
A Comprehensive Overview of Baboon Phylogenetic History
 ,
,  and
and 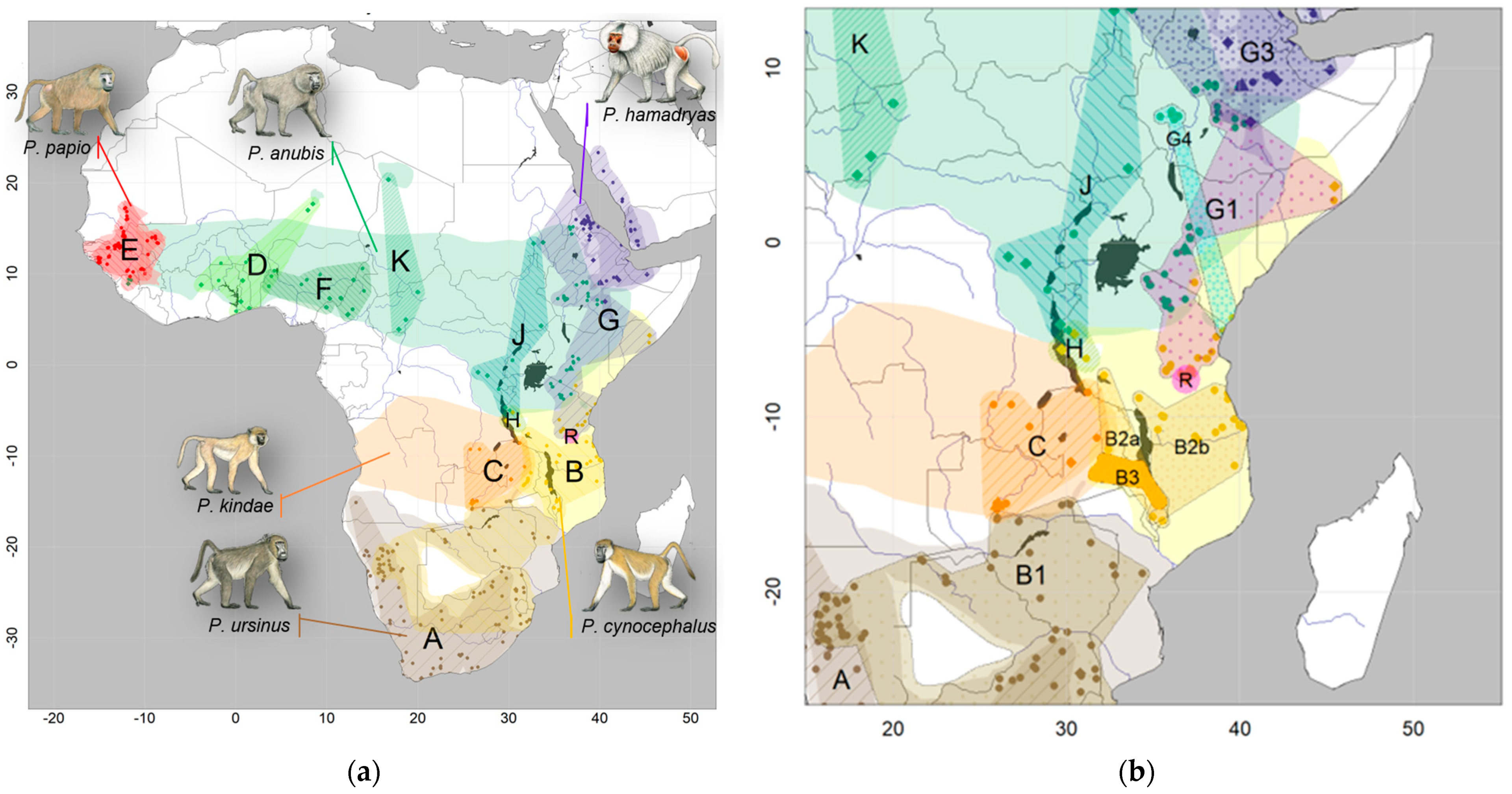{kind=link}
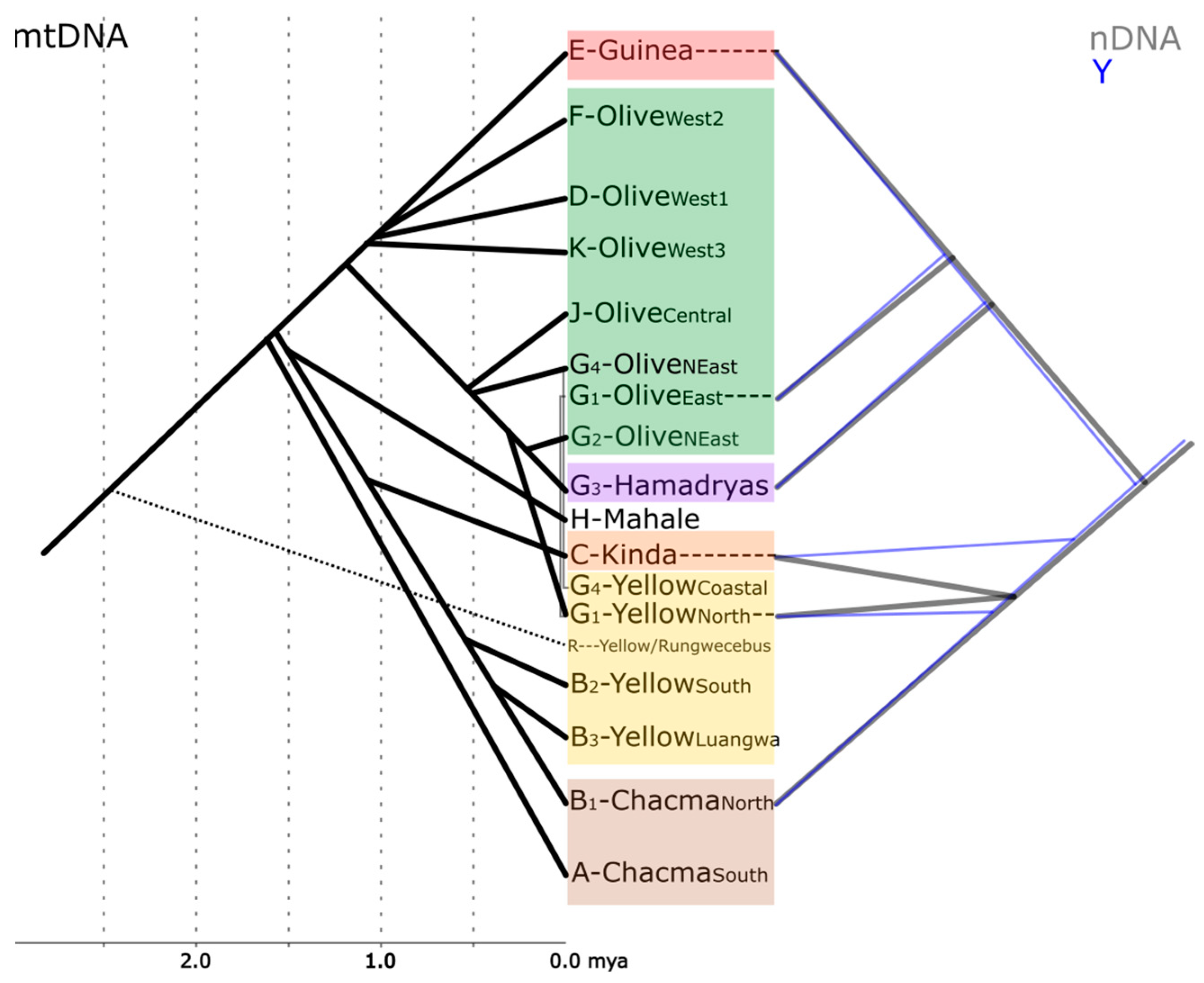{kind=link}
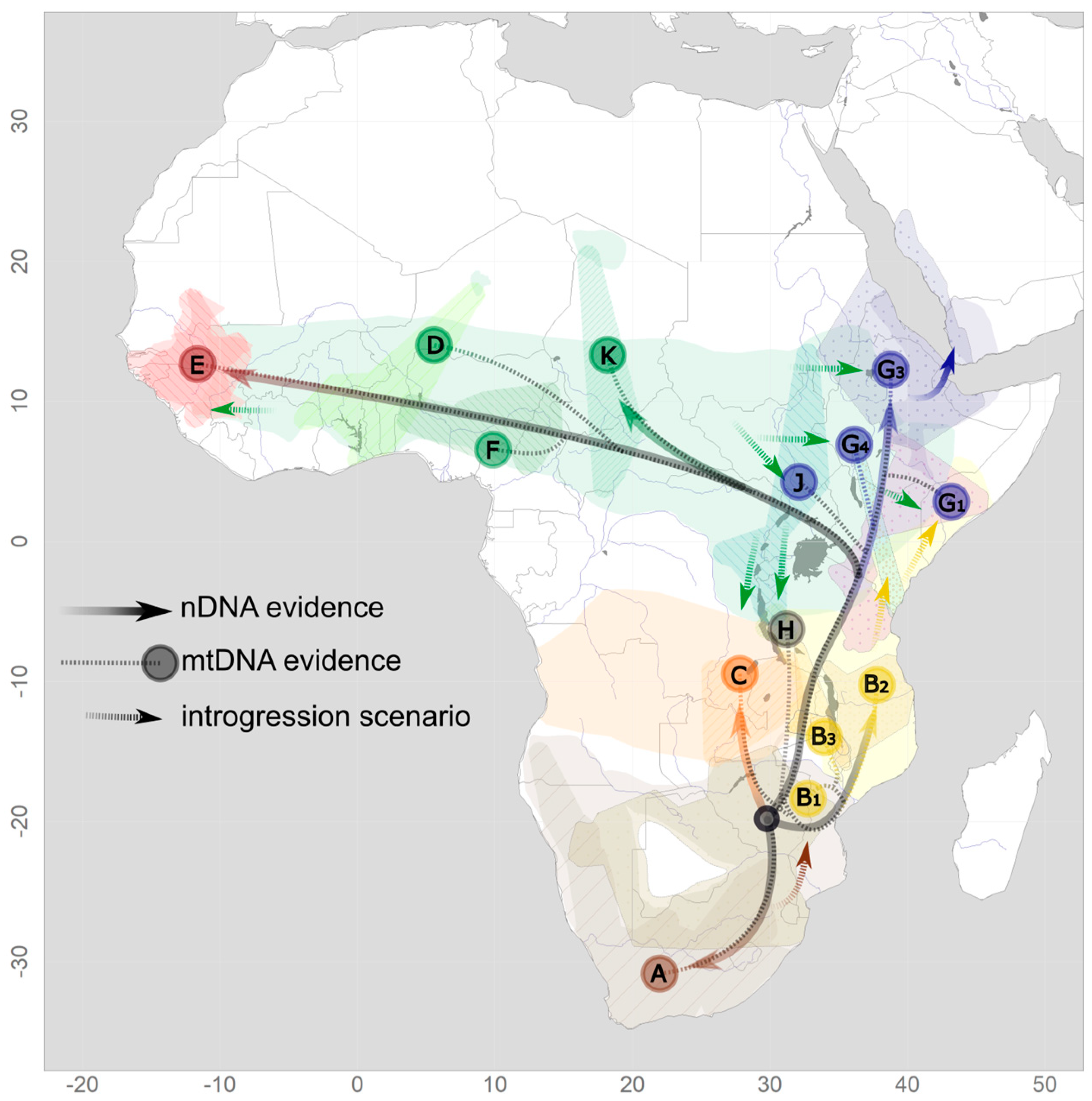{kind=link}
Abstract
1. Hybridization in Primates
2. Disentangling the Phylogeny of Baboons
3. The Cradle of Baboons: Introgressive Hybridization between Southern Baboons
4. The Two Main Clades Mingling: Past and Present Hybridization of Baboons in Eastern Africa
5. A Comprehensive Phylogeographic Scenario for Baboons
6. Outstanding Questions and Future Research Directions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arnold, M.L. Natural Hybridization and Evolution; Oxford University Press: New York, NY, USA, 1997. [Google Scholar]
- Holliday, T.W. Species Concepts, Reticulation, and Human Evolution. Curr. Anthropol. 2003, 44, 653–673. [Google Scholar] [CrossRef]
- Mallet, J. Hybridization as an Invasion of the Genome. Trends Ecol. Evol. 2005, 20, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Abbott, R.; Albach, D.; Ansell, S.; Arntzen, J.W.; Baird, S.J.E.; Bierne, N.; Boughman, J.; Brelsford, A.; Buerkle, C.A.; Buggs, R.; et al. Hybridization and Speciation. J. Evol. Biol. 2013, 26, 229–246. [Google Scholar] [CrossRef]
- Seehausen, O. Conservation: Losing Biodiversity by Reverse Speciation. Curr. Biol. 2006, 16, R334–R337. [Google Scholar] [CrossRef]
- Butlin, R.K. Reinforcement: An Idea Evolving. Trends Ecol. Evol. 1995, 10, 432–434. [Google Scholar] [CrossRef]
- Barton, N.H.; Hewitt, G.M. Analysis of Hybrid Zones. Annu. Rev. Ecol. Syst. 1985, 16, 113–148. [Google Scholar] [CrossRef]
- Shurtliff, Q.R. Mammalian Hybrid Zones: A Review. Mamm. Rev. 2013, 43, 1–21. [Google Scholar] [CrossRef]
- Arnold, M.L. Evolution Through Genetic Exchange; Oxford University Press: New York, NY, USA, 2006. [Google Scholar]
- Hedrick, P.W. Adaptive Introgression in Animals: Examples and Comparison to New Mutation and Standing Variation as Sources of Adaptive Variation. Mol. Ecol. 2013, 22, 4606–4618. [Google Scholar] [CrossRef]
- Tricou, T.; Tannier, E.; de Vienne, D.M. Ghost Lineages Can Invalidate or Even Reverse Findings Regarding Gene Flow. PLoS Biol. 2022, 20, e3001776. [Google Scholar] [CrossRef] [PubMed]
- Zinner, D.; Arnold, M.L.; Roos, C. The Strange Blood: Natural Hybridization in Primates. Evol. Anthropol. 2011, 20, 96–103. [Google Scholar] [CrossRef]
- Cortés-Ortiz, L.; Roos, C.; Zinner, D. Introduction to Special Issue on Primate Hybridization and Hybrid Zones. Int. J. Primatol. 2019, 40, 1–8. [Google Scholar] [CrossRef]
- Tung, J.; Barreiro, L.B. The Contribution of Admixture to Primate Evolution. Curr. Opin. Genet. Dev. 2017, 47, 61–68. [Google Scholar] [CrossRef]
- Arnold, M.L.; Meyer, A. Natural Hybridization in Primates: One Evolutionary Mechanism. Zoology 2006, 109, 261–276. [Google Scholar] [CrossRef]
- Zinner, D.; Chuma, I.S.; Knauf, S.; Roos, C. Inverted Intergeneric Introgression between Critically Endangered Kipunjis and Yellow Baboons in Two Disjunct Populations. Biol. Lett. 2018, 14, 20170729. [Google Scholar] [CrossRef]
- Vilgalys, T.P.; Fogel, A.S.; Anderson, J.A.; Mututua, R.S.; Warutere, J.K.; Siodi, I.L.; Kim, S.Y.; Voyles, T.N.; Robinson, J.A.; Wall, J.D.; et al. Selection against Admixture and Gene Regulatory Divergence in a Long-Term Primate Field Study. Science 2022, 377, 635–641. [Google Scholar] [CrossRef]
- Rogers, J.; Raveendran, M.; Harris, R.A.; Mailund, T.; Leppälä, K.; Athanasiadis, G.; Schierup, M.H.; Cheng, J.; Munch, K.; Walker, J.A.; et al. The Comparative Genomics and Complex Population History of Papio Baboons. Sci. Adv. 2019, 5, eaau6947. [Google Scholar] [CrossRef]
- Nichols, R. Gene Trees and Species Trees Are Not the Same. Trends Ecol. Evol. 2001, 16, 358–364. [Google Scholar] [CrossRef]
- Maddison, W.P. Gene Trees in Species Trees. Syst. Biol. 1997, 46, 523–536. [Google Scholar] [CrossRef]
- Abi-Rached, L.; Jobin, M.J.; Kulkarni, S.; Mcwhinnie, A.; Dalva, K.; Gragert, L.; Gharizadeh, B.; Luo, M.; Plummer, F.A.; Kimani, J.; et al. The Shaping of Modern Human Immune Systems by Multiregional Admixture with Archaic Humans. Science 2011, 334, 89–94. [Google Scholar] [CrossRef]
- Dannemann, M.; Racimo, F. Something Old, Something Borrowed: Admixture and Adaptation in Human Evolution. Curr. Opin. Genet. Dev. 2018, 53, 1–8. [Google Scholar] [CrossRef]
- Chiou, K.L.; Bergey, C.M.; Burrell, A.S.; Disotell, T.R.; Rogers, J.; Jolly, C.J.; Phillips-Conroy, J.E. Genome-Wide Ancestry and Introgression in a Zambian Baboon Hybrid Zone. Mol. Ecol. 2021, 30, 1907–1920. [Google Scholar] [CrossRef]
- Wall, J.D.; Schlebusch, S.A.; Alberts, S.C.; Cox, L.A.; Snyder-Mackler, N.; Nevonen, K.A.; Carbone, L.; Tung, J. Genomewide Ancestry and Divergence Patterns from Low-Coverage Sequencing Data Reveal a Complex History of Admixture in Wild Baboons. Mol. Ecol. 2016, 25, 3469–3483. [Google Scholar] [CrossRef]
- Hill, W.C.O. Primates: Comparative Anatomy and Taxonomy, Vol. 8; Edinburgh University Press: Edinburgh, UK, 1970. [Google Scholar]
- Groves, C.P. Primate Taxonomy; Smithsonian Institution Press: Washington, DC, USA, 2001. [Google Scholar]
- Jolly, C.J. Species, Subspecies and Baboon Systematics. In Species, Species Concepts and Primate Evolution; Kimbel, W.H., Martin, L.B., Eds.; Plenum Press: New York, NY, USA, 1993; pp. 67–107. [Google Scholar]
- Anandam, M.V.; Bennett, E.L.; Davenport, T.R.B.; Davies, N.J.; Detwiler, K.M.; Engelhardt, A.; Eudey, A.A.; Gadsby, E.L.; Groves, C.P.; Healy, A.; et al. Species Accounts of Cercopithecidae. In Handbook of the Mammals of the World Vol. 3 Primates; Mittermeier, R.A., Rylands, A.B., Wilson, D.E., Eds.; Lynx Edicions: Barcelona, Spain, 2013; pp. 628–753. [Google Scholar]
- Newman, T.K.; Jolly, C.J.; Rogers, J. Mitochondrial Phylogeny and Systematics of Baboons (Papio). Am. J. Phys. Anthropol. 2004, 124, 17–27. [Google Scholar] [CrossRef]
- Wildman, D.E.; Bergman, T.J.; Al-Aghbari, A.; Sterner, K.N.; Newman, T.K.; Pines, M.; Jolly, C.J.; Disotell, T.R. Mitochondrial Evidence for the Origin of Hamadryas Baboons. Mol. Phylogenet. Evol. 2004, 32, 287–296. [Google Scholar] [CrossRef]
- Jolly, C.J. The Origins and Specialisations of the Long-Faced Cercopithecoidea. Ph.D. Thesis, University of London, London, UK, 1965. [Google Scholar]
- Zinner, D.; Groeneveld, L.F.; Keller, C.; Roos, C. Mitochondrial Phylogeography of Baboons (Papio spp.)—Indication for Introgressive Hybridization? BMC Evol. Biol. 2009, 9, 83. [Google Scholar] [CrossRef]
- Burrell, A.S. Phylogenetics and Population Genetics of Central African Baboons. Ph.D. Thesis, New York University, New York, NY, USA, 2008. [Google Scholar]
- Zinner, D.; Buba, U.; Roos, C. Pan-African Voyagers: The Phylogeography of Baboons. In Primates of Gashaka. Socioecology and Conservation in Nigeria’s Biodiversity Hotspot; Sommer, V., Roos, C., Eds.; Springer: New York, NY, USA, 2011; pp. 267–307. [Google Scholar]
- Zinner, D.; Keller, C.; Nyahongo, J.W.; Butynski, T.M.; Jong, Y.A.D.; Roos, C. Distribution of Mitochondrial Clades and Morphotypes of Baboons Papio spp. (Primates: Cercopithecidae) in Eastern Africa. J. East Afr. Nat. Hist. 2015, 104, 143–168. [Google Scholar] [CrossRef]
- Zinner, D.; Wertheimer, J.; Liedigk, R.; Groeneveld, L.F.; Roos, C. Baboon Phylogeny as Inferred from Complete Mitochondrial Genomes. Am. J. Phys. Anthropol. 2013, 150, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Roos, C.; Knauf, S.; Chuma, I.S.; Maille, A.; Callou, C.; Sabin, R.; Portela Miguez, R.; Zinner, D. New Mitogenomic Lineages in Papio Baboons and Their Phylogeographic Implications. Am. J. Phys. Anthropol. 2021, 174, 407–417. [Google Scholar] [CrossRef]
- Burrell, A.S.; Jolly, C.J.; Tosi, A.J.; Disotell, T.R. Mitochondrial Evidence for the Hybrid Origin of the Kipunji, Rungwecebus kipunji (Primates: Papionini). Mol. Phylogenet. Evol. 2009, 51, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Zinner, D.; Arnold, M.L.; Roos, C. Is the New Primate Genus Rungwecebus a Baboon? PLoS ONE 2009, 4, e4859. [Google Scholar] [CrossRef] [PubMed]
- Roberts, T.E.; Davenport, T.R.B.; Hildebrandt, K.B.P.; Jones, T.; Stanley, W.T.; Sargis, E.J.; Olson, L.E. The Biogeography of Introgression in the Critically Endangered African Monkey Rungwecebus kipunji. Biol. Lett. 2010, 6, 233–237. [Google Scholar] [CrossRef]
- Haus, T.; Akom, E.; Agwanda, B.; Hofreiter, M.; Roos, C.; Zinner, D. Mitochondrial Diversity and Distribution of African Green Monkeys (Chlorocebus Gray, 1870). Am. J. Primatol. 2013, 75, 350–360. [Google Scholar] [CrossRef]
- Boissinot, S.; Alvarez, L.; Giraldo-Ramirez, J.; Tollis, M. Neutral Nuclear Variation in Baboons (genus Papio) Provides Insights into Their Evolutionary and Demographic Histories. Am. J. Phys. Anthropol. 2014, 155, 621–634. [Google Scholar] [CrossRef]
- Santander, C.; Molinaro, L.; Mutti, G.; Martínez, F.I.; Mathe, J.; Ferreira da Silva, M.J.; Caldon, M.; Oteo-Garcia, G.; Aldeias, V.; Archer, W.; et al. Genomic Variation in Baboons from Central Mozambique Unveils Complex Evolutionary Relationships with Other Papio Species. BMC Ecol. Evol. 2022, 22, 1–16. [Google Scholar] [CrossRef]
- Jordan, V.E.; Walker, J.A.; Beckstrom, T.O.; Steely, C.J.; McDaniel, C.L.; St Romain, C.P.; Baboon Genome Analysis Consortium; Worley, K.C.; Phillips-Conroy, J.; Jolly, C.J.; et al. A Computational Reconstruction of Papio Phylogeny Using Alu Insertion Polymorphisms. Mob. DNA 2018, 9, 13. [Google Scholar] [CrossRef]
- Walker, J.A.; Jordan, V.E.; Storer, J.M.; Steely, C.J.; Gonzalez-Quiroga, P.; Beckstrom, T.O.; Rewerts, L.C.; St Romain, C.P.; Rockwell, C.E.; Rogers, J.; et al. Alu Insertion Polymorphisms Shared by Papio Baboons and Theropithecus Gelada Reveal an Intertwined Common Ancestry. Mob. DNA 2019, 10, 46. [Google Scholar] [CrossRef]
- Dolotovskaya, S.; Torroba Bordallo, J.; Haus, T.; Noll, A.; Hofreiter, M.; Zinner, D.; Roos, C. Comparing Mitogenomic Timetrees for Two African Savannah Primate Genera (Chlorocebus and Papio). Zool. J. Linn. Soc. 2017, 181, 471–483. [Google Scholar] [CrossRef]
- Liedigk, R.; Roos, C.; Brameier, M.; Zinner, D. Mitogenomics of the Old World Monkey Tribe Papionini. BMC Evol. Biol. 2014, 14, 176. [Google Scholar] [CrossRef]
- Grathwol, F.; Roos, C.; Zinner, D.; Hume, B.; Porcier, S.; Berthet, D.; Cuisin, J.; Merker, S.; Ottoni, C.; Van Neer, W.; et al. Adulis and the Transshipment of Baboons during Classical Antiquity. Unpublished manuscript. OSF. [CrossRef]
- Keller, C.; Roos, C.; Groeneveld, L.F.; Fischer, J.; Zinner, D. Introgressive Hybridization in Southern African Baboons Shapes Patterns of mtDNA Variation. Am. J. Phys. Anthropol. 2010, 142, 125–136. [Google Scholar] [CrossRef]
- Sithaldeen, R.; Bishop, J.M.; Ackermann, R.R. Mitochondrial DNA Analysis Reveals Plio-Pleistocene Diversification within the Chacma Baboon. Mol. Phylogenet. Evol. 2009, 53, 1042–1048. [Google Scholar] [CrossRef]
- Sithaldeen, R.; Ackermann, R.R.; Bishop, J.M. Pleistocene Aridification Cycles Shaped the Contemporary Genetic Architecture of Southern African Baboons. PLoS ONE 2015, 10, e0123207. [Google Scholar] [CrossRef]
- Stone, O.M.L.; Herries, A.I.R.; Brink, J.S.; Laffan, S.W. The Chacma Baboon (Papio ursinus) through Time: A Model of Potential Core Habitat Regions during a Glacial–interglacial Cycle. Evol. Ecol. 2016, 30, 755–782. [Google Scholar] [CrossRef]
- Martinez, F.I.; Capelli, C.; Ferreira da Silva, M.J.; Aldeias, V.; Alemseged, Z.; Archer, W.; Bamford, M.; Biro, D.; Bobe, R.; Braun, D.R.; et al. A Missing Piece of the Papio Puzzle: Gorongosa Baboon Phenostructure and Intrageneric Relationships. J. Hum. Evol. 2019, 130, 1–20. [Google Scholar] [CrossRef]
- deMenocal, P.B. African Climate Change and Faunal Evolution during the Pliocene-Pleistocene. Earth Planet. Sci. Lett. 2004, 220, 3–24. [Google Scholar] [CrossRef]
- Maslin, M.A.; Brierley, C.M.; Milner, A.M.; Shultz, S.; Trauth, M.H.; Wilson, K.E. East African Climate Pulses and Early Human Evolution. Quat. Sci. Rev. 2014, 101, 1–17. [Google Scholar] [CrossRef]
- Trauth, M.H.; Bergner, A.G.N.; Foerster, V.; Junginger, A.; Maslin, M.A.; Schaebitz, F. Episodes of Environmental Stability versus Instability in Late Cenozoic Lake Records of Eastern Africa. J. Hum. Evol. 2015, 87, 21–31. [Google Scholar] [CrossRef]
- Lorenzen, E.D.; Heller, R.; Siegismund, H.R. Comparative Phylogeography of African Savannah Ungulates. Mol. Ecol. 2012, 21, 3656–3670. [Google Scholar] [CrossRef]
- Chala, D.; Roos, C.; Svenning, J.C.; Zinner, D. Species-Specific Effects of Climate Change on the Distribution of Suitable Baboon Habitats—Ecological Niche Modeling of Current and Last Glacial Maximum Conditions. J. Hum. Evol. 2019, 132, 215–226. [Google Scholar] [CrossRef]
- Corti, G. Continental Rift Evolution: From Rift Initiation to Incipient Break-up in the Main Ethiopian Rift, East Africa. Earth-Sci. Rev. 2009, 96, 1–53. [Google Scholar] [CrossRef]
- Mollel, G.F.; Swisher, C.C., 3rd. The Ngorongoro Volcanic Highland and Its Relationships to Volcanic Deposits at Olduvai Gorge and East African Rift Volcanism. J. Hum. Evol. 2012, 63, 274–283. [Google Scholar] [CrossRef]
- Hutchison, W.; Fusillo, R.; Pyle, D.M.; Mather, T.A.; Blundy, J.D.; Biggs, J.; Yirgu, G.; Cohen, B.E.; Brooker, R.A.; Barfod, D.N.; et al. A Pulse of Mid-Pleistocene Rift Volcanism in Ethiopia at the Dawn of Modern Humans. Nat. Commun. 2016, 7, 13192. [Google Scholar] [CrossRef]
- Butynski, T.M.; de Jong, Y.A.; Perkin, A.W.; Bearder, S.K.; Honess, P.E. Taxonomy, Distribution, and Conservation Status of Three Species of Dwarf Galagos (Galagoides) in Eastern Africa. Primate Conserv. 2006, 2006, 63–79. [Google Scholar] [CrossRef]
- McDonald, M.M.; Cunneyworth, P.M.K.; Anderson, A.G.; Wroblewski, E. Mitochondrial Genetic Diversity and Divergence Dating of Angolan Colobus Monkeys (Colobus angolensis) in the Eastern Forests of Kenya and Tanzania: Implications for Subspeciation and Reconstructing Historical Biogeography. Am. J. Primatol. 2022, 84, e23384. [Google Scholar] [CrossRef]
- Alberts, S.C.; Altmann, J. Immigration and Hybridization Patterns of Yellow and Anubis Baboons in and around Amboseli, Kenya. Am. J. Primatol. 2001, 53, 139–154. [Google Scholar] [CrossRef]
- Bergey, C.M.; Phillips-Conroy, J.E.; Disotell, T.R.; Jolly, C.J. Dopamine Pathway Is Highly Diverged in Primate Species That Differ Markedly in Social Behavior. Proc. Natl. Acad. Sci. USA 2016, 113, 201525530. [Google Scholar] [CrossRef]
- Phillips-Conroy, J.E.; Jolly, C.J. Male Dispersal and Philopatry in the Awash Baboon Hybrid Zone. Primate Rep. 2004, 68, 27–52. [Google Scholar]
- Charpentier, M.; Cherel, E.; Renoult, J.; Jenkins, T.; Benoit, L.; Barthes, N.; Alberts, S.C.; Tung, J. Genetic Structure in a Dynamic Baboon Hybrid Zone Corroborates Behavioral Observations in a Hybrid Population. Mol. Ecol. 2012, 21, 715–731. [Google Scholar] [CrossRef]
- Grathwol, F.; Roos, C.; Zinner, D.; Cuisin, J.; Kopp, G.H. New Olive Baboon Mitogenomes from Central and West Africa. University of Konstanz: Konstanz, Germany, manuscript in preparation. 2023. [Google Scholar]
- Frost, S.R. Evolution of the Cercopithecidae. In The International Encyclopedia of Primatology, Vol.2; Fuentes, a., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2017; pp. 1–3. [Google Scholar]
- Gilbert, C.C.; Frost, S.R.; Pugh, K.D.; Anderson, M.; Delson, E. Evolution of the Modern Baboon (Papio hamadryas): A Reassessment of the African Plio-Pleistocene Record. J. Hum. Evol. 2018, 122, 38–69. [Google Scholar] [CrossRef]
- Jablonski, N.G. Fossil Old World Monkeys: The Late Neogene Radiation. In The Primate Fossil Record; Hartwig, W.C., Ed.; Cambridge University Press: Cambridge, UK, 2002; pp. 255–299. [Google Scholar]
- Brasil, M.F.; Monson, T.A.; Taylor, C.E.; Yohler, R.M.; Hlusko, L.J. A Pleistocene Assemblage of near-Modern Papio hamadryas from the Middle Awash Study Area, Afar Rift, Ethiopia. Am. J. Biol. Anthropol. 2023, 180, 48–76. [Google Scholar] [CrossRef]
- Jablonski, N.G.; Frost, S.R. Cercopithecoidea. In Cenozoic Mammals of Africa; Werdelin, L., Sanders, W.J., Eds.; University of California Press: Berkeley, CA, USA, 2010; pp. 393–428. [Google Scholar]
- Liu, Y.; Mao, X.; Krause, J.; Fu, Q. Insights into Human History from the First Decade of Ancient Human Genomics. Science 2021, 373, 1479–1484. [Google Scholar] [CrossRef]
- Zinner, D.; Klapproth, M.; Schell, A.; Ohrndorf, L.; Chala, D.; Ganzhorn, J.U.; Fischer, J. Comparative Ecology of Guinea Baboons (Papio papio). Primate Biol. 2021, 8, 19–35. [Google Scholar] [CrossRef]
- Rowan, J.; Kamilar, J.M.; Beaudrot, L.; Reed, K.E. Strong Influence of Palaeoclimate on the Structure of Modern African Mammal Communities. Proc. R. Soc. B. 2016, 283, 20161207. [Google Scholar] [CrossRef]
- Fischer, J.; Higham, J.P.; Alberts, S.C.; Barrett, L.; Beehner, J.C.; Bergman, T.J.; Carter, A.J.; Collins, A.; Elton, S.; Fagot, J.; et al. Insights into the Evolution of Social Systems and Species from Baboon Studies. eLife 2019, 8, e50989. [Google Scholar] [CrossRef]
- Jolly, C.J. Philopatry at the Frontier: A Demographically Driven Scenario for the Evolution of Multilevel Societies in Baboons (Papio). J. Hum. Evol. 2020, 146, 102819. [Google Scholar] [CrossRef]
- Atickem, A.; Stenseth, N.C.; Drouilly, M.; Bock, S.; Roos, C.; Zinner, D. Deep Divergence among Mitochondrial Lineages in African Jackals. Zool. Scr. 2018, 47, 1–8. [Google Scholar] [CrossRef]
- Kingdon, J.; Happold, D.; Butynski, T.M.; Hoffmann, M.; Happold, M.; Kalina, J. (Eds.) Mammals of Africa. Vol. I-VI.; Bloomsbury Natural History: London, UK, 2013. [Google Scholar]
- Kopp, G.H.; Ferreira da Silva, M.J.; Fischer, J.; Brito, J.C.; Regnaut, S.; Roos, C.; Zinner, D. The Influence of Social Systems on Patterns of Mitochondrial DNA Variation in Baboons. Int. J. Primatol. 2014, 35, 210–225. [Google Scholar] [CrossRef]
- Kopp, G.H. Gene Flow Dynamics in Baboons: The Influence of Social Systems. Ph.D. Thesis, Georg-August-University, Göttingen, Germany, 2015. [Google Scholar]
- Snyder-Mackler, N.; Majoros, W.H.; Yuan, M.L.; Shaver, A.O.; Gordon, J.B.; Kopp, G.H.; Schlebusch, S.A.; Wall, J.D.; Alberts, S.C.; Mukherjee, S.; et al. Efficient Genome-Wide Sequencing and Low Coverage Pedigree Analysis from Non-Invasively Collected Samples. Genetics 2016, 203, 699–714. [Google Scholar] [CrossRef]
- Chiou, K.L.; Bergey, C.M. Methylation-Based Enrichment Facilitates Low-Cost, Noninvasive Genomic Scale Sequencing of Populations from Feces. Sci. Rep. 2018, 8, 1975. [Google Scholar] [CrossRef]
- Orkin, J.D.; Montague, M.J.; Tejada-Martinez, D.; de Manuel, M.; Del Campo, J.; Cheves Hernandez, S.; Di Fiore, A.; Fontsere, C.; Hodgson, J.A.; Janiak, M.C.; et al. The Genomics of Ecological Flexibility, Large Brains, and Long Lives in Capuchin Monkeys Revealed with fecalFACS. Proc. Natl. Acad. Sci. USA 2021, 118, e2010632118. [Google Scholar] [CrossRef]
- Burrell, A.S.; Disotell, T.R.; Bergey, C.M. The Use of Museum Specimens with High-Throughput DNA Sequencers. J. Hum. Evol. 2015, 79, 35–44. [Google Scholar] [CrossRef]
- Bi, K.; Linderoth, T.; Vanderpool, D.; Good, J.M.; Nielsen, R.; Moritz, C. Unlocking the Vault: Next-Generation Museum Population Genomics. Mol. Ecol. 2013, 22, 6018–6032. [Google Scholar] [CrossRef]
- Rowe, K.C.; Singhal, S.; Macmanes, M.D.; Ayroles, J.F.; Morelli, T.L.; Rubidge, E.M.; Bi, K.; Moritz, C.C. Museum Genomics: Low-Cost and High-Accuracy Genetic Data from Historical Specimens. Mol. Ecol. Resour. 2011, 11, 1082–1092. [Google Scholar] [CrossRef]
- Paijmans, J.L.A.; Gilbert, M.T.P.; Hofreiter, M. Mitogenomic Analyses from Ancient DNA. Mol. Phylogenet. Evol. 2012, 69, 404–416. [Google Scholar] [CrossRef]
- Roos, C.; Portela Miguez, R.; Sabin, R.; Louis, E.E.; Hofreiter, M.; Zinner, D. Mitogenomes of Historical Type Specimens Unravel the Taxonomy of Sportive Lemurs (Lepilemur spp.) in Northwest Madagascar. Zool. Res. 2021, 42, 428–432. [Google Scholar] [CrossRef]
- Chiou, K.L.; Bergey, C.M.; Burrell, A.S.; Disotell, T.R.; Rogers, J.; Jolly, C.J.; Phillips-Conroy, J.E. Genomic Signatures of Extreme Body Size Divergence in Baboons. bioRxiv 2019, 578740. [Google Scholar]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kopp, G.H.; Sithaldeen, R.; Trede, F.; Grathwol, F.; Roos, C.; Zinner, D. A Comprehensive Overview of Baboon Phylogenetic History. Genes 2023, 14, 614. https://doi.org/10.3390/genes14030614
Kopp GH, Sithaldeen R, Trede F, Grathwol F, Roos C, Zinner D. A Comprehensive Overview of Baboon Phylogenetic History. Genes. 2023; 14(3):614. https://doi.org/10.3390/genes14030614
Chicago/Turabian StyleKopp, Gisela H., Riashna Sithaldeen, Franziska Trede, Franziska Grathwol, Christian Roos, and Dietmar Zinner. 2023. "A Comprehensive Overview of Baboon Phylogenetic History" Genes 14, no. 3: 614. https://doi.org/10.3390/genes14030614
APA StyleKopp, G. H., Sithaldeen, R., Trede, F., Grathwol, F., Roos, C., & Zinner, D. (2023). A Comprehensive Overview of Baboon Phylogenetic History. Genes, 14(3), 614. https://doi.org/10.3390/genes14030614