


Genome-Wide Association Study of Arsenic Accumulation in Polished Rice

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Field and Pot Experiments
2.2. Sampling and As Determination
2.3. SLAF-Seq Library Construction, Sequencing Screening, and SNP Calling Analysis
2.4. Genome-Wide Association Study
2.5. Candidate Gene Prediction and Detection
3. Results
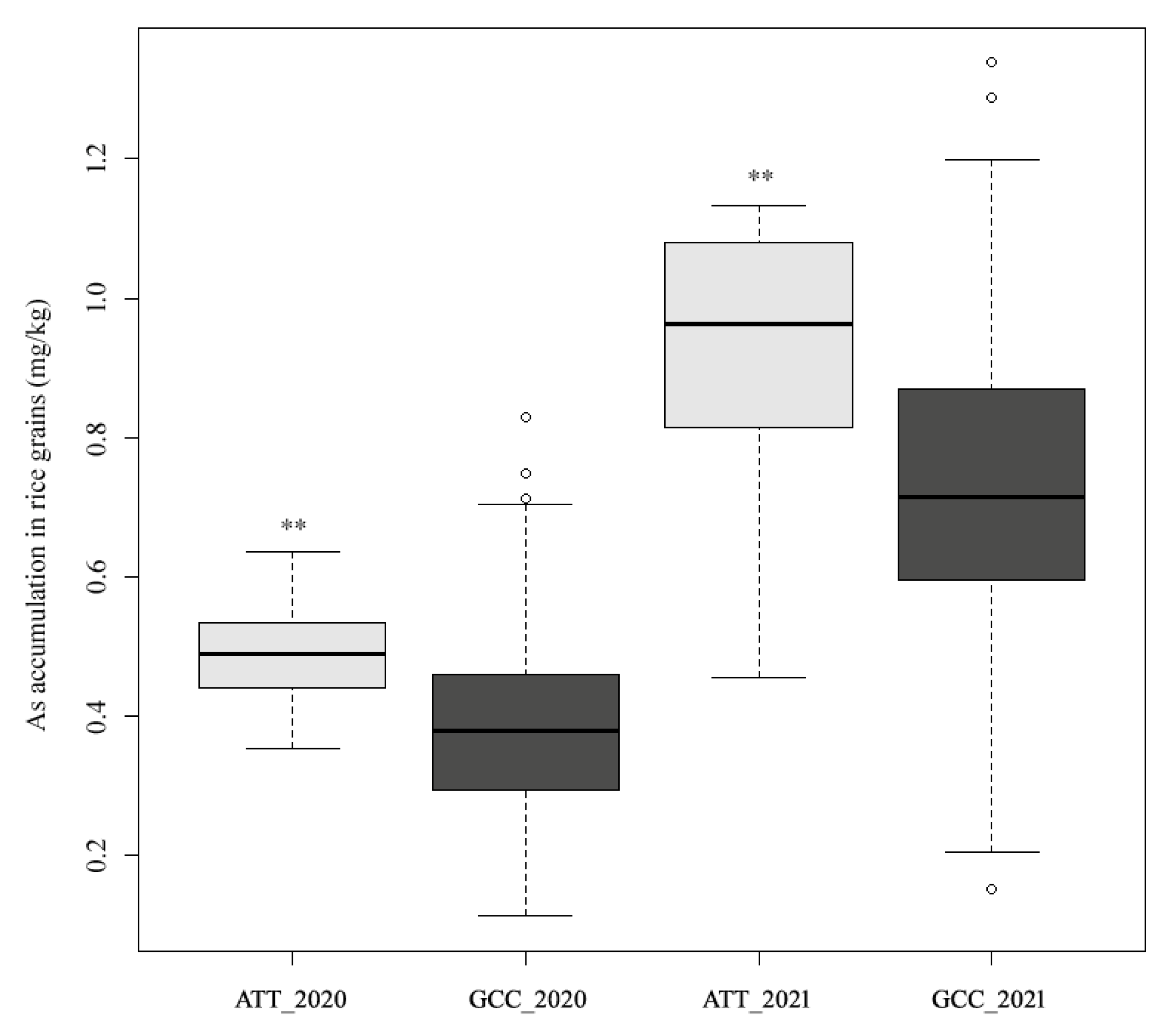
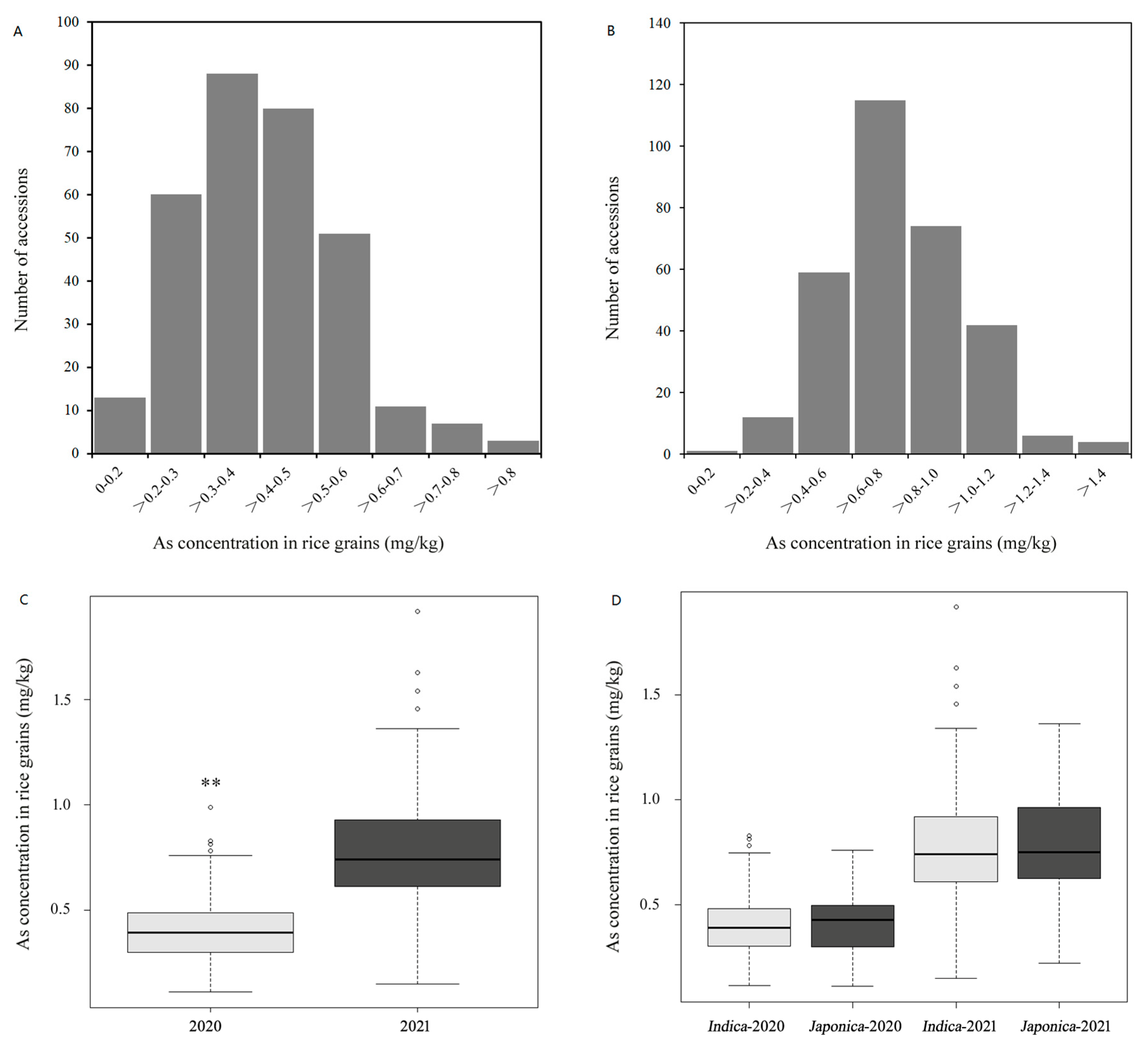
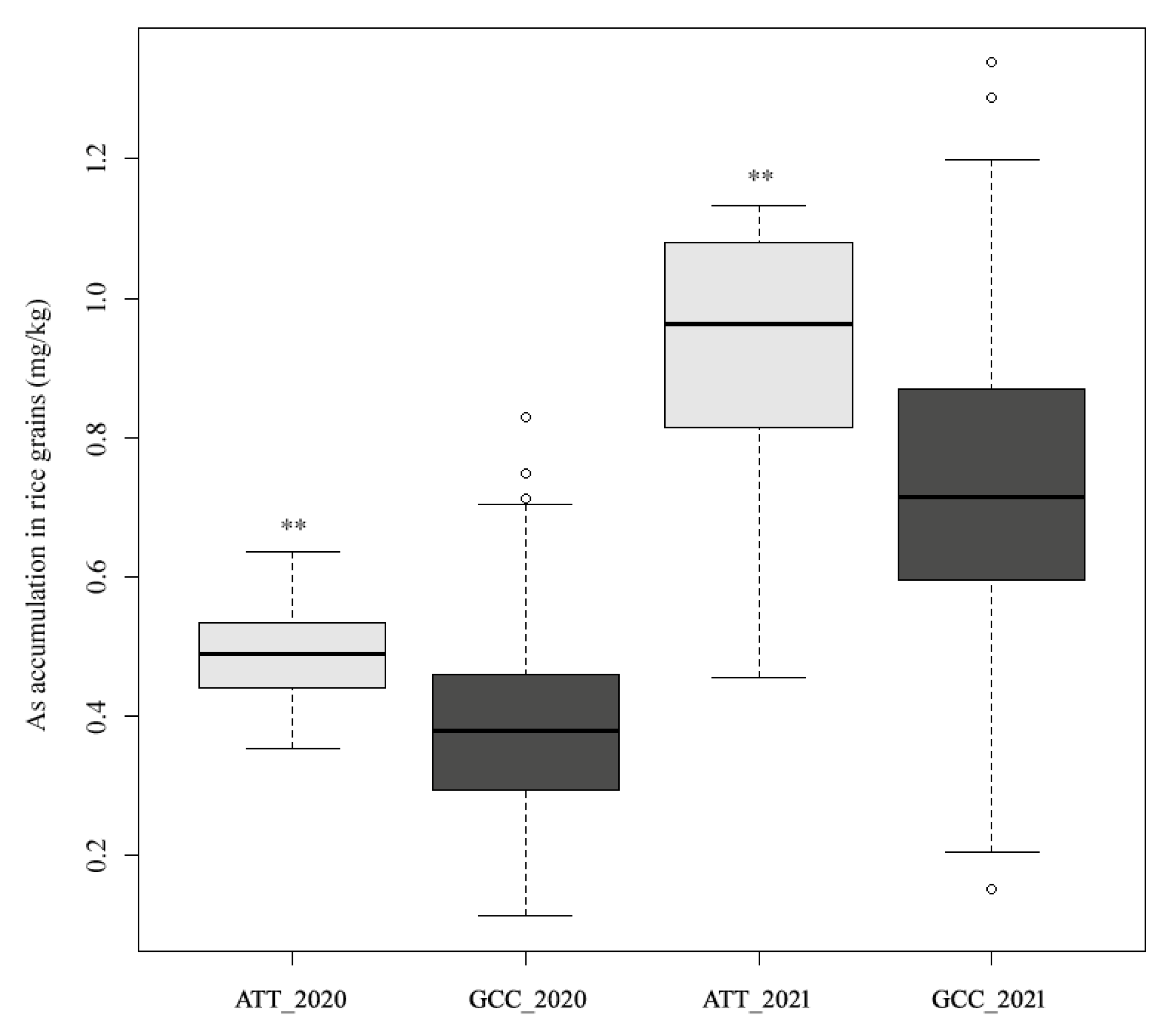
3.1. Variation in As Accumulation in Polished Rice
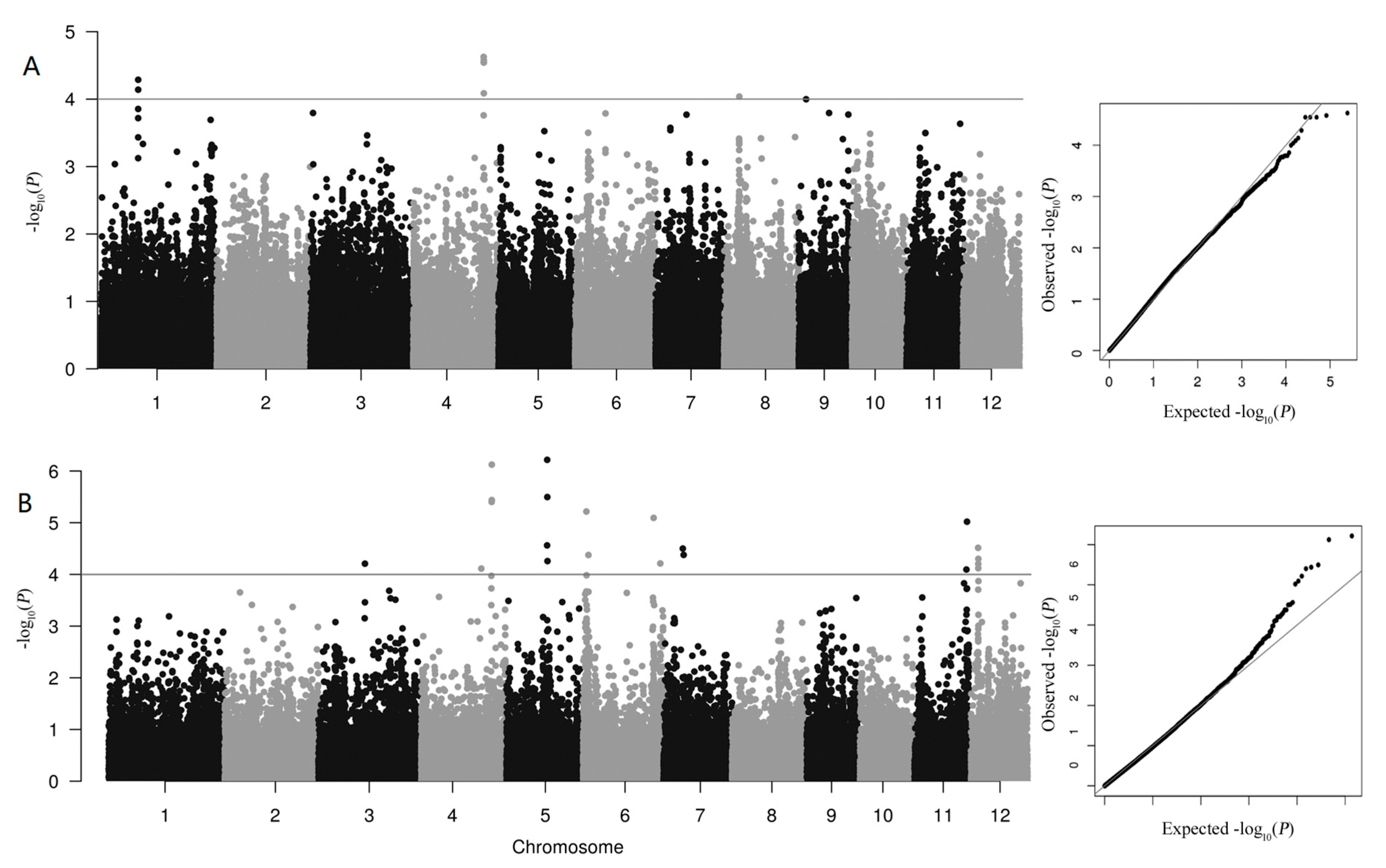
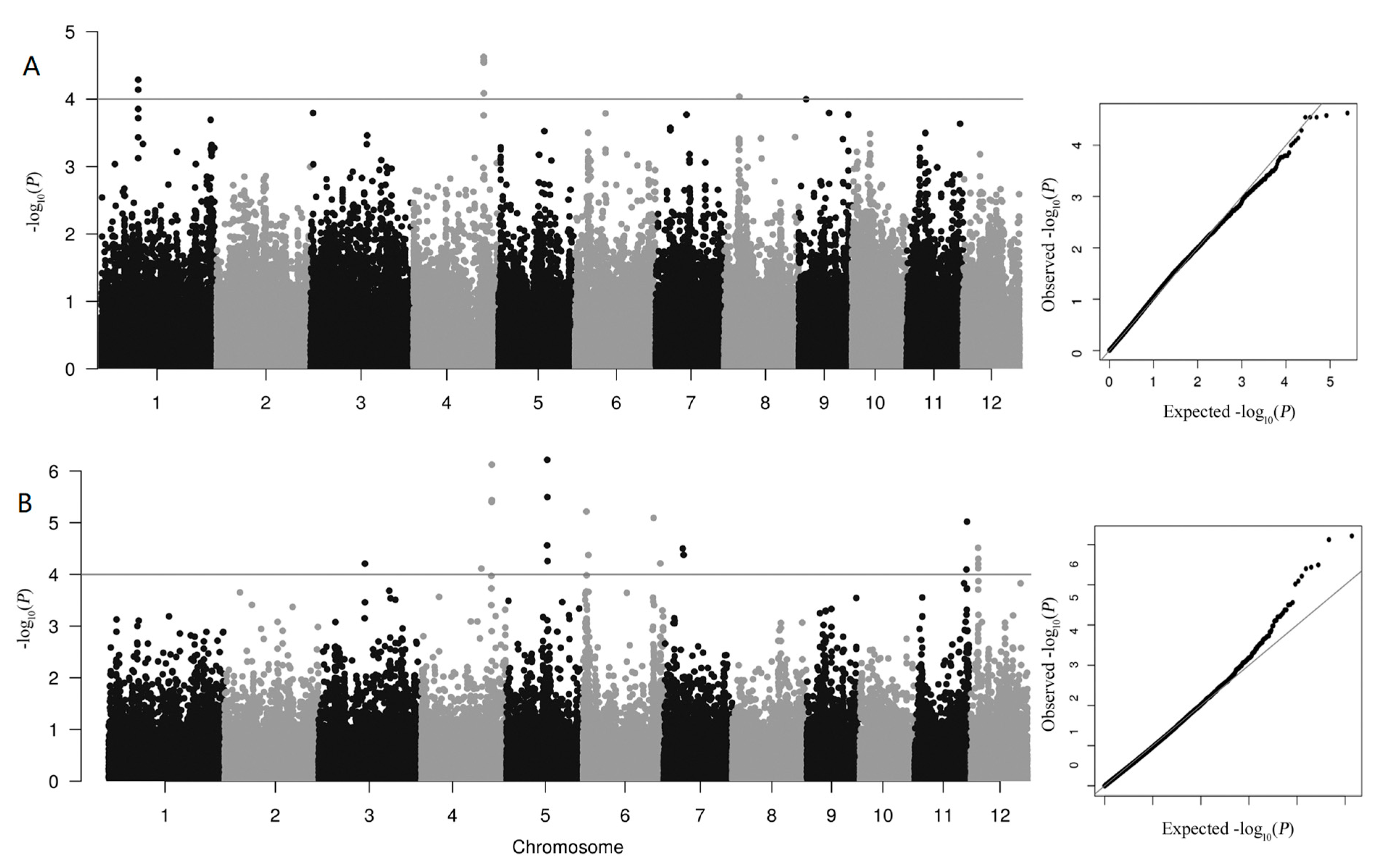
3.2. GWAS Analysis of QTLs Associated with As Accumulation
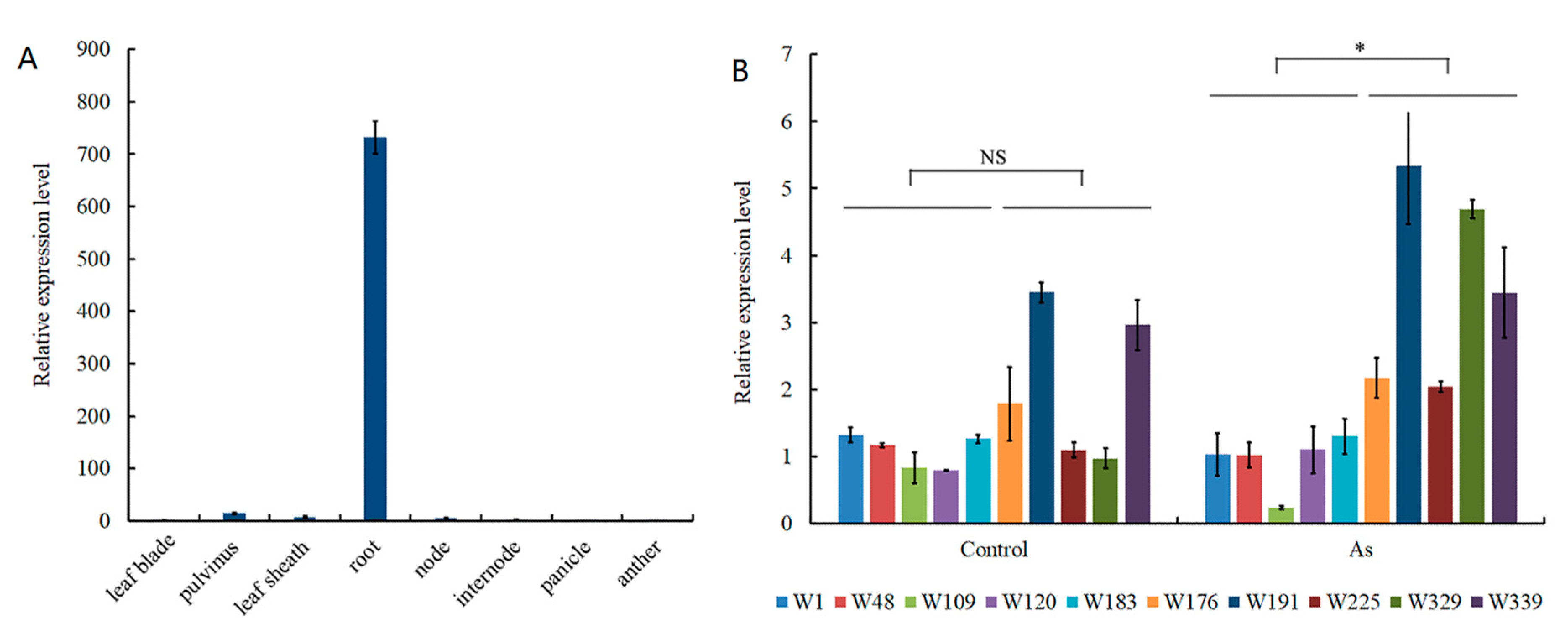
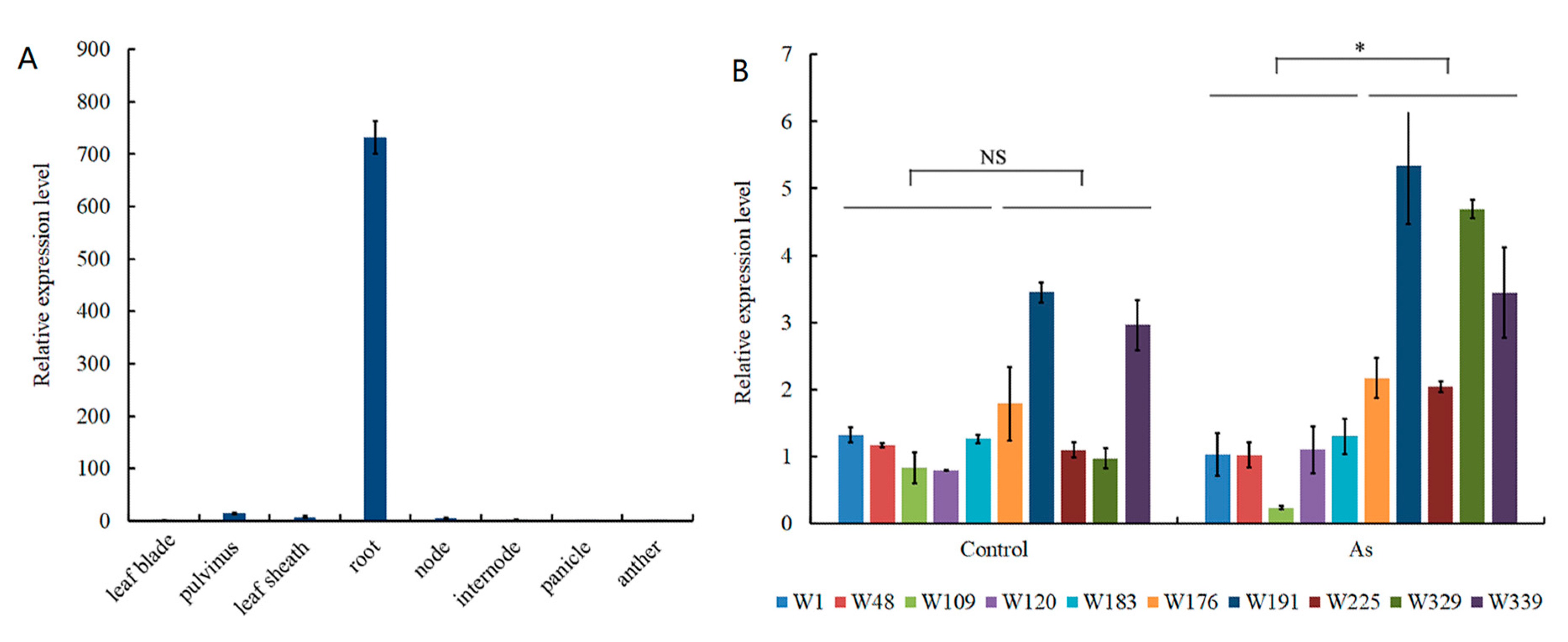
3.3. Candidate Gene Analysis for As Accumulation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, Z.; Zhang, D.; Xiong, X.; Yan, B.; Xie, W.; Sheen, J.; Li, J.F. A potent Cas9-derived gene activator for plant and mammalian cells. Nat. Plants 2017, 3, 930–936. [Google Scholar] [CrossRef]
- Smith, A.H.; Lopipero, P.A.; Bates, M.N.; Steinmaus, C.M. Public health. Arsenic epidemiology and drinking water standards. Science 2002, 296, 2145–2146. [Google Scholar] [CrossRef] [PubMed]
- Mawia, A.M.; Hui, S.; Zhou, L.; Li, H.; Tabassum, J.; Lai, C.; Wang, J.; Shao, G.; Wei, X.; Tang, S.; et al. Inorganic arsenic toxicity and alleviation strategies in rice. J. Hazard. Mater. 2021, 408, 124751. [Google Scholar] [CrossRef] [PubMed]
- Kandhol, N.; Singh, V.P.; Herrera-Estrella, L.; Tran, L.P.; Tripathi, D.K. Arsenite: The umpire of arsenate perception and responses in plants. Trends Plant Sci. 2022, 27, 420–422. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.G.; Sun, G.X.; Lei, M.; Teng, M.; Liu, Y.X.; Chen, N.C.; Wang, L.H.; Carey, A.M.; Deacon, C.; Raab, A.; et al. High percentage inorganic arsenic content of mining impacted and nonimpacted Chinese rice. Environ. Sci. Technol. 2008, 42, 5008–5013. [Google Scholar] [CrossRef]
- Williams, P.N.; Islam, M.R.; Adomako, E.E.; Raab, A.; Hossain, S.A.; Zhu, Y.G.; Feldmann, J.; Meharg, A.A. Increase in rice grain arsenic for regions of Bangladesh irrigating paddies with elevated arsenic in groundwaters. Environ. Sci. Technol. 2006, 40, 4903–4908. [Google Scholar] [CrossRef]
- Williams, P.N.; Raab, A.; Feldmann, J.; Meharg, A.A. Market basket survey shows elevated levels of As in South Central U.S. processed rice compared to California: Consequences for human dietary exposure. Environ. Sci. Technol. 2007, 41, 2178–2183. [Google Scholar] [CrossRef]
- Zhang, X.; Zhong, T.; Chen, D.; Cheng, M.; Liu, L.; Zhang, X.; Li, X. Assessment of arsenic (As) occurrence in arable soil and its related health risk in China. Environ. Geochem. Health 2016, 38, 691–702. [Google Scholar] [CrossRef]
- Zhou, Y.; Niu, L.; Liu, K.; Yin, S.; Liu, W. Arsenic in agricultural soils across China: Distribution pattern, accumulation trend, influencing factors, and risk assessment. Sci. Total Environ. 2018, 616–617, 156–163. [Google Scholar] [CrossRef]
- Xu, X.Y.; McGrath, S.P.; Meharg, A.A.; Zhao, F.J. Growing rice aerobically markedly decreases arsenic accumulation. Environ. Sci. Technol. 2008, 42, 5574–5579. [Google Scholar] [CrossRef]
- Su, Y.H.; Mcgrath, S.P.; Zhao, F.J. Rice is more efficient in arsenite uptake and translocation than wheat and barley. Plant Soil 2009, 328, 27–34. [Google Scholar] [CrossRef]
- Norton, G.J.; Adomako, E.E.; Deacon, C.M.; Carey, A.M.; Price, A.H.; Meharg, A.A. Effect of organic matter amendment, arsenic amendment and water management regime on rice grain arsenic species. Environ. Pollut. 2013, 177, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.P.; Singh, S.; Kumar, J.; Prasad, S.M. Investigating the roles of ascorbate-glutathione cycle and thiol metabolism in arsenate tolerance in ridged Luffa seedlings. Protoplasma 2015, 252, 1217–1229. [Google Scholar] [CrossRef] [PubMed]
- Muehe, E.M.; Wang, T.; Kerl, C.F.; Planer-Friedrich, B.; Fendorf, S. Rice production threatened by coupled stresses of climate and soil arsenic. Nat. Commun. 2019, 10, 4985. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, P.K.; Kim, K. A review of the arsenic concentration in paddy rice from the perspective of geoscience. Geosci. J. 2013, 17, 107–122. [Google Scholar] [CrossRef]
- Jaishankar, M.; Tseten, T.; Anbalagan, N.; Mathew, B.B.; Beeregowda, K.N. Toxicity, mechanism and health effects of some heavy metals. Interdiscip. Toxicol. 2014, 7, 60–72. [Google Scholar] [CrossRef]
- Khan, I.; Awan, S.A.; Rizwan, M.; Ali, S.; Zhang, X.; Huang, L. Arsenic behavior in soil-plant system and its detoxification mechanisms in plants: A review. Environ. Pollut. 2021, 286, 117389. [Google Scholar] [CrossRef]
- Zvobgo, G.; LwalabaWaLwalaba, J.; Sagonda, T.; Mutemachani Mapodzeke, J.; Muhammad, N.; Haider Shamsi, I.; Zhang, G. Phosphate alleviates arsenate toxicity by altering expression of phosphate transporters in the tolerant barley genotypes. Ecotoxicol. Environ. Saf. 2018, 147, 832–839. [Google Scholar] [CrossRef]
- Chen, J.; Liu, Y.; Ni, J.; Wang, Y.; Bai, Y.; Shi, J.; Gan, J.; Wu, Z.; Wu, P. OsPHF1 regulates the plasma membrane localization of low- and high-affinity inorganic phosphate transporters and determines inorganic phosphate uptake and translocation in rice. Plant Physiol. 2011, 157, 269–278. [Google Scholar] [CrossRef]
- Wu, Z.; Ren, H.; McGrath, S.P.; Wu, P.; Zhao, F.J. Investigating the contribution of the phosphate transport pathway to arsenic accumulation in rice. Plant Physiol. 2011, 157, 498–508. [Google Scholar] [CrossRef]
- Kamiya, T.; Islam, R.; Duan, G.; Uraguchi, S.; Fujiwara, T. Phosphate deficiency signaling pathway is a target of arsenate and phosphate transporter OsPT1 is involved in as accumulation in shoots of rice. Soil Sci. Plant Nutr. 2013, 59, 580–590. [Google Scholar] [CrossRef]
- Wang, P.; Zhang, W.; Mao, C.; Xu, G.; Zhao, F.J. The role of OsPT8 in arsenate uptake and varietal difference in arsenate tolerance in rice. J. Exp. Bot. 2016, 67, 6051–6059. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Li, P.; Xu, T.; Zeng, L.; Cheng, D.; Yang, M.; Luo, J.; Lian, X. OsPT4 contributes to arsenate uptake and transport in rice. Front. Plant Sci. 2017, 8, 2197. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.; Wang, T.; Chen, Z.; Tang, Z.; Wu, Z.; Salt, D.E.; Chao, D.Y.; Zhao, F.J. OsHAC1;1 and OsHAC1;2 function as arsenate reductases and regulate arsenic accumulation. Plant Physiol. 2016, 172, 1708–1719. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Shi, S.; Wang, L.; Tang, Z.; Lv, T.; Zhu, X.; Ding, X.; Wang, Y.; Zhao, F.J.; Wu, Z. OsHAC4 is critical for arsenate tolerance and regulates arsenic accumulation in rice. New Phytol. 2017, 215, 1090–1101. [Google Scholar] [CrossRef] [PubMed]
- Angulo-Bejarano, P.I.; Puente-Rivera, J.; Cruz-Ortega, R. Metal and metalloid toxicity in plants: An overview on molecular aspects. Plants 2021, 10, 635. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.F.; Yamaji, N.; Mitani, N.; Xu, X.Y.; Su, Y.H.; McGrath, S.P.; Zhao, F.J. Transporters of arsenite in rice and their role in arsenic accumulation in rice grain. Proc. Natl. Acad. Sci. USA 2008, 105, 9931–9935. [Google Scholar] [CrossRef]
- Ma, J.F.; Yamaji, N.; Mitani, N.; Tamai, K.; Konishi, S.; Fujiwara, T.; Katsuhara, M.; Yano, M. An efflux transporter of silicon in rice. Nature 2007, 448, 209–212. [Google Scholar] [CrossRef]
- Sharma, S.S.; Kumar, V.; Dietz, K.J. Emerging Trends in Metalloid-Dependent Signaling in Plants. Trends Plant Sci. 2021, 26, 452–471. [Google Scholar] [CrossRef]
- Zhang, J.; Hamza, A.; Xie, Z.; Hussain, S.; Brestic, M.; Tahir, M.A.; Ulhassan, Z.; Yu, M.; Allakhverdiev, S.I.; Shabala, S. Arsenic transport and interaction with plant metabolism: Clues for improving agricultural productivity and food safety. Environ. Pollut. 2021, 290, 117987. [Google Scholar] [CrossRef]
- Dasgupta, T.; Hossain, S.A.; Meharg, A.A.; Price, A.H. An arsenate tolerance gene on chromosome 6 of rice. New Phytol. 2004, 163, 45–49. [Google Scholar] [CrossRef]
- Syed, M.A.; Iftekharuddaula, K.M.; Mian, M.A.K.; Rasul, M.G.; Rahmam, G.K.M.M.; Panaullah, G.M.; Lauren, J.G.; Duxbury, J.M.; Biswas, P.S. Main effect QTLs associated with arsenic phyto-toxicity tolerance at seedling stage in rice (Oryza sativa L.). Euphytica 2016, 209, 805–814. [Google Scholar] [CrossRef]
- Zhang, J.; Zhu, Y.G.; Zeng, D.L.; Cheng, W.D.; Qian, Q.; Duan, G.L. Mapping quantitative trait loci associated with arsenic accumulation in rice (Oryza sativa). New Phytol. 2008, 177, 350–356. [Google Scholar] [CrossRef] [PubMed]
- Norton, G.J.; Deacon, C.M.; Xiong, L.; Huang, S.; Meharg, A.A.; Price, A.H. Genetic mapping of the rice ionome in leaves and grain: Identification of QTLs for 17 elements including arsenic, cadmium, iron and selenium. Plant Soil 2010, 329, 139–153. [Google Scholar] [CrossRef]
- Norton, G.J.; Duan, G.L.; Lei, M.; Zhu, Y.G.; Meharg, A.A.; Price, A.H. Identification of quantitative trait loci for rice grain element composition on an arsenic impacted soil: Influence of flowering time on genetic loci. Ann. Appl. Biol. 2012, 161, 46–56. [Google Scholar] [CrossRef]
- Kuramata, M.; Abe, T.; Kawasaki, A.; Ebana, K.; Shibaya, T.; Yano, M.; Ishikawa, S. Genetic diversity of arsenic accumulation in rice and QTL analysis of methylated arsenic in rice grains. Rice 2013, 6, 3. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Pinson, S.R.; Tarpley, L.; Huang, X.Y.; Lahner, B.; Yakubova, E.; Baxter, I.; Guerinot, M.L.; Salt, D.E. Mapping and validation of quantitative trait loci associated with concentrations of 16 elements in unmilled rice grain. Theor. Appl. Genet. 2014, 127, 137–165. [Google Scholar] [CrossRef]
- Sun, X.; Liu, D.; Zhang, X.; Li, W.; Liu, H.; Hong, W.; Jiang, C.; Guan, N.; Ma, C.; Zeng, H.; et al. SLAF-seq: An efficient method of large-scale de novo SNP discovery and genotyping using high-throughput sequencing. PLoS ONE 2013, 8, e58700. [Google Scholar] [CrossRef]
- GB 5009. 268-2016; Chinese National Standard: Determination of Multielement in Food. National Health and Family Planning Commission of China: Beijing, China, 2016.
- Pan, X.; Li, Y.; Liu, W.; Liu, S.; Min, J.; Xiong, H.; Dong, Z.; Duan, Y.; Yu, Y.; Li, X. QTL mapping and candidate gene analysis of cadmium accumulation in polished rice by genome-wide association study. Sci. Rep. 2020, 10, 11791. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; 1000 Genome Project Data Processing Subgroup. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.C.; Chow, C.C.; Tellier, L.C.; Vattikuti, S.; Purcell, S.M.; Lee, J.J. Second-generation PLINK: Rising to the challenge of larger and richer datasets. GigaScience 2015, 4, 7. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Stephens, M. Genome-wide efficient mixed-model analysis for association studies. Nat. Genet. 2021, 44, 821–824. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.; Guo, Z.; Li, X.; Ye, H.; Li, X.; Xiong, L. New insights into the genetic basis of natural chilling and cold shock tolerance in rice by genome-wide association analysis. Plant Cell Environ. 2016, 39, 556–570. [Google Scholar] [CrossRef] [PubMed]
- GB 2762-2022; Chinese National Standard: Limits of Contaminants in Food. National Health Commission of China: Beijing, China, 2022.
- Huang, T.L.; Nguyen, Q.T.; Fu, S.F.; Lin, C.Y.; Chen, Y.C.; Huang, H.J. Transcriptomic changes and signalling pathways induced by arsenic stress in rice roots. Plant Mol. Biol. 2012, 80, 587–608. [Google Scholar] [CrossRef]
- Chen, Y.; Han, Y.H.; Cao, Y.; Zhu, Y.G.; Rathinasabapathi, B.; Ma, L.Q. Arsenic transport in rice and biological solutions to reduce arsenic risk from rice. Front. Plant Sci. 2017, 8, 268. [Google Scholar] [CrossRef]
- Norton, G.J.; Douglas, A.; Lahner, B.; Yakubova, E.; Guerinot, M.L.; Pinson, S.R.; Tarpley, L.; Eizenga, G.C.; McGrath, S.P.; Zhao, F.J.; et al. Genome wide association mapping of grain arsenic, copper, molybdenum and zinc in rice (Oryza sativa L.) grown at four international field sites. PLoS ONE 2014, 9, e89685. [Google Scholar] [CrossRef]
- Huang, X.; Wei, X.; Sang, T.; Zhao, Q.; Feng, Q.; Zhao, Y.; Li, C.; Zhu, C.; Lu, T.; Zhang, Z.; et al. Genome-wide association studies of 14 agronomic traits in rice landraces. Nat. Genet. 2010, 42, 961–967. [Google Scholar] [CrossRef]
- Korte, A.; Farlow, A. The advantages and limitations of trait analysis with GWAS: A review. Plant Methods 2013, 9, 29. [Google Scholar] [CrossRef]
- Yu, J.; Pressoir, G.; Briggs, W.H.; Vroh Bi, I.; Yamasaki, M.; Doebley, J.F.; McMullen, M.D.; Gaut, B.S.; Nielsen, D.M.; Holland, J.B.; et al. A unified mixed-model method for association mapping that accounts for multiple levels of relatedness. Nat. Genet. 2006, 38, 203–208. [Google Scholar] [CrossRef]
- Famoso, A.N.; Zhao, K.; Clark, R.T.; Tung, C.W.; Wright, M.H.; Bustamante, C.; Kochian, L.V.; McCouch, S.R. Genetic architecture of aluminum tolerance in rice (Oryza sativa) determined through genome-wide association analysis and QTL mapping. PLoS Genet. 2011, 7, e1002221. [Google Scholar] [CrossRef] [PubMed]
- Rubio, V.; Linhares, F.; Solano, R.; Martín, A.C.; Iglesias, J.; Leyva, A.; Paz-Ares, J. A conserved MYB transcription factor involved in phosphate starvation signaling both in vascular plants and in unicellular algae. Genes Dev. 2001, 15, 2122–2133. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Xu, Y.; Ma, Q.; Xu, W.; Wang, T.; Xue, Y.; Chong, K. Overexpression of an R1R2R3 MYB gene, OsMYB3R-2, increases tolerance to freezing, drought, and salt stress in transgenic Arabidopsis. Plant Physiol. 2007, 143, 1739–1751. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.Z.; Chen, M.X.; Yu, L.J.; Xie, L.J.; Yuan, L.B.; Qi, H.; Xiao, M.; Guo, W.; Chen, Z.; Yi, K.; et al. OsARM1, an R2R3 MYB transcription factor, is involved in regulation of the response to arsenic stress in rice. Front. Plant Sci. 2017, 8, 1868. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, H.Y.; Chen, Y.F. The transcription factor MYB40 is a central regulator in arsenic resistance in Arabidopsis. Plant Commun. 2021, 2, 100234. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Field ID | As-2020 | As-2021 | Accession Name | Type |
|---|---|---|---|---|
| W165 | 0.23 ± 0.0127 | 0.26 ± 0.0134 * | Baimidongzhan | Landrace |
| W176 | 0.13 ± 0.0106 * | 0.15 ± 0.0374 * | Chenwan-3 | Bred variety |
| W191 | 0.25 ± 0.0233 | 0.22 ± 0.0537 * | Feitianzao | Landrace |
| W329 | 0.12 ± 0.0092 * | 0.20 ± 0.0276 * | Aizizhan | Landrace |
| W348 | 0.27 ± 0.0240 | 0.30 ± 0.0113 | Hong-410 | Landrace |
| W351 | 0.29 ± 0.0170 | 0.29 ± 0.0078 * | Jiayu-948 | Bred variety |
| QTLs | Chr. | Position of Lead SNP | allele0 | allele1 | p-Value |
|---|---|---|---|---|---|
| 2020 | |||||
| qAs1 | 1 | 15248892 | G | A | 5.16 × 10−5 |
| qAs4 | 4 | 28589826 | G | A | 2.36 × 10−5 |
| 2021 | |||||
| qAs4 | 4 | 28644370 | C | T | 7.52 × 10−5 |
| qAs5 | 5 | 16538553 | C | T | 6.08 × 10−7 |
| qAs12 | 12 | 2908161 | A | G | 3.06 × 10−5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, Z.; Guo, L.; Li, X.; Li, Y.; Liu, W.; Chen, Z.; Liu, L.; Liu, Z.; Guo, Y.; Pan, X. Genome-Wide Association Study of Arsenic Accumulation in Polished Rice. Genes 2023, 14, 2186. https://doi.org/10.3390/genes14122186
Dong Z, Guo L, Li X, Li Y, Liu W, Chen Z, Liu L, Liu Z, Guo Y, Pan X. Genome-Wide Association Study of Arsenic Accumulation in Polished Rice. Genes. 2023; 14(12):2186. https://doi.org/10.3390/genes14122186
Chicago/Turabian StyleDong, Zheng, Liang Guo, Xiaoxiang Li, Yongchao Li, Wenqiang Liu, Zuwu Chen, Licheng Liu, Zhixi Liu, Yujing Guo, and Xiaowu Pan. 2023. "Genome-Wide Association Study of Arsenic Accumulation in Polished Rice" Genes 14, no. 12: 2186. https://doi.org/10.3390/genes14122186
APA StyleDong, Z., Guo, L., Li, X., Li, Y., Liu, W., Chen, Z., Liu, L., Liu, Z., Guo, Y., & Pan, X. (2023). Genome-Wide Association Study of Arsenic Accumulation in Polished Rice. Genes, 14(12), 2186. https://doi.org/10.3390/genes14122186