
Association between Variants in the OCA2-HERC2 Region and Blue Eye Colour in HERC2 rs12913832 AA and AG Individuals
 , , ,
, , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Population and Selection of Individuals for Genotyping
2.2. Eye Colour Categorisation and Quantitative Eye Colour Measurements
2.3. Quantitative Skin Colour Measurements
2.4. Probe Design and Sequencing the OCA2-HERC2 Region
2.5. Analysis of Sequencing Data
2.6. Variant Typing
3. Results
3.1. Sequencing of the HERC2-OCA2 Region and Discovery of Candidate Blue Eye Colour Variants
3.2. Candidate Blue Eye Colour Variants and Their Association with Eye Colour in the Norwegian Population
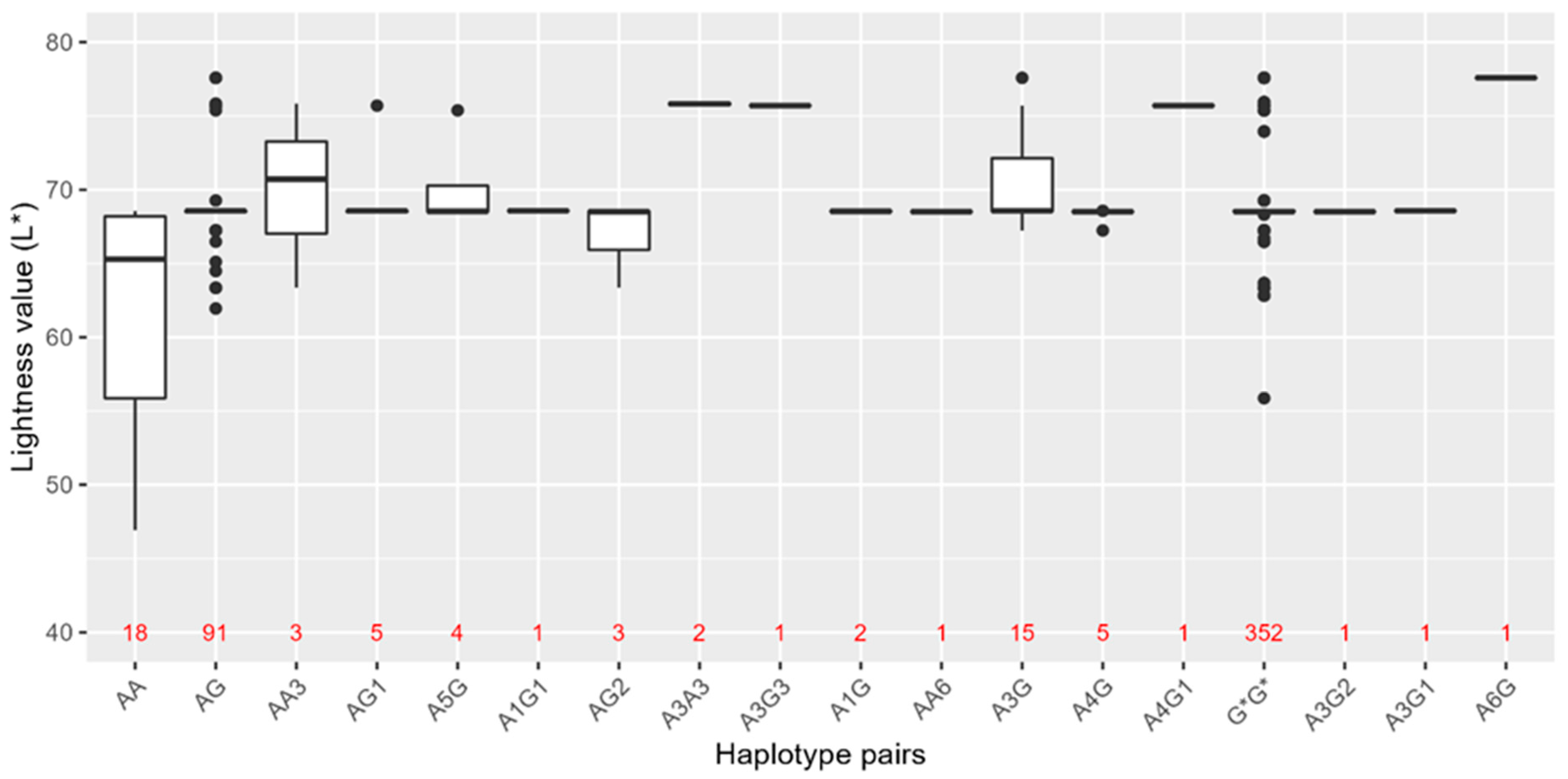
3.3. HERC2-OCA2 Haplotypes and Their Association with Skin Colour in the Norwegian Population
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sturm, R.A.; Larsson, M. Genetics of human iris colour and patterns. Pigment. Cell Melanoma Res. 2009, 22, 544–562. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Wen, B.; Kayser, M. Colorful DNA polymorphisms in humans. Semin. Cell Dev. Biol. 2013, 24, 562–575. [Google Scholar] [CrossRef] [PubMed]
- Visser, M.; Kayser, M.; Palstra, R.-J. HERC2 rs12913832 modulates human pigmentation by attenuating chromatin-loop formation between a long-range enhancer and the OCA2 promoter. Genome Res. 2012, 22, 446–455. [Google Scholar] [CrossRef]
- Sturm, R.A.; Duffy, D.L.; Zhao, Z.Z.; Leite, F.P.; Stark, M.S.; Hayward, N.K.; Martin, N.G.; Montgomery, G.W. A Single SNP in an Evolutionary Conserved Region within Intron 86 of the HERC2 Gene Determines Human Blue-Brown Eye Color. Am. J. Hum. Genet. 2008, 82, 424–431. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, M.P.; Paschou, P.; Grigorenko, E.; Gurwitz, D.; Barta, C.; Lu, R.-B.; Zhukova, O.V.; Kim, J.-J.; Siniscalco, M.; New, M.; et al. A global view of the OCA2-HERC2 region and pigmentation. Hum. Genet. 2011, 131, 683–696. [Google Scholar] [CrossRef] [PubMed]
- Walsh, S.; Wollstein, A.; Liu, F.; Chakravarthy, U.; Rahu, M.; Seland, J.H.; Soubrane, G.; Tomazzoli, L.; Topouzis, F.; Vingerling, J.R.; et al. DNA-based eye colour prediction across Europe with the IrisPlex system. Forensic Sci. Int. Genet. 2012, 6, 330–340. [Google Scholar] [CrossRef]
- Eiberg, H.; Troelsen, J.; Nielsen, M.; Mikkelsen, A.; Mengel-From, J.; Kjaer, K.W.; Hansen, L. Blue eye color in humans may be caused by a perfectly associated founder mutation in a regulatory element located within the HERC2 gene inhibiting OCA2 expression. Hum. Genet. 2008, 123, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Andersen, J.D.; Pietroni, C.; Johansen, P.; Andersen, M.M.; Pereira, V.; Børsting, C.; Morling, N. Importance of nonsynonymous OCA 2 variants in human eye color prediction. Mol. Genet. Genom. Med. 2016, 4, 420–430. [Google Scholar] [CrossRef]
- Andersen, J.D.; Johansen, P.; Harder, S.; Christoffersen, S.R.; Delgado, M.C.; Henriksen, S.T.; Nielsen, M.M.; Sørensen, E.; Ullum, H.; Hansen, T.; et al. Genetic analyses of the human eye colours using a novel objective method for eye colour classification. Forensic Sci. Int. Genet. 2013, 7, 508–515. [Google Scholar] [CrossRef]
- Salvo, N.M.; Janssen, K.; Kirsebom, M.K.; Meyer, O.S.; Berg, T.; Olsen, G.-H. Predicting eye and hair colour in a Norwegian population using Verogen’s ForenSeq™ DNA signature prep kit. Forensic Sci. Int. Genet. 2021, 56, 102620. [Google Scholar] [CrossRef]
- Liu, F.; van Duijn, K.; Vingerling, J.R.; Hofman, A.; Uitterlinden, A.G.; Janssens, A.C.J.; Kayser, M. Eye color and the prediction of complex phenotypes from genotypes. Curr. Biol. 2009, 19, R192–R193. [Google Scholar] [CrossRef] [PubMed]
- Duffy, D.; Montgomery, G.; Chen, W.; Zhao, Z.Z.; Le, L.; James, M.R.; Hayward, N.K.; Martin, N.; Sturm, R.A. A Three–Single-Nucleotide Polymorphism Haplotype in Intron 1 of OCA2 Explains Most Human Eye-Color Variation. Am. J. Hum. Genet. 2007, 80, 241–252. [Google Scholar] [CrossRef] [PubMed]
- Walsh, S.; Liu, F.; Ballantyne, K.N.; van Oven, M.; Lao, O.; Kayser, M. IrisPlex: A sensitive DNA tool for accurate prediction of blue and brown eye colour in the absence of ancestry information. Forensic Sci. Int. Genet. 2011, 5, 170–180. [Google Scholar] [CrossRef] [PubMed]
- Walsh, S.; Liu, F.; Wollstein, A.; Kovatsi, L.; Ralf, A.; Kosiniak-Kamysz, A.; Branicki, W.; Kayser, M. The HIrisPlex system for simultaneous prediction of hair and eye colour from DNA. Forensic Sci. Int. Genet. 2012, 7, 98–115. [Google Scholar] [CrossRef]
- Eaton, K.; Edwards, M.; Krithika, S.; Cook, G.; Norton, H.; Parra, E.J. Association study confirms the role of two OCA2 polymorphisms in normal skin pigmentation variation in East Asian populations. Am. J. Hum. Biol. 2015, 27, 520–525. [Google Scholar] [CrossRef]
- Meyer, O.; Salvo, N.; Kjærbye, A.; Kjersem, M.; Andersen, M.; Sørensen, E.; Ullum, H.; Janssen, K.; Morling, N.; Børsting, C.; et al. Prediction of Eye Colour in Scandinavians Using the EyeColour 11 (EC11) SNP Set. Genes 2021, 12, 821. [Google Scholar] [CrossRef]
- Schubert, M.; Lindgreen, S.; Orlando, L. AdapterRemoval v2: Rapid adapter trimming, identification, and read merging. BMC Res. Notes 2016, 9, 88. [Google Scholar] [CrossRef]
- Lindgreen, S. AdapterRemoval: Easy cleaning of next-generation sequencing reads. BMC Res. Notes 2012, 5, 337. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate long-read alignment with Burrows–Wheeler transform. Bioinformatics 2010, 26, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Li, H. Aligning Sequence Reads, Clone Sequences and Assembly Contigs with BWA-MEM. 2013. Available online: http://github.com/lh3/bwa (accessed on 31 January 2022).
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows–Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [PubMed]
- Quinlan, A.R.; Hall, I.M. BEDTools: A flexible suite of utilities for comparing genomic features. Bioinformatics 2010, 26, 841–842. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing. 2019. Available online: https://www.r-project.org/ (accessed on 10 March 2023).
- Barrett, J.C.; Fry, B.; Maller, J.; Daly, M.J. Haploview: Analysis and visualization of LD and haplotype maps. Bioinformatics 2005, 21, 263–265. [Google Scholar] [CrossRef] [PubMed]
- The ENCODE Project Consortium; Moore, J.E.; Purcaro, M.J.; Pratt, H.E.; Epstein, C.B.; Shoresh, N.; Adrian, J.; Kawli, T.; Davis, C.A.; Dobin, A.; et al. Expanded encyclopaedias of DNA elements in the human and mouse genomes. Nature 2020, 583, 699–710. [Google Scholar] [CrossRef] [PubMed]
- Stephens, M.; Smith, N.J.; Donnelly, P. A New Statistical Method for Haplotype Reconstruction from Population Data. Am. J. Hum. Genet. 2001, 68, 978–989. [Google Scholar] [CrossRef]
- Stephens, M.; Scheet, P. Accounting for Decay of Linkage Disequilibrium in Haplotype Inference and Missing-Data Imputation. Am. J. Hum. Genet. 2005, 76, 449–462. [Google Scholar] [CrossRef] [PubMed]
- Salvo, N.M.; Janssen, K.; Olsen, G.-H.; Berg, T.; Andersen, J.D. Association between copy number variations in the OCA2-HERC2 locus and human eye colour. Forensic Sci. Int. Genet. Suppl. Ser. 2022, 8, 82–84. [Google Scholar] [CrossRef]
- Meyer, O.S.; Børsting, C.; Andersen, J.D. Perception of blue and brown eye colours for forensic DNA phenotyping. Forensic Sci. Int. Genet. Suppl. Ser. 2019, 7, 476–477. [Google Scholar] [CrossRef]
- Ruiz, Y.; Phillips, C.; Gomez-Tato, A.; Alvarez-Dios, J.; de Cal, M.C.; Cruz, R.; Maroñas, O.; Söchtig, J.; Fondevila, M.; Rodriguez-Cid, M.; et al. Further development of forensic eye color predictive tests. Forensic Sci. Int. Genet. 2012, 7, 28–40. [Google Scholar] [CrossRef]
- Pietroni, C.; Andersen, J.D.; Johansen, P.; Andersen, M.M.; Harder, S.; Paulsen, R.R.; Børsting, C.; Morling, N. The effect of gender on eye colour variation in European populations and an evaluation of the IrisPlex prediction model. Forensic Sci. Int. Genet. 2014, 11, 1–6. [Google Scholar] [CrossRef]
- Hart, K.L.; Kimura, S.L.; Mushailov, V.; Budimlija, Z.M.; Prinz, M.; Wurmbach, E. Improved eye- and skin-color prediction based on 8 SNPs. Croat. Med. J. 2013, 54, 248–256. [Google Scholar] [CrossRef] [PubMed]
- Paparazzo, E.; Gozalishvili, A.; Lagani, V.; Geracitano, S.; Bauleo, A.; Falcone, E.; Passarino, G.; Montesanto, A. A new approach to broaden the range of eye colour identifiable by IrisPlex in DNA phenotyping. Sci. Rep. 2022, 12, 12803. [Google Scholar] [CrossRef] [PubMed]
- Sulem, P.; Gudbjartsson, D.; Stacey, S.N.; Helgason, A.; Rafnar, T.; Magnusson, K.P.; Manolescu, A.; Karason, A.; Palsson, A.; Thorleifsson, G.; et al. Genetic determinants of hair, eye and skin pigmentation in Europeans. Nat. Genet. 2007, 39, 1443–1452. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Kraft, P.; Nan, H.; Guo, Q.; Chen, C.; Qureshi, A.; Hankinson, S.E.; Hu, F.B.; Duffy, D.; Zhao, Z.Z.; et al. A Genome-Wide Association Study Identifies Novel Alleles Associated with Hair Color and Skin Pigmentation. PLoS Genet. 2008, 4, e1000074. [Google Scholar] [CrossRef]
- Sulem, P.; Gudbjartsson, D.F.; Stacey, S.N.; Helgason, A.; Rafnar, T.; Jakobsdottir, M.; Steinberg, S.; Gudjonsson, S.A.; Palsson, A.; Thorleifsson, G.; et al. Two newly identified genetic determinants of pigmentation in Europeans. Nat. Genet. 2008, 40, 835–837. [Google Scholar] [CrossRef] [PubMed]
- Kanetsky, P.A.; Swoyer, J.; Panossian, S.; Holmes, R.; Guerry, D.; Rebbeck, T.R. A Polymorphism in the Agouti Signaling Protein Gene Is Associated with Human Pigmentation. Am. J. Hum. Genet. 2002, 70, 770–775. [Google Scholar] [CrossRef]
- Frudakis, T.; Thomas, M.; Gaskin, Z.; Venkateswarlu, K.; Chandra, K.S.; Ginjupalli, S.; Gunturi, S.; Natrajan, S.; Ponnuswamy, V.K.; Ponnuswamy, K.N. Sequences Associated with Human Iris Pigmentation. Genetics 2003, 165, 2071–2083. [Google Scholar] [CrossRef]
- Pośpiech, E.; Karłowska-Pik, J.; Ziemkiewicz, B.; Kukla, M.; Skowron, M.; Wojas-Pelc, A.; Branicki, W. Further evidence for population specific differences in the effect of DNA markers and gender on eye colour prediction in forensics. Int. J. Leg. Med. 2016, 130, 923–934. [Google Scholar] [CrossRef]
- Le, L.; Escobar, I.E.; Ho, T.; Lefkovith, A.J.; Latteri, E.; Haltaufderhyde, K.D.; Dennis, M.K.; Plowright, L.; Sviderskaya, E.V.; Bennett, D.C.; et al. SLC45A2 protein stability and regulation of melanosome pH determine melanocyte pigmentation. Mol. Biol. Cell 2020, 31, 2687–2702. [Google Scholar] [CrossRef]
- Simcoe, M.; Valdes, A.; Liu, F.; Furlotte, N.A.; Evans, D.M.; Hemani, G.; Ring, S.M.; Smith, G.D.; Duffy, D.L.; Zhu, G.; et al. Genome-wide association study in almost 195,000 individuals identifies 50 previously unidentified genetic loci for eye color. Sci. Adv. 2021, 7, eabd1239. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| rs12913832 | Brown (Control Group) | Blue (Study Group) |
|---|---|---|
| AA | 21 | 3 |
| AG | 30 | 40 |
| Total | 51 | 43 |
| Candidate Variant | Chromosomal Position (GRCh38) | Frequency Brown (n = 46) 1 | Frequency Blue (n = 19) 1 | Variants in Strong LD 2 (r2 ≥ 0.8) |
|---|---|---|---|---|
| rs74409036 | 27,818,606 | 0.03 | 0.11 | rs78114576 |
| rs78544415 | 27,842,634 | 0.00 | 0.05 | rs79071800, rs28565430, rs28423991, rs74474678 and rs80177321 |
| rs72714116 | 28,083,061 | 0.00 | 0.08 | rs72714116 |
| rs191109490 | 28,120,403 | 0.00 | 0.05 | rs543971307 and rs145048438 |
| rs551217952 | 28,139,387 | 0.01 | 0.05 | rs182498200 and rs184120129 |
| rs62008729 | 28,173,140 | 0.01 | 0.08 | rs62008729 |
| Variant Allele Frequency | ||||
|---|---|---|---|---|
| rs ID | Reference Allele | Variant Allele | Frequency Norway (n = 519) | Frequency Europe (GnomAD) |
| rs74409036 | G | A | 0.031 | 0.052 |
| rs78544415 | C | T | 0.026 | 0.035 |
| rs72714116 | C | T | 0.004 | 0.020 |
| rs191109490 | A | C | 0.002 | 0.002 |
| rs551217952 | A | C | 0.003 | 0.003 |
| rs62008729 | C | T | 0.082 | 0.024 |
| Haplotypes a,c | rs74409036:A | rs78544415:T | rs74653330:T | rs121918166:T | rs72714116:T | rs191109490:C | rs551217952:C | Frequency NOR b (n = 519) |
|---|---|---|---|---|---|---|---|---|
| A | - | - | - | - | - | - | - | 0.135 |
| G | - | - | - | - | - | - | - | 0.772 |
| G1 | A | - | - | - | - | - | - | 0.028 |
| G2 | - | T | - | - | - | - | - | 0.025 |
| G3 | - | T | T | - | - | - | - | 0.001 |
| G4 | - | - | - | T | - | - | - | 0.001 |
| A1 | A | - | - | - | - | - | C | 0.003 |
| A3 | - | - | T | - | - | - | - | 0.024 |
| A4 | - | - | - | T | - | - | - | 0.006 |
| A5 | - | - | - | - | T | - | - | 0.004 |
| A6 | - | - | - | - | - | C | - | 0.002 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salvo, N.M.; Andersen, J.D.; Janssen, K.; Meyer, O.L.; Berg, T.; Børsting, C.; Olsen, G.-H. Association between Variants in the OCA2-HERC2 Region and Blue Eye Colour in HERC2 rs12913832 AA and AG Individuals. Genes 2023, 14, 698. https://doi.org/10.3390/genes14030698
Salvo NM, Andersen JD, Janssen K, Meyer OL, Berg T, Børsting C, Olsen G-H. Association between Variants in the OCA2-HERC2 Region and Blue Eye Colour in HERC2 rs12913832 AA and AG Individuals. Genes. 2023; 14(3):698. https://doi.org/10.3390/genes14030698
Chicago/Turabian StyleSalvo, Nina Mjølsnes, Jeppe Dyrberg Andersen, Kirstin Janssen, Olivia Luxford Meyer, Thomas Berg, Claus Børsting, and Gunn-Hege Olsen. 2023. "Association between Variants in the OCA2-HERC2 Region and Blue Eye Colour in HERC2 rs12913832 AA and AG Individuals" Genes 14, no. 3: 698. https://doi.org/10.3390/genes14030698
APA StyleSalvo, N. M., Andersen, J. D., Janssen, K., Meyer, O. L., Berg, T., Børsting, C., & Olsen, G.-H. (2023). Association between Variants in the OCA2-HERC2 Region and Blue Eye Colour in HERC2 rs12913832 AA and AG Individuals. Genes, 14(3), 698. https://doi.org/10.3390/genes14030698