
Growth Axis Somatostatin, Growth Hormone Receptor, and Insulin-like Growth Factor-1 Genes Express and Are Affected by the Injection of Exogenous Growth Hormone in Chinemys reevesii

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animal
2.2. Sample Preparation
2.3. Tissue Collection and Isolation of RNA
2.4. Cloning and DNA Sequencing of the SS Gene of the Reeves’ Turtle
2.5. RT-PCR
2.6. Statistical Analysis
3. Results
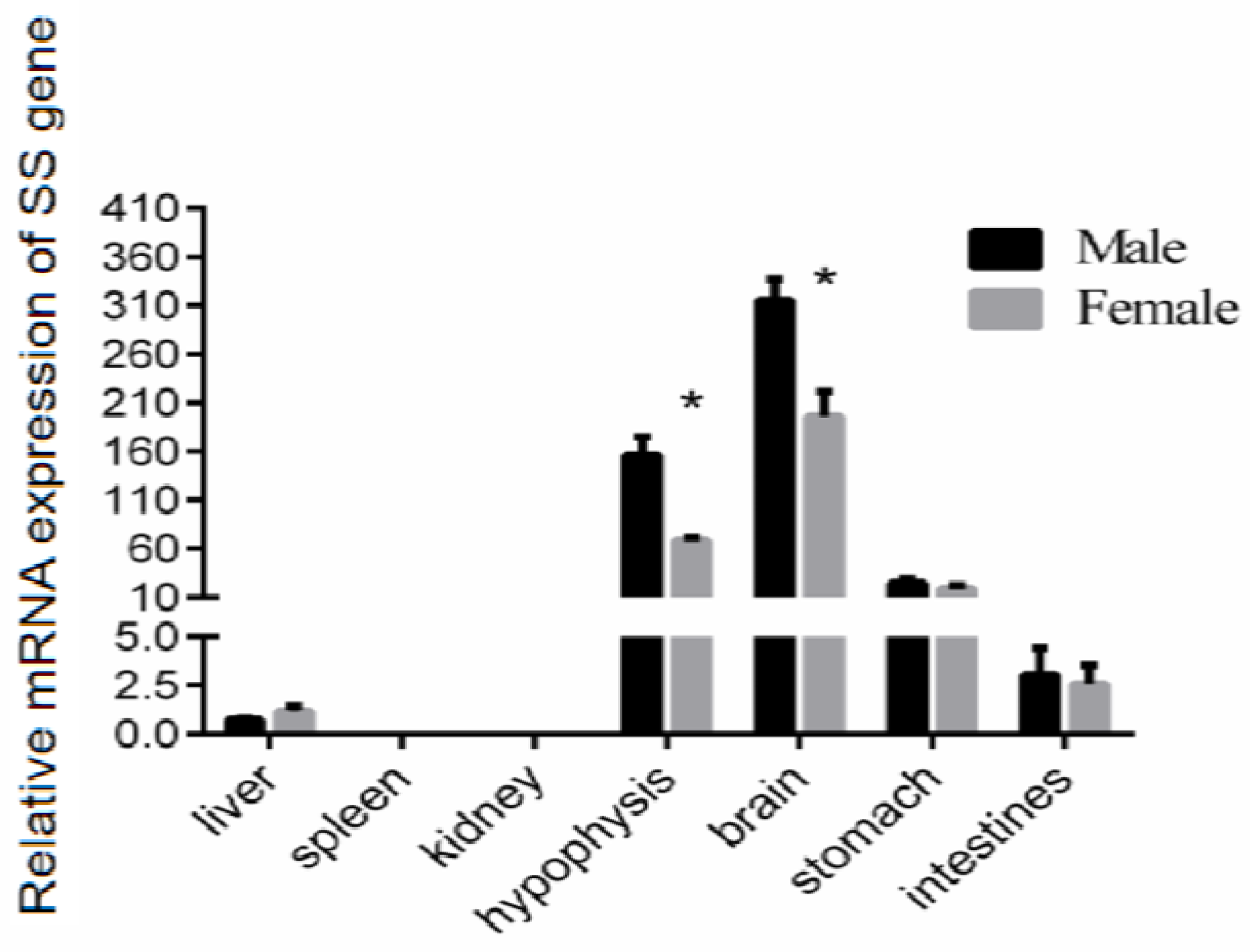
3.1. Tissue Distribution of SS Gene Expression in the Reeves’ Turtle
3.2. Modulation of SS Gene Expression in the Brain under Periodic Injection of Growth Hormone
3.3. Effect of Exogenous Growth Hormone on SS Gene Expression in Tissues of the Reeves’ Turtle
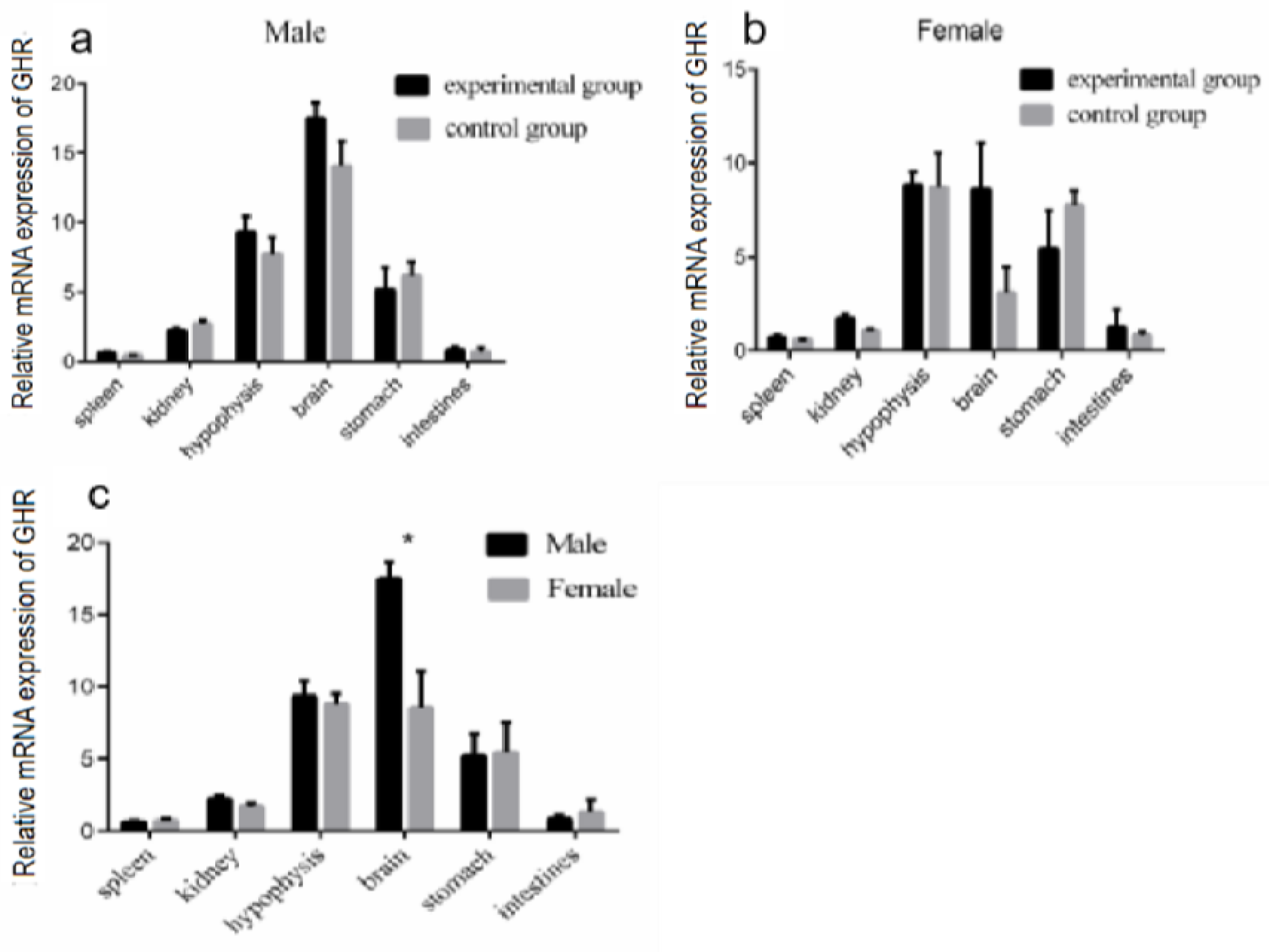
3.4. Modulation of GHR Gene Expression in the Liver under Periodic Injection of Growth Hormone
3.5. Modulation of GHR mRNA Expression in Different Tissues of Reeves’ Turtles after Four Weeks of GH Injection
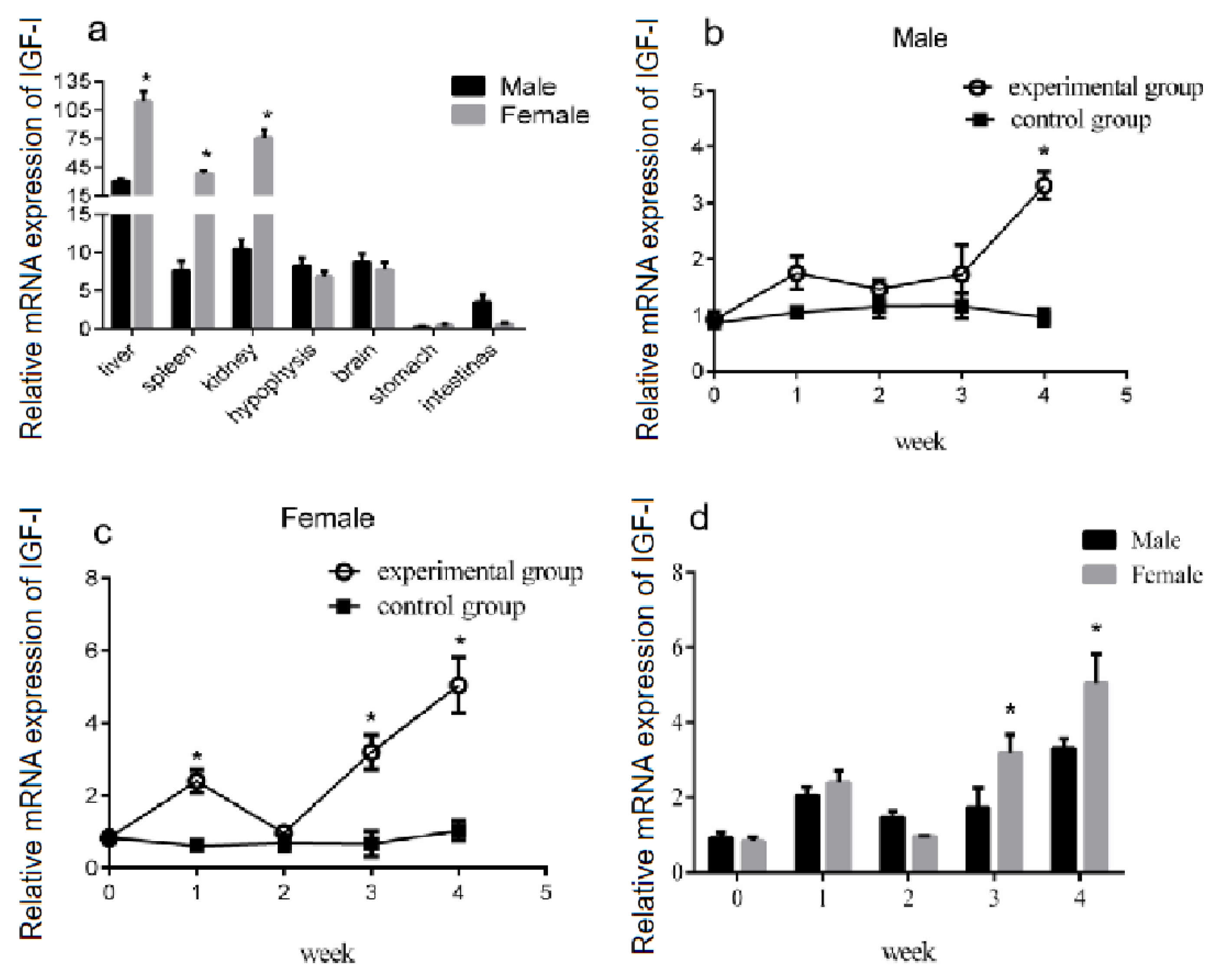
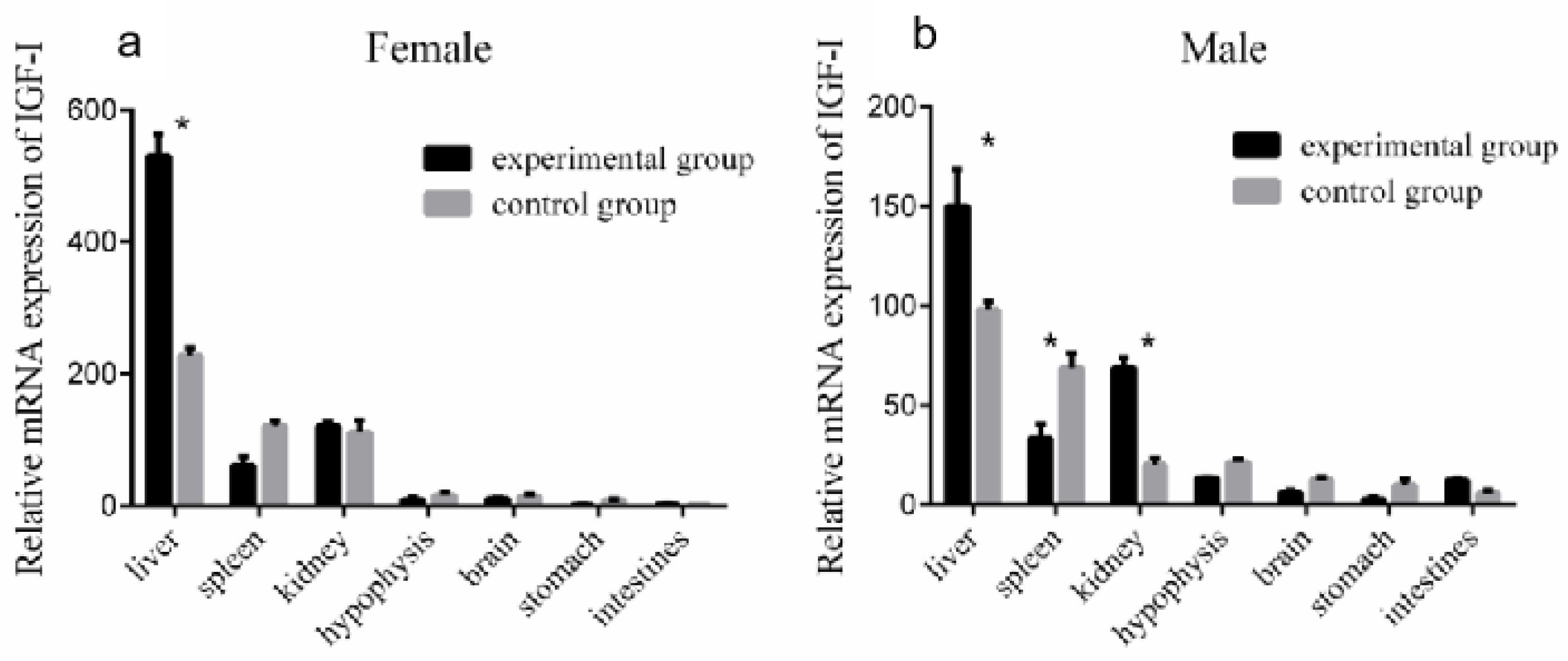
3.6. Effect of Periodic Exogenous GH Injection on IGF-I Gene Expression in the Liver
3.7. Changes in IGF-I Gene mRNA Expression in the Liver after Periodic Injection of Growth Hormone
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Omeyer, L.C.M.; Fuller, W.J.; Godley, B.J.; Snape, R.T.E. The effect of biologging systems on reproduction, growth and survival of adult sea turtles. Mov. Ecol. 2019, 7, 2. [Google Scholar] [CrossRef] [PubMed]
- Congdon, J.D.; Nagle, R.D.; Kinney, O.M.; van Loben Sels, R.C.; Quinter, T.; Tinkle, D.W. Testing hypotheses of aging in long-lived painted turtles (Chrysemys picta). Exp. Gerontol. 2003, 38, 765–772. [Google Scholar] [CrossRef] [PubMed]
- Congdon, J.D.; Nagle, R.D.; Kinney, O.M.; van Loben Sels, R.C. Hypotheses of aging in a long-lived vertebrate, Blanding’s turtle (Emydoidea blandingii). Exp. Gerontol. 2001, 36, 813–827. [Google Scholar] [CrossRef] [PubMed]
- Omeyer, L.C.M.; Fuller, W.J.; Godley, B.J.; Snape, R.T.E. Determinate or indeterminate growth? Revisiting the growth strategy of sea turtles. Mar. Ecol. Prog. Ser. 2018, 596, 199–211. [Google Scholar] [CrossRef]
- Frýdlová, P.; Mrzílková, J.; Šeremeta, M.; Křemen, J. Determinate growth is predominant and likely ancestral in squamate reptiles. Proc. R. Soc. B Biolpgical Sci. 2021, 287, 20202737. [Google Scholar] [CrossRef]
- Frýdlová, P.; Mrzílková, J.; Šeremeta, M.; Křemen, J. Universality of indeterminate growth in lizards rejected: The micro-ct reveals contrasting timing of growth cartilage persistence in iguanas, agamas, and chameleons. Sci. Rep. 2019, 9, 18913. [Google Scholar] [CrossRef]
- Frýdlová, P.; Nutilová, V.; Dudák, J.; Žemlička, J.; Němec, P.; Velenský, P.; Jirásek, T. Patterns of growth in monitor lizards (Varanidae) as revealed by computed tomography of femoral growth plates. Zoomorphology 2017, 136, 95–106. [Google Scholar] [CrossRef]
- Zhu, W.; He, Y.; Ruan, Z.; Zhang, X.; Liao, L.; Gao, Y.; Lin, N.; Chen, X.; Liang, R.; Liu, W. Identification of the cDNA Encoding the Growth Hormone Receptor (GHR) and the Regulation of GHR and IGF-I Gene Expression by Nutritional Status in Reeves’ Turtle (Chinemys reevesii). Front. Genet. 2020, 11, 587. [Google Scholar] [CrossRef]
- Liu, W.-S.; Ma, J.-E.; Li, W.-X.; Zhang, J.-G.; Wang, J.; Nie, Q.-H.; Qiu, F.-F.; Fang, M.-X.; Zeng, F.; Wang, X.; et al. The Long Intron 1 of Growth Hormone Gene from Reeves’ Turtle (Chinemys reevesii) Correlates with Negatively Regulated GH Expression in Four Cell Lines. Int. J. Mol. Sci. 2016, 17, 543. [Google Scholar] [CrossRef]
- Very, N.M.; Kittilson, J.D.; Klein, S.E.; Sheridan, M.A. Somatostatin inhibits basal and growth hormone-stimulated hepatic insulin-like growth factor-I production. Mol. Cell. Endocrinol. 2008, 281, 19–26. [Google Scholar] [CrossRef]
- Liu, W.S.; Li, Y. Studies on the biochemical indices of blood in Chinemys reevesii. Acta Hydrobiol. Sin. 2006, 30, 499–501. [Google Scholar]
- Liu, W.S.; Li, Y. Studies on the micro-structure of hypophysis gland and ultrastructure of adenohypophysis in Chinemys reevesii. Acta Hydrobiol. Sin. 2005, 29, 661–666. [Google Scholar]
- Krulich, L.; Dhariwal, A.P.S.; Mccann, S.M. Stimulatory and inhibitory effects of purified hypothalamic extracts on growth hormone release from rat pituitary in vitro. Endocrinology 1968, 83, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Brazeau, P.; Vale, W.; Burgus, R.; Ling, N.; Butcher, M.; Rivier, J.; Guillemin, R. Hypothalamic Polypeptide That Inhibits the Secretion of Immunoreactive Hypophysis Growth Hormone. Science 1973, 179, 77–79. [Google Scholar] [CrossRef] [PubMed]
- Pradayrol, L.; Chayvialle, J.A.; Carlquist, M.; Mutt, V. Isolation of a porcine intestinal peptide with C-terminal somatostatin. Biochem. Biophys. Res. Commun. 1978, 85, 701–708. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.-W.; Otto, C.J.; Peter, R.E. Evolution of neuroendocrine peptide systems: Hormone and somatostatin. Comp. Biochem. Physiol. C Pharmacol. Toxicol. Endocrinol. 1998, 119, 375–388. [Google Scholar] [CrossRef]
- Barnett, P. Somatostatin and somatostatin receptor physiology. Endocrine 2003, 20, 25–64. [Google Scholar] [CrossRef]
- Brockway, D.F.; Griffith, K.R.; Aloimonos, C.M.; Clarity, T.T.; Moyer, J.B.; Smith, G.C.; Dao, N.C.; Hossain, M.S.; Drew, P.J.; Gordon, J.A.; et al. Somatostatin peptide signaling dampens cortical circuits and promotes exploratory behavior. Cell Rep. 2023, 42, 112976. [Google Scholar] [CrossRef]
- Nagaeva, E.; Schäfer, A.; Linden, A.M.; Elsilä, L.V.; Egorova, K.; Umemori, J.; Ryazantseva, M.; Korpi, E.R. Somatostatin-Expressing Neurons in the Ventral Tegmental Area Innervate Specific Forebrain Regions and Are Involved in Stress Response. eNeuro 2023, 10, ENEURO.0149-23.2023. [Google Scholar] [CrossRef]
- Tang, M.; Liu, C.; Li, R.; Lin, H.; Peng, Y.; Lang, Y.; Su, K.; Xie, Z.; Li, M.; Yang, X.; et al. Spatial and temporal expression pattern of somatostatin receptor 2 in mouse. Chin. J. Biotechnol. 2023, 39, 2656–2668. [Google Scholar] [CrossRef]
- Peluso, G.; Petillo, O.; Melone, M.A.; Mazzarella, G.; Ranieri, M.; Tajana, G.F. Modulation of cytokine production in activated human monocytes by somatostatin. Neuropeptides 1996, 30, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Elliott, D.E.; Li, J.; Blum, A.M.; Metwali, A.; Patel, Y.C.; Weinstock, J.V. SSTR2A is the dominant somatostatin receptor subtype expressed by inflammatory cells, is widely expressed and directly regulates T cell IFN-γ release. Eur. J. Immunol. 1999, 29, 2454–2463. [Google Scholar] [CrossRef]
- Herszényi, L.; Mihály, E.; Tulassay, Z. Somatostatin and gastrointestinal tract. Clinical experiences. Orvosi Hetil. 2013, 154, 1535–1540. [Google Scholar] [CrossRef]
- Kappeler, L.; Zizzari, P.; Grouselle, D.; Epelbaum, J.; Bluet-Pajot, M.T. Plasma and Hypothalamic Peptide-Hormone Levels Regulating Somatotroph Function and Energy Balance in Fed and Fasted States: A Comparative Study in Four Strains of Rats. J. Neuroendocrinol. 2004, 16, 980–988. [Google Scholar] [CrossRef]
- Ben-Shlomo, A.; Melmed, S. Pituitary somatostatin receptor signaling. Trends Endocrinol. Metab. 2010, 21, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Froidevaux, S.; Eberle, A.N. Somatostatin analogs and radiopeptides in cancer therapy. Biopolymers 2002, 66, 161–183. [Google Scholar] [CrossRef] [PubMed]
- Strosberg, J.; Kvols, L. Antiproliferative effect of somatostatin analogs in gastroenteropancreatic neuroendocrine tumors. World J. Gastroenterol. 2010, 16, 2963–2970. [Google Scholar] [CrossRef]
- Gadelha, M.R.; Bronstein, M.D.; Brue, T.; Coculescu, M.; Fleseriu, M.; Guitelman, M.; Pronin, V.; Raverot, G.; Shimon, I.; Lievre, K.K.; et al. Pasireotide versus continued treatment with octreotide or lanreotide in patients with inadequately controlled acromegaly (PAOLA): A randomised, phase 3 trial. Lancet Diabetes Endocrinol. 2014, 2, 875–884. [Google Scholar] [CrossRef]
- McLean, E.; Donaldson, E.M. The Role of Growth Hormone in the Growth of Poikilotherms. Endocrinol. Growth Dev. Metab. Vertebr. 1993, 248, 43–71. [Google Scholar]
- Reindl, K.; Sheridan, M.A. Peripheral regulation of the growth hormone-insulin-like growth factor system in fish and other vertebrates. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2012, 163, 231–245. [Google Scholar] [CrossRef]
- Shafik, B.M.; Kamel, E.R.; Mamdouh, M.; Elrafaay, S.; Nassan, M.A.; El-Bahy, S.M.; El-Tarabany, M.S.; Manaa, E.A. Performance, Blood Lipid Profile, and the Expression of Growth Hormone Receptor (GHR) and Insulin-Like Growth Factor-1 (IGF-1) Genes in Purebred and Crossbred Quail Lines. Animals 2022, 12, 1245. [Google Scholar] [CrossRef] [PubMed]
- Pierce, A.L.; Fox, B.K.; Davis, L.K.; Visitacion, N. Prolactin receptor, growth hormone receptor, and putative somatolactin receptor in Mozambique tilapia: Tissue specific expression and differential regulation by salinity and fasting. Gen. Comp. Endocrinol. 2007, 154, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Sparkman, A.M.; Schwartz, T.S.; Schwartz, T.S.; Madden, J.A.; Boyken, S.E.; Ford, N.B.; Serb, J.M.; Bronikowski, A.M. Rates of molecular evolution vary in vertebrates for insulin-like growth factor-1 (IGF-1), a pleiotropic locus that regulates life history traits. Gen. Comp. Endocrinol. 2012, 178, 164–173. [Google Scholar] [CrossRef] [PubMed]
- Denley, A.; Cosgrove, L.J.; Booker, G.W.; Wallace, J.C.; Forbes, B.E. Molecular interactions of the IGF system. Cytokine Growth Factor Rev. 2005, 16, 421–439. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhu, X.; Chen, Y. Age and growth in reeves’ turtle Chinemys reebesii Gray. J. Dalian Fish. Univ. 2005, 20, 73–76. [Google Scholar] [CrossRef]
- Yang, Y.; Ren, Z.Y.; Su, C.B.; Wang, R.Z.; Ma, W.B. Diagnosis and treatment of pituitary growth hormone adenoma. Chin. J. Neurosurg. Dis. Res. 2004, 04, 254–255. [Google Scholar]
- Jiang, Q.; Lian, A.; He, Q. Dopamine inhibits somatolactin gene expression in tilapia pituitary cells through the dopamine D2 receptors. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2016, 197, 35–42. [Google Scholar] [CrossRef]
- Ye, X.; Li, W.S.; Lin, H.R. Progress of research on fish somatostatin and receptor. J. Fish. China 2007, 31, 264–272. [Google Scholar]
- Li, C.J.; Wei, Q.W.; Zhou, L.; Cao, H.; Zhang, Y.; Gui, J.F. Molecular and expression characterization of two somatostatin genes in the Chinese sturgeon, Acipenser sinensis. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2009, 154, 127–134. [Google Scholar] [CrossRef]
- Kamegai, J.; Unterman, T.G.; Frohman, L.A.; Kineman, R.D. Hypothalamic/hypophysis-axis of the spontaneous dwarf rat: Autofeedback regulation of growth hormone(GH) includes suppression of GH releasing-hormone receptor messenger ribonucleic acid. Endocrinology 1998, 139, 3554–3560. [Google Scholar] [CrossRef][Green Version]
- Hu, B.; Hu, S.; Yang, M.; Liao, Z.; Zhang, D.; Luo, Q.; Zhang, X.; Li, H. Growth Hormone Receptor Gene is Essential for Chicken Mitochondrial Function In Vivo and In Vitro. Int. J. Mol. Sci. 2019, 20, 1608. [Google Scholar] [CrossRef] [PubMed]
- Won, E.T.; Douros, J.D.; Hurt, D.A.; Borski, R.J. Leptin stimulates hepatic growth hormone receptor and insulin-like growth factor gene expression in a teleost fish, the hybrid striped bass. Gen. Comp. Endocrinol. 2016, 229, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Mertani, H.C.; Morel, G. In situ gene expression of growth hormone (GH) receptor and GH binding protein in adult male rat tissues. Mol. Cell. Endocrinol. 1995, 109, 47–61. [Google Scholar] [CrossRef]
- Burnside, J.; Liou, S.S.; Zhong, C.; Cogburn, L.A. Abnormal growth hormone receptor gene expression in the sex-linked dwarf chicken. Gen. Comp. Endocrinol. 1992, 88, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Ymer, S.I.; Herington, A.C. Developmental expression of the growth hormone receptor gene in rabbit tissues. Mol. Cell. Endocrinol. 1992, 83, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Yakar, S.; Setser, J.; Zhao, H.; Stannard, B.; Haluzik, M.; Glatt, V.; Bouxsein, M.L.; Kopchick, J.J.; LeRoith, D. Inhibition of growth hormone action improves insulin sensitivity in liver IGF-1–deficient mice. J. Clin. Investig. 2004, 113, 96–105. [Google Scholar] [CrossRef]
- Kim, I.; Jin, E.J.; Baik, K.; Park, C.H.; Kim, W.K.; Kang, C.W.; Ko, Y.; Jang, I.; Choi, W.S.; Lee, C.Y. Expression and secretion of the insulin-like growth factor system components by pig liver cells. Asian Australas. J. Anim. Sci. 2008, 21, 1244–1251. [Google Scholar] [CrossRef]
- de Brun, V.; Meikle, A.; Casal, A.; Carriquiry, M.; Menezes, C.; Forcada, F.; Sosa, C.; Abecia, J.A. Hepatic Expression of Insulin-Like Growth Factor-1 in Underfed Pregnant Ewes. J. Agric. Ence Technol. A 2016, 6, 355–362. [Google Scholar]
- Schmid, A.C.; Lutz, I.; Kloas, W.; Reinecke, M. Thyroid hormone stimulates hepatic IGF-I mRNA expression in a bony fish, the tilapia Oreochromis mossambicus, in vitro and in vivo. Gen. Comp. Endocrinol. 2003, 130, 129–134. [Google Scholar] [CrossRef]
- López-Fernández, J.; Sánchez-Franco, F.; Velasco, B.; Tolón, R.M.; Pazos, F.; Cacicedo, L. Growth hormone induces somatostatin and insulin-like growth factor I gene expression in the cerebral hemispheres of aging rats. Endocrinology 1996, 137, 4384–4391. [Google Scholar] [CrossRef]
- Pedroso, F.L.; Fukada, H.; Masumoto, T. In vivo and in vitro effect of recombinant salmon growth hormone treatment on IGF-I and IGFBPs in yellowtail Seriola quinqueradiata. Fish. Sci. 2009, 75, 887–894. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, R.-L.; Liang, R.; Luo, Y.-Y.; Ruan, Z.-H.; Li, Y.-F.; Liu, W.-S. Growth Axis Somatostatin, Growth Hormone Receptor, and Insulin-like Growth Factor-1 Genes Express and Are Affected by the Injection of Exogenous Growth Hormone in Chinemys reevesii. Genes 2023, 14, 2032. https://doi.org/10.3390/genes14112032
Xie R-L, Liang R, Luo Y-Y, Ruan Z-H, Li Y-F, Liu W-S. Growth Axis Somatostatin, Growth Hormone Receptor, and Insulin-like Growth Factor-1 Genes Express and Are Affected by the Injection of Exogenous Growth Hormone in Chinemys reevesii. Genes. 2023; 14(11):2032. https://doi.org/10.3390/genes14112032
Chicago/Turabian StyleXie, Rui-Lin, Rui Liang, Yuan-Yuan Luo, Zhuo-Hao Ruan, Yi-Fu Li, and Wen-Sheng Liu. 2023. "Growth Axis Somatostatin, Growth Hormone Receptor, and Insulin-like Growth Factor-1 Genes Express and Are Affected by the Injection of Exogenous Growth Hormone in Chinemys reevesii" Genes 14, no. 11: 2032. https://doi.org/10.3390/genes14112032
APA StyleXie, R.-L., Liang, R., Luo, Y.-Y., Ruan, Z.-H., Li, Y.-F., & Liu, W.-S. (2023). Growth Axis Somatostatin, Growth Hormone Receptor, and Insulin-like Growth Factor-1 Genes Express and Are Affected by the Injection of Exogenous Growth Hormone in Chinemys reevesii. Genes, 14(11), 2032. https://doi.org/10.3390/genes14112032