M.E.; Tillier, A.; Pleijel, F. The Phylogenetic Position of Siboglinidae (Annelida) Inferred from 18S RRNA, 28S RRNA and Morphological Data. Cladistics 2004, 20, 518–533. [Google Scholar] [CrossRef] [PubMed]
- Struck, T.H. Phylogeny. In Handbook of Zoology. Annelida. Volume 1 Annelida Basal Groups and Pleistoannelida, Sedentaria I; Purschke, G., Böggemann, M., Westheide, W., Eds.; De Gruyter: Berlin/Heidelberg, Germany; Boston, MA, USA, 2019; pp. 37–68. ISBN 9783110291582. [Google Scholar]
- Sen, A.; Duperron, S.; Hourdez, S.; Piquet, B.; Léger, N.; Gebruk, A.; Le Port, A.S.; Svenning, M.M.; Andersen, A.C. Cryptic Frenulates Are the Dominant Chemosymbiotrophic Fauna at Arctic and High Latitude Atlantic Cold Seeps. PLoS ONE 2018, 13, e0209273. [Google Scholar] [CrossRef] [PubMed]
- Karaseva, N.; Gantsevich, M.; Obzhirov, A.; Shakirov, R.; Starovoitov, A.; Smirnov, R.; Malakhov, V. Correlation of the Siboglinid (Annelida: Siboglinidae) Distribution to Higher Concentrations of Hydrocarbons in the Sea of Okhotsk. Mar. Pollut. Bull. 2020, 158, 111448. [Google Scholar] [CrossRef] [PubMed]
- Smirnov, R.V. On a New Classification of the Genus Siboglinum Caullery, 1914 (Annelida: Pogonophora). Proc. Zool. Inst. RAS 2014, 318, 48–69. [Google Scholar] [CrossRef]
- Halanych, K.M.; Feldman, R.A.; Vrijenhoek, R.C. Molecular Evidence That Sclerolinum Brattstromi Is Closely Related to Vestimentiferans, Not to Frenulate Pogonophorans (Siboglinidae, Annelida). Biol. Bull. 2001, 201, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Smirnov, R.V. A Revision of the Oligobrachiidae (Annelida: Pogonophora), with Notes on the Morphology and Distribution of Oligobrachia Haakonmosbiensis Smirnov. Mar. Biol. Res. 2014, 10, 972–982. [Google Scholar] [CrossRef]
- Halanych, K.M. Molecular Phylogeny of Siboglinid Annelids (a.k.a. Pogonophorans): A Review. Hydrobiologia 2005, 535–536, 297–307. [Google Scholar] [CrossRef]
- Eichinger, I.; Hourdez, S.; Bright, M. Morphology, Microanatomy and Sequence Data of Sclerolinum Contortum (Siboglindae, Annelida) of the Gulf of Mexico. Org. Divers. Evol. 2013, 13, 311–329. [Google Scholar] [CrossRef]
- Andrade, S.C.S.; Novo, M.; Kawauchi, G.Y.; Worsaae, K.; Pleijel, F.; Giribet, G.; Rouse, G.W. Articulating “Archiannelids”: Phylogenomics and Annelid Relationships, with Emphasis on Meiofaunal Taxa. Mol. Biol. Evol. 2015, 32, 2860–2875. [Google Scholar] [CrossRef]
- Ivanov, A.V. Pogonophora; Academic Press: London, UK, 1963. [Google Scholar]
- Webb, M. Evolutionary Paths within the Phylum Pogonophora. Sarsia 1964, 16, 59–64. [Google Scholar] [CrossRef]
- Southward, E.C. Recent Researches on the Pogonophora. Oceanogr. Mar. Biol. An Annu. Rev. 1971, 9, 193–220. [Google Scholar]
- Rouse, G.W. A Cladistic Analysis of Siboglinidae Caullery, 1914 (Polychaeta, Annelida): Formerly the Phyla Pogonophora and Vestimentifera. Zool. J. Linn. Soc. 2001, 132, 55–80. [Google Scholar] [CrossRef]
- Li, Y.; Kocot, K.M.; Schander, C.; Santos, S.R.; Thornhill, D.J.; Halanych, K.M. Mitogenomics Reveals Phylogeny and Repeated Motifs in Control Regions of the Deep-Sea Family Siboglinidae (Annelida). Mol. Phylogenet. Evol. 2015, 85, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.J.; Yu, W.B.; Yang, J.B.; Song, Y.; Depamphilis, C.W.; Yi, T.S.; Li, D.Z. GetOrganelle: A Fast and Versatile Toolkit for Accurate de Novo Assembly of Organelle Genomes. Genome Biol. 2020, 21, 241. [Google Scholar] [CrossRef]
- Bernt, M.; Donath, A.; Jühling, F.; Externbrink, F.; Florentz, C.; Fritzsch, G.; Pütz, J.; Middendorf, M.; Stadler, P.F. MITOS: Improved de Novo Metazoan Mitochondrial Genome Annotation. Mol. Phylogenet. Evol. 2013, 69, 313–319. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Larsson, A. AliView: A Fast and Lightweight Alignment Viewer and Editor for Large Datasets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef]
- Grant, J.R.; Stothard, P. The CGView Server: A Comparative Genomics Tool for Circular Genomes. Nucleic Acids Res. 2008, 36, 181–184. [Google Scholar] [CrossRef]
- Shen, W.; Le, S.; Li, Y.; Hu, F. SeqKit: A Cross-Platform and Ultrafast Toolkit for FASTA/Q File Manipulation. PLoS ONE 2016, 11, e0163962. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Nguyen, M.A.T.; Von Haeseler, A. Ultrafast Approximation for Phylogenetic Bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast Model Selection for Accurate Phylogenetic Estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. Mrbayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for Inference of Large Phylogenetic Trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010. [Google Scholar]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef]
- Boore, J.L.; Brown, W.M. Mitochondrial Genomes of Galathealinum, Helobdella, and Platynereis: Sequence and Gene Arrangement Comparisons Indicate That Pogonophora Is Not a Phylum and Annelida and Arthropoda Are Not Sister Taxa. Mol. Biol. Evol. 2000, 17, 87–106. [Google Scholar] [CrossRef]
- Jennings, R.M.; Halanych, K.M. Mitochondrial Genomes of Clymenella Torquata (Maldanidae) and Riftia Pachyptila (Siboglinidae): Evidence for Conserved Gene Order in Annelida. Mol. Biol. Evol. 2005, 22, 210–222. [Google Scholar] [CrossRef]
- Winnepenninckx, B.; Backeljau, T.; De Wachter, R. Phylogeny of Protostome Worms Derived from 18S RRNA Sequences. Mol. Biol. Evol. 1995, 12, 641–649. [Google Scholar]
- Hilário, A.; Johnson, S.B.; Cunha, M.R.; Vrijenhoek, R.C. High Diversity of Frenulates (Polychaeta: Siboglinidae) in the Gulf of Cadiz Mud Volcanoes: A DNA Taxonomy Analysis. Deep. Res. Part I Oceanogr. Res. Pap. 2010, 57, 143–150. [Google Scholar] [CrossRef]
- McCowin, M.F.; Collins, P.C.; Rouse, G.W. Updated Phylogeny of Vestimentifera (Siboglinidae, Polychaeta, Annelida) Based on Mitochondrial Genomes, with a New Species. Mol. Phylogenet. Evol. 2023, 187, 107872. [Google Scholar] [CrossRef]
- Li, Y.; Kocot, K.M.; Whelan, N.V.; Santos, S.R.; Waits, D.S.; Thornhill, D.J.; Halanych, K.M. Phylogenomics of Tubeworms (Siboglinidae, Annelida) and Comparative Performance of Different Reconstruction Methods. Zool. Scr. 2017, 46, 200–213. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Model (ML) | Model (BI) |
|---|---|---|
| ATP6 | GTR + F + G4 | nst = 6 rates = gammangammacat = 4 |
| ATP8 | TPM2u + F + I + G4 | nst = 2 rates = invgamma ngammacat = 4 |
| COX1 | GTR + F + R4 | nst = 6 rates = gamma ngammacat = 4 |
| COX2 | GTR + F + R4 | nst = 6 rates = gamma ngammacat = 4 |
| COX3 | GTR + F + I + G4 | nst = 6 rates = invgamma ngammacat = 4 |
| CYTB | TIM2 + F + I + G4 | nst = 6 rates = invgamma ngammacat = 4 |
| NAD1 | TIM2 + F + I + G4 | nst = 6 rates = invgamma ngammacat = 4 |
| NAD2 | TIM2 + F + I + G4 | nst = 6 rates = invgamma ngammacat = 4 |
| NAD3 | TIM + F + I + G4 | nst = 6 rates = invgamma ngammacat = 4 |
| NAD4 | TIM + F + I + G4 | nst = 6 rates = invgamma ngammacat = 4 |
| NAD4L | TIM2 + F + G4 | nst = 6 rates = gamma ngammacat = 4 |
| NAD5 | TIM2 + F + I + G4 | nst = 6 rates = invgamma ngammacat = 4 |
| NAD6 | HKY + F + I + G4 | nst = 2 rates = invgamma ngammacat = 4 |
| Gene | Length | fcd | scd | Length | fcd | scd |
|---|---|---|---|---|---|---|
| Siboglinum plumosum | Oligobrachia dogieli | |||||
| ATP6 | 681 | ATG | TAA | 681 | ATG | TAA |
| NAD5 | 1681 | ATT | TAA * | 1690 | ATG | TAA * |
| NAD4L | 288 | ATG | TAA | 288 | ATG | TAA |
| NAD4 | 1350 | ATG | TAA | 1353 | ATG | TAA |
| NAD1 | 921 | ATA | TAA | 924 | ATG | TAA |
| NAD3 | 339 | ATT | TAA | 354 | ATG | TAA |
| NAD2 | 990 | ATA | TAA * | 996 | ATG | TAA |
| COX1 | 1548 | ATG | TAA | 1544 | ATG | TAA * |
| COX2 | 675 | ATG | TAG | 687 | ATG | TAG |
| ATP8 | 153 | ATG | TAA | 156 | ATG | TAA |
| COX3 | 777 | ATC | TAA | 780 | ATG | TAA |
| NAD6 | 466 | ATG | TAA * | 469 | ATG | TAA * |
| COB | 1092 | ATG | TAA * | 1135 | ATG | TAA * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skalon, E.K.; Starunova, Z.I.; Petrov, S.A.; Smirnov, R.V.; Zaitseva, O.V.; Starunov, V.V. The Mitochondrial Genomes of Siboglinum plumosum and Oligobrachia dogieli (Annelida: Siboglinidae) and Their Phylogenetic Analysis. Genes 2024, 15, 77. https://doi.org/10.3390/genes15010077
Skalon EK, Starunova ZI, Petrov SA, Smirnov RV, Zaitseva OV, Starunov VV. The Mitochondrial Genomes of Siboglinum plumosum and Oligobrachia dogieli (Annelida: Siboglinidae) and Their Phylogenetic Analysis. Genes. 2024; 15(1):77. https://doi.org/10.3390/genes15010077
Chicago/Turabian StyleSkalon, Elizaveta K., Zinaida I. Starunova, Sergey A. Petrov, Roman V. Smirnov, Olga V. Zaitseva, and Viktor V. Starunov. 2024. "The Mitochondrial Genomes of Siboglinum plumosum and Oligobrachia dogieli (Annelida: Siboglinidae) and Their Phylogenetic Analysis" Genes 15, no. 1: 77. https://doi.org/10.3390/genes15010077
APA StyleSkalon, E. K., Starunova, Z. I., Petrov, S. A., Smirnov, R. V., Zaitseva, O. V., & Starunov, V. V. (2024). The Mitochondrial Genomes of Siboglinum plumosum and Oligobrachia dogieli (Annelida: Siboglinidae) and Their Phylogenetic Analysis. Genes, 15(1), 77. https://doi.org/10.3390/genes15010077