
Three Blind Moles: Molecular Evolutionary Insights on the Tempo and Mode of Convergent Eye Degeneration in Notoryctes typhlops (Southern Marsupial Mole) and Two Chrysochlorids (Golden Moles)

Abstract
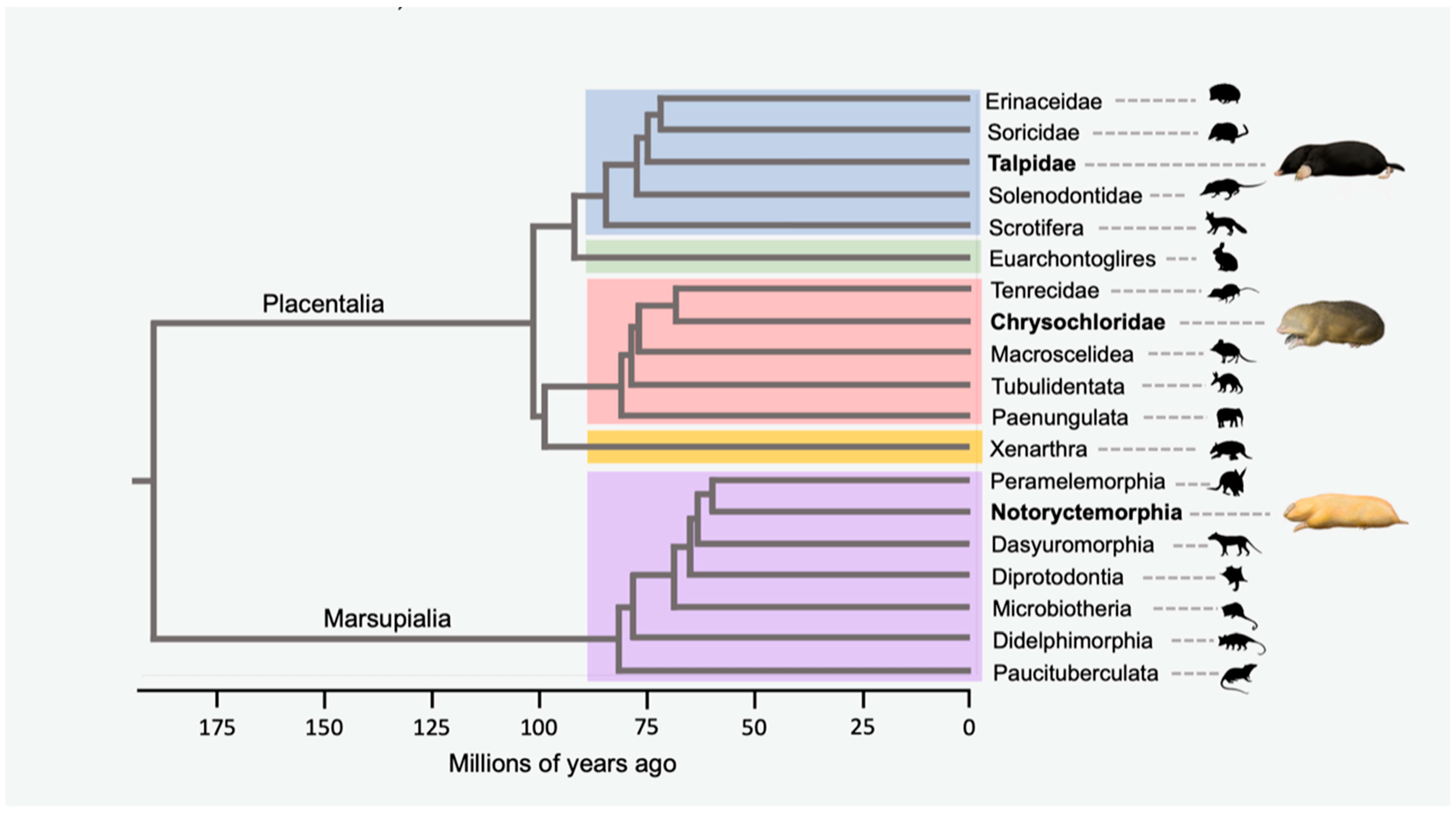
1. Introduction
2. Materials and Methods
2.1. Gene Sampling
2.2. Taxon Sampling
2.3. BLAST Searches and Data Collection
2.4. New Genome Data
2.5. Alignments and Tabulation of Inactivating Mutations
2.6. Phylogenetic Analyses
2.7. Selection Analyses
2.8. Estimation of Gene Inactivation Times
3. Results
3.1. Alignments, Gene Trees and Species Trees
3.2. Gene Inactivation in the Marsupial Mole

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene (# of Marsupial Exons/# of Afrothere Exons) | Inactivating Mutations | |||
|---|---|---|---|---|
| Notoryctes typhlops | Chrysochloridae | C. asiatica | A. hottentotus | |
| ||||
| CNGA3 (8/7) | 471–472D (E5) | None | None | 2074–2076S (E7 A) |
| CNGB3 (18/18) | 1000D (E9, NYGC only) 1478–1575D (E13), 1654–1658I (E14), 1823–1827D (E16) | 1811–1813S (E16), 2069–2071S (E17) | In1Do (AT), 655–657S (E6), 751–753S (E6), In8Ac (GG), 1000–1002S (E9), 1147–1149S (E10), 1192–1194S (E11), In12Do (AT), 1874–1876S (E16), 1955D (E17), 2021–2023S (E17), 2086D (E17), 2107–2108D (E18), 2460I (E18) | 451–453S (E4), 640–642S (E6), 1404–1407D (E12), 1659D (E14), 1759I (E15), 2085–2088D (E17), 2213–2232D (E18), 2242–2427I (E18) |
| GNAT2 (8/8) | 1–118D (E1), 135–136D (E2) | None | 1–118D (E1 B), 355–356D (E4), 827I (E7), 887–888D (E8), 974–976S (E8) | None |
| GNB3# (9/9) | None | None | None | None |
| GNGT2 (2/2) | None | None | None | None |
| OPN1LW (=LWS) (6/6) | None | 180–182S (E2), 241D (E2), 1043D (E6) | 1–3SCM (E1), 390–392SBCM (E2), 556–560D (E3), In4E5BD (97 bp of In4 and 1 bp (750) of E5), E5In5BD (6 bp (984–989) of E5 and 33 bp of In5), 1027I (E6) | In1E2BD (last 12 bp of In1 and first 13 bp (116–128) of E2), 142D (E2), 163–164I (E2), 286–326D (E2), 479D (E3), In4Do (TT), In4E5BD (121 bp of In4 and 22 bp (750–771) of E5), 1026D (E6), 1099–1101TCM (E6) |
| OPN1SW (=SWS1) (5/5) | None | 263–271STSD (E1) | 773–774D (E4) | 219I (E1), 388–389D (E2), 710–712S (E4), 878–880CBSM (E4), 932–934S (E5) |
| PDE6C (22/22) | None | None | 3D (E1), 175D (E1), 634–636S (E2), In10Do (GG), 1109–1112I (E8), 1754–1863D (E14), 1943–1945S (E15), 2624–2626TCM (E22) | None |
| PDE6H (3/3) | None | None | In2Ac (AA), 240–252D (E3, also 27 bp in 3′ UTR) | None |
| ||||
| ARR3 (16/0) | 1075D (E14), In14Do (CT), E15In15BD (2 bp (1084,1085) of E15, 7 bp of In15), In15Ac (AC), 1164D (E16) | Pseudogene in Afrotheria | Pseudogene in Afrotheria | Pseudogene in Afrotheria |
| GRK7 (4/4) | 1306–1308S (E3, DNA Zoo only) | 1567–1569S (E4), 1631–1662D (E4) | 1–1060NBR/NMR (E1–2), 1234–1237I (E3), 1365D (E4) | 1–3SCM (E1), 13–16I (E1), 497–499S (E1), 936D (E2), 962D (E2), 1236–1237I (E3) |
| SLC24A2# (10/10) | None | None | None | None |
| ||||
| CNGA1 (8/8) | None | None | None | None |
| CNGB1 (18/32) | None | None | None | None |
| GNAT1 (8/8) | None | None | None | None |
| GNB1# (9/9) | None | None | None | None |
| GNGT1 (2/2) | None | None | None | None |
| PDE6A (22/22) | None | None | None | None |
| PDE6B (22/22) | In6Do (AT, NYGC only) | None | None | None |
| PDE6G (3/3) | None | None | None | None |
| RHO (5/5) | None | None | None | None |
| ||||
| GRK1 (7/7) | None | None | None | None |
| SAG (15/15) | 8–23D (E1), 173–366D (E4), 412–414S (E5, NYGC only), 805I (E10), 1106–1108S (E15, NYGC only) | None | None | None |
| SLC24A1 (8/9) | 408D (E1, NYGC only), 414I (E1), 515–517S (E1), 1634I (E1), 1670D (E1), 2387–2684D (E5), 4293–4295S (E8) | None | 2110–2132D (E4) | 2111–2132D (E4) |
| Gene (# of Marsupial Exons/# of Afrothere Exons) | Inactivating Mutations | |||
|---|---|---|---|---|
| Notoryctes typhlops | Chrysochloridae | C. asiatica | A. hottentotus | |
| ||||
| CRYAA (3/3) | 376–378S (E3) | None | In1Do (CT) | None |
| CRYAB (3/3) | None | None | None | None |
| ||||
| CRYBA1 (6/6) | 91–93S (E2), 324–330D (E4), 405–412D (E5), 433–435S (E5), 466D (E5), 593I (E6, NYGC only), 625I (E6), 646–648TCM (E6) | 174–216D (E3), 502–649NBR/NRM (E6) | 84I (E2), 168–169D (E3), 254–256S (E4), 420D (E5), 483–495S (E5), In5Do (TT) | 1–3SCM (E1), 359–501NRM (E5), 570–573D (E6), 586–589D (E6), 647–649TCM (E6) |
| CRYBA4 (5/5) | 307–309S (E4, NYGC only), 427–429S (E4) | NBR/NMR | None | None |
| CRYBB1 (5/5) | 1–3SCM (E1), 203D (E2), 301–303S (E3), 385–387S (E3), 523–525S (E4), In4Do (AT) | 232–234S (E2), 542–546D (E4), 569–570D (E4), 575–762NBR/NMR (E5) | 14D (E1), 194–212D (E2), 518D (E4), In4Do (AT) | 328D (E3), 441–451D (E4), In4Do (AG) |
| CRYBB2 (5/5) | In2Ac (GG, NYGC only), 241–243S (E3), In4Do (AT) | None | 25–51D (E1), 252–267D (E3), 397D (E4), 467D (E5), 619–621TCM (E5) | In1Do (GG), 109–128D (E2) |
| CRYBB3 (5/5) | 439–441S (E4, DNA Zoo only), 635–636TCM (E5) | In1E2BD (14 bp of I1 and 9 bp [77,78,79,80,81,82,83,84,85] of E2), 177D (E2), 265–267S (E3), 437–439S (E4), 638–640TCM | 115–117S (E2), 129–132D (E2), 418–420S (E4), 443–453D (E4) | 293–294D (E3) |
| ||||
| CRYGS (3/3) | 19–21S (E1, NYGC only), 121–123S (E2), 267–297D (E3), 511–513S (E3) | 151–154I (E2), 413–415S (E3), 492D (E3), 519–526I (E3) | 200–202S (E2) | 194–196S (E2), 244D (E2) |
| ||||
| BFSP1 (8/8) | 355–357S (E1), In1Ac (A-), 424–519NBR/NMR (E3), In5Ac (TG), 1105–1108D (E8), 1255–1257S (E8), 1582–1584S (E8), 1642–1644S (E8), 1725D (E8) | 16I (E1), 356D (E1), In1Ac (TG), 538–540S (E3) | 366–367D (E1), E2INV A, 409–411S (E2), 530D (E3) | 38–102D (E1), 279–289I (E1), 343–345S (E1), 366–375D (E1) |
| BFSP2 (7/7) | 145D (E1), 466–548NBR/NMR (E2), 571–573S (E3), 868–999NBR/NMR (E5), In6Ac (GA) | None | 999–1002I (E5), In5Do (GG), 1120–1121I (E6), 1164I (E6), 1193D (E6), In6Do (AT) | 1180–1181D (E6) |
| GJA8 (1/1) | 302D (E1), 561–562I (E1), 980–983D (E1) | None | 22–198AfSI (E1), 309–311S (E1), 889I (E1), 991–997D (E1), 1311–1314D (E1), 1524–1525D (E1), 1555D (E1) | NRM B |
| LIM2 (4/4) | None | None | 1–3SCM (E1), 156–160I (E1), 162–164S (E1) | None |
| MIP (4/4) | 62D (E1), 324–331I (E1), 567–569S (E3) | In3Ac (GG), 663–665S (E4), 708–711D (E4) | 165D (E1), 426–428S (E2), 788D (E4) | 328–335I (E1), In2Do (AT), 556–574D (E3), 764–788D (E4) |
| KRT12 (8/8) | 774D (E2), In2Ac (AC), 1108–1110S (E4), 1450I (E6) | 250–272D (E1), 454–456S (E1), 523–524D (E1), | In2Ac (GG), 637–639S (E3), 692–698D (E3), 863–872I (E4), 1003I (E5), 1238I (E6), 1322I (E6), In6Ac (CG), 1415–1417S (E8) | 17–20D (E1), 535–537S (E1), In1Ac (Ac), 638–630D (E3), 945I (E4), In7Ac (GG), |
3.3. Gene Inactivation in Golden Moles
3.4. Gene Inactivation in Other Taxa
3.5. Selection Analyses and the Timing of Gene Inactivation
4. Discussion
4.1. Patterns of Inactivation in the Eye Genes of Golden Moles and Marsupial Moles
4.2. The Tempo of Convergent Eye Degeneration in Golden Moles and Marsupial Moles
4.3. Notoryctid Evolution and the Fossil Record
4.4. Chrysochlorid Evolution and the Fossil Record
4.5. Circadian Photoentrainment in Golden Moles and Marsupial Moles
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stayton, C.T. The definition, recognition, and interpretation of convergent evolution, and two new measures for quantifying and assessing the significance of convergence. Evolution 2015, 69, 2140–2153. [Google Scholar] [CrossRef]
- Stayton, C.T. What does convergent evolution mean? The interpretation of convergence and its implications in the search for limits to evolution. Interface Focus 2015, 5, 20150039. [Google Scholar] [CrossRef] [PubMed]
- Meredith, R.W.; Gatesy, J.; Murphy, W.J.; Ryder, O.A.; Springer, M.S. Molecular decay of the tooth gene enamelin (ENAM) mirrors the loss of enamel in the fossil record of placental mammals. PLoS Genet. 2009, 5, e1000634. [Google Scholar] [CrossRef]
- Sharma, V.; Hecker, N.; Roscito, J.G.; Foerster, L.; Langer, B.E.; Hiller, M. A genomics approach reveals insights into the importance of gene losses for mammalian adaptations. Nat. Commun. 2018, 9, 1215. [Google Scholar] [CrossRef] [PubMed]
- Kowalczyk, A.; Chikina, M.; Clark, N. Complementary evolution of coding and noncoding sequence underlies mammalian hairlessness. eLife 2022, 11, e76911. [Google Scholar] [CrossRef]
- Losos, J.B. Convergence, adaptation, and constraint. Evolution 2011, 65, 1827–1840. [Google Scholar] [CrossRef] [PubMed]
- Muschick, M.; Indermaur, A.; Salzburger, W. Convergent evolution within the adaptive radiation of cichlid fishes. Curr. Biol. 2012, 22, 2362–2368. [Google Scholar] [CrossRef]
- Fish, F.E. Aquatic locomotion: Environmental constraints that drive convergent evolution. In Convergent Evolution: Animal Form and Function; Bels, V.L., Russell, A.P., Eds.; Fascinating Life Sciences: Cham, Switzerland, 2023; pp. 477–522. [Google Scholar]
- Ord, T.J.; Stamps, J.A.; Losos, J.B. Convergent evolution in the territorial communication of a classic adaptive radiation. Caribbean Anolis lizards. Anim. Behav. 2013, 85, 1415–1426. [Google Scholar] [CrossRef]
- Springer, M.S.; Kirsch, J.A.W.; Case, J.A. The chronicle of marsupial evolution. In Molecular Evolution and Adaptive Radiation; Givnish, T.J., Sytsma, K.J., Eds.; Cambridge University Press: Cambridge, MA, USA, 1997; pp. 129–161. [Google Scholar]
- Meredith, R.W.; Janecka, J.E.; Gatesy, J.; Ryder, O.A.; Fisher, C.A.; Teeling, E.C.; Goodbla, A.; Eizirik, E.; Simao, T.L.L.; Stadler, T.; et al. Impacts of the Cretaceous Terrestrial Revolution and KPg extinction on mammal diversification. Science 2011, 334, 521–524. [Google Scholar] [CrossRef]
- Shimer, H.W. Adaptations to aquatic, arboreal, fossorial and cursorial habits in mammals. III. Fossorial adaptations. Am. Nat. 1903, 37, 819–825. [Google Scholar] [CrossRef]
- Warburton, N.M. Functional morphology of marsupial moles (Marsupialia, Notoryctidae). Verh. Des Nat. Ver. Hambg. 2006, 42, 39–149. [Google Scholar]
- Nowak, R.M. Walker’s Mammals of the World: Monotremes, Marsupials, Afrotherians, Xenarthrans, and Sundatherians; Johns Hopkins University Press: Baltimore, MD, USA, 2018. [Google Scholar]
- Cope, E.D. On the habits and affinities of the new Australian mammal, Notoryctes typhlops. Am. Nat. 1892, 26, 121–128. [Google Scholar] [CrossRef]
- Sweet, G. The eyes of Chrysochloris hottentota and C. asiatica. J. Cell Sci. 1909, 2, 327–338. [Google Scholar] [CrossRef]
- Sweet, G. Contributions to our knowledge of the anatomy of Notoryctes typhlops, Stirling. Part III.–The eye. Q. J. Mier. Sc. London 1906, 50, 547–571. [Google Scholar]
- Johnson, K.A.; Walton, D.W. Notoryctidae. In Fauna of Australia: Volume 1B Mammalia; Walton, D.W., Richardson, B.J., Eds.; AGPS: Canberra, Australia, 1989; pp. 1–24. [Google Scholar]
- Bhagwandin, A.; Ndlovu, N.; Bronner, G.N.; Bennett, N.C.; Manger, P.R. The hypercholinergic brain of the Cape golden mole (Chrysochloris asiatica). J. Chem. Neuroanat. 2020, 110, 101856. [Google Scholar] [CrossRef] [PubMed]
- Clark, W.L.G. The brain of the Insectivora. Proc. Zool. Soc. Lond. 1932, 102, 975–1013. [Google Scholar] [CrossRef]
- Gubbay, V. A comparison of the development of the rudimentary eye of Eremitalpa granti (Broom) with that of the normal eye of Elephantulus myurus jamesoni (Chubb). S. Afr. J. Sci. 1956, 52, 182–186, 193–198. [Google Scholar]
- Stephan, H.; Baron, G.; Frahm, H.D. Comparative Brain Research in Mammals. Vol. 1: Insectivora; Springer: New York, NY, USA, 1991. [Google Scholar]
- Perrin, M.R.; Fielden, L.J. Eremitalpa granti. Mamm. Species 1999, 629, 1–4. [Google Scholar] [CrossRef]
- Darwin, C. On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life; John Murray: London, UK, 1859. [Google Scholar]
- Spencer, B. Report on the Work of the Horn Expedition to Central Australia. Part I.—Introduction, Narrative, Summary of Results, Supplement to Zoological Report, Map; Ford and Son: Carlton, Australia, 1896. [Google Scholar]
- Pepper, M.; Keogh, J.S. Life in the “dead heart” of Australia: The geohistory of the Australian deserts and its impact on genetic diversity of arid zone lizards. J. Biogeogr. 2021, 48, 716–746. [Google Scholar] [CrossRef]
- Asher, R.J.; Maree, S.; Bronner, G.; Bennett, N.C.; Bloomer, P.; Czechowski, P.; Meyer, M.; Hofrreiter, M. A phylogenetic estimate for golden moles (Mammalia, Afrotheria, Chrysochloridae). BMC Evol. Biol. 2010, 10, 69. [Google Scholar] [CrossRef]
- Heritage, S.; Seiffert, E.R.; Borths, M.R. Recommended fossil calibrations for time-scaled molecular phylogenies of Afrotheria. Afrotherian Conserv. 2021, 17, 1–8. [Google Scholar]
- Springer, M.S.; Burk, A.; Kavanagh, J.R.; Waddell, V.G.; Stanhope, M.J. The interphotoreceptor retinoid binding protein gene in therian mammals: Implications for higher level relationships and evidence for loss of function in the marsupial mole. Proc. Natl. Acad. Sci. USA 1997, 94, 13754–13759. [Google Scholar] [CrossRef]
- Emerling, C.A.; Springer, M.S. Eyes underground: Regression of visual protein networks in subterranean mammals. Mol. Phylogenet. Evol. 2014, 78, 260–270. [Google Scholar] [CrossRef] [PubMed]
- Carmona, F.D.; Glösmann, M.; Ou, J.; Jiménez, R.; Collinson, J.M. Retinal development and function in a ‘blind’ mole. Proc. R. Soc. B 2010, 277, 1513–1522. [Google Scholar] [CrossRef] [PubMed]
- Lamb, T.D. Evolution of the genes mediating phototransduction in rod and cone photoreceptors. Prog. Retin. Eye Res. 2020, 76, 100823. [Google Scholar] [CrossRef] [PubMed]
- Lamb, T.D.; Patel, H.R.; Chuah, A.; Hunt, D.M. Evolution of the shut-off steps of vertebrate phototransduction. Open Biol. 2018, 8, 170232. [Google Scholar] [CrossRef]
- Lamb, T.D.; Hunt, D.M. Evolution of the calcium feedback steps of vertebrate phototransduction. Open Biol. 2018, 8, 180119. [Google Scholar] [CrossRef]
- Invergo, B.M.; Montanucci, L.; Laayouni, H.; Bertranpetit, J. A system-level, molecular evolutionary analysis of mammalian phototransduction. BMC Evol. Biol. 2013, 13, 52. [Google Scholar] [CrossRef]
- Lamb, T.D. Photoreceptor physiology and evolution: Cellular and molecular basis of rod and cone phototransduction. J. Physiol. 2022, 600, 4585–4601. [Google Scholar] [CrossRef]
- Weiss, E.R.; Ducceschi, M.H.; Horner, T.J.; Li, A.; Craft, C.M.; Osawa, S. Species-specific differences in expression of G-protein-coupled receptor kinase (GRK) 7 and GRK1 in mammalian cone photoreceptor cells: Implications for cone cell phototransduction. J. Neurosci. 2001, 21, 9175–9184. [Google Scholar] [CrossRef]
- Ye, Y.; Sun, Z.; Guo, A.; Song, L.S.; Grobe, J.L.; Chen, S. Ablation of the GNB3 gene in mice does not affect body weight, metabolism or blood pressure, but causes bradycardia. Cell. Signal. 2014, 26, 2514–2520. [Google Scholar] [CrossRef][Green Version]
- Ozdemir, A.C.; Wynn, G.M.; Vester, A.; Weitzmann, M.N.; Neigh, G.N.; Srinivasan, S.; Rudd, M.K. GNB3 overexpression causes obesity and metabolic syndrome. PLoS ONE 2017, 12, e0188763. [Google Scholar] [CrossRef]
- Keers, R.; Bonvicini, C.; Scassellati, C.; Uher, R.; Placentino, A.; Giovannini, C.; Rietschel, M.; Henigsberg, N.; Kozel, D.; Mors, O.; et al. Variation in GNB3 predicts response and adverse reactions to antidepressants. J. Psychopharmacol. 2011, 25, 867–874. [Google Scholar] [CrossRef]
- Kumar, R.; Kohli, S.; Alam, P.; Barkotoky, R.; Gupta, M.; Tyagi, S.; Jain, S.K.; Pasha, M.Q. Interactions between the FTO and GNB3 genes contribute to varied clinical phenotypes in hypertension. PLoS ONE 2013, 8, e63934. [Google Scholar] [CrossRef] [PubMed]
- Bian, Q.; Li, H.; Wang, X.; Liang, T.; Zhang, K. Multiomics integrated analysis dientifies SLC24A2 as a potential link between type 2 diabetes and cancer. J. Diabetes Res. 2022, 2022, 4629419. [Google Scholar] [CrossRef]
- Petrovski, S.; Küry, S.; Myers, C.T.; Anyane-Yeboa, K.; Cogné, B.; Bialer, M.; Xia, F.; Hemati, P.; Riviello, J.; Mehaffey, M.; et al. Germline de novo mutations in GNB1 cause severe neurodevelopmental disability, hypotonia, and seizures. Am. J. Hum. Genet. 2016, 98, 1001–1010. [Google Scholar] [CrossRef]
- Steinrücke, S.; Lohmann, K.; Domingo, A.; Rolfs, A.; Bäumer, T.; Spiegler, J.; Hartmann, C.; Münchau, A. Novel GNB1 missense mutation in a patient with generalized dystonia, hypotonia, and intellectual disability. Neurol. Genet. 2016, 2, e106. [Google Scholar] [CrossRef] [PubMed]
- Lohmann, K.; Masuho, I.; Patil, D.N.; Baumann, H.; Hebert, E.; Steinrücke, S.; Trujillano, D.; Skamangas, N.K.; Dobricic, V.; Hüning, I.; et al. Novel GNB1 mutations disrupt assembly and function of G protein heterotrimers and cause global developmental delay in humans. Hum. Mol. Genet. 2017, 26, 1078–1086. [Google Scholar] [CrossRef]
- Hemati, P.; Revah-Politi, A.; Bassan, H.; Petrovski, S.; Bilancia, C.G.; Ramsey, K.; Griffin, N.G.; Bier, L.; Cho, M.T.; Rosello, M.; et al. Refining the phenotype associated with GNB1 mutations: Clinical data on 18 newly identified patients and review of the literature. Am. J. Med. Genet. Part A 2018, 176, 2259–2275. [Google Scholar] [CrossRef] [PubMed]
- Michalakis, S.; Reisert, J.; Geiger, H.; Wetzel, C.; Zong, X.; Bradley, J.; Spehr, M.; Hüttl, S.; Gerstner, A.; Pfeifer, A.; et al. Loss of CNGB1 protein leads to olfactory dysfunction and subciliary cyclic nucleotide-gated channel trapping. J. Biol. Chem. 2006, 281, 35156–35166. [Google Scholar] [CrossRef] [PubMed]
- Miyashita, Y.; Moriya, T.; Asami, K.; Kubota, T.; Yamada, K. Expression of opsin molecule in cultured murine melanocyte. J. Investig. Dermatol. Symp. Proc. 2001, 6, 54–57. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Son, E.D.; Jung, J.Y.; Choi, H.; Lee, T.R.; Shin, D.W. Violet light down-regulates the expression of specific differentiation markers through rhodopsin in normal human epidermal keratinocytes. PLoS ONE 2013, 8, e73678. [Google Scholar] [CrossRef] [PubMed]
- de Assis, L.V.M.; Moraes, M.N.; da Silveira Cruz-Machado, S.; Castrucci, A.M.L. The effect of white light on normal and malignant murine melanocytes: A link between opsins, clock genes, and melanogenesis. Biochim. Biophys. Acta 2016, 1863, 1119–1133. [Google Scholar] [CrossRef]
- Roy, D.; Levi, K.; Kiss, V.; Nevo, R.; Eisenbach, M. Rhodopsin and melanopsin coexist in mammalian sperm cells and activate different signaling pathways for thermotaxis. Sci. Rep. 2020, 10, 112. [Google Scholar] [CrossRef] [PubMed]
- Xiao, W.; Yu, M.; Yuan, Y.; Liu, X.; Chen, Y. Thermotaxis of mammalian sperm. Mol. Hum. Reprod. 2022, 28, gaac027. [Google Scholar] [CrossRef]
- Wistow, G. The human crystallin gene families. Hum. Genom. 2012, 6, 26. [Google Scholar] [CrossRef]
- Rao, N.A.; Saraswathy, S.; Pararajasegaram, G.; Bhat, S.P. Small heat shock protein αA-crystallin prevents photoreceptor degeneration in experimental autoimmune uveitis. PLoS ONE 2012, 7, e33582. [Google Scholar] [CrossRef]
- Billingsley, G.; Santhiya, S.T.; Paterson, A.D.; Ogata, K.; Wodak, S.; Hosseini, S.M.; Manisastry, S.M.; Vijayalakshmi, P.; Gopinath, P.M.; Graw, J.; et al. CRYBA4, a novel human cataract gene, is also involved in microphthalmia. Am. J. Hum. Genet. 2006, 79, 702–709. [Google Scholar] [CrossRef]
- Zhou, G.; Zhou, N.; Hu, S.; Zhao, L.; Zhang, C.; Qi, Y. A missense mutation in CRYBA4 associated with congenital cataract and microcornea. Mol. Vis. 2010, 16, 1019–1024. [Google Scholar] [PubMed]
- Willoughby, C.E.; Shafiq, A.; Ferrini, W.; Chan, L.L.; Billingsley, G.; Priston, M.; Mok, C.; Chandna, A.; Kaye, S.; Héon, E. CRYBB1 mutation associated with congenital cataract and microcornea. Mol. Vis. 2005, 11, 587–593. [Google Scholar]
- Clark, A.R.; Lubsen, N.H.; Slingsby, C. sHSP in the eye lens: Crystallin mutations, cataract and proteostasis. Int. J. Biochem. Cell Biol. 2012, 44, 1687–1697. [Google Scholar] [CrossRef] [PubMed]
- Perng, M.D.; Zhang, Q.; Quinlan, R.A. Insights into the beaded filament of the eye lens. Exp. Cell Res. 2007, 313, 2180–2188. [Google Scholar] [CrossRef]
- Kerscher, S.; Church, R.L.; Boyd, Y.; Lyon, M.F. Mapping of four mouse genes encoding eye lens-specific structural, gap junction, and integral membrane proteins: Cryba1 (CrystallinβA3/A1), Crybb2 (CrystallinβB2), Gja8 (MP70), and Lim2 (MP19). Genomics 1995, 29, 445–450. [Google Scholar] [CrossRef]
- Mulders, S.M.; Preston, G.M.; Deen, P.M.; Guggino, W.B.; van Os, C.H.; Agre, P. Water channel properties of major intrinsic protein of lens. J. Biol. Chem. 1995, 270, 9010–9016. [Google Scholar] [CrossRef]
- Chepelinsky, A.B. Structural function of MIP/aquaporin 0 in the eye lens; genetic defects lead to congenital inherited cataracts. In Aquaporins; Beitz, E., Ed.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 265–297. [Google Scholar]
- Liu, C.Y.; Zhu, G.; Converse, R.; Kao, C.W.; Nakamura, H.; Tseng, S.C.; Mui, M.M.; Seyer, J.; Justice, M.J.; Stech, M.E. Characterization and chromosomal localization of the cornea-specific murine keratin gene Krt1. 12. J. Biol. Chem. 1994, 269, 24627–24636. [Google Scholar] [CrossRef]
- Sasamoto, Y.; Hayashi, R.; Park, S.J.; Saito-Adachi, M.; Suzuki, Y.; Kawasaki, S.; Quantock, A.J.; Nakai, K.; Tsujikawa, M.; Nishida, K. PAX6 isoforms, along with reprogramming factors, differentially regulate the induction of cornea-specific genes. Sci. Rep. 2016, 6, 20807. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef]
- Katoh, K.; Toh, H. Recent developments in the MAFFT multiple sequence alignment program. Brief. Bioinform. 2008, 9, 286–298. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 1–19. [Google Scholar] [CrossRef]
- Bowmaker, J.K. Evolution of vertebrate visual pigments. Vision Res. 2008, 48, 2022–2041. [Google Scholar] [CrossRef]
- Yokoyama, S. Evolution of dim-light and color vision pigments. Annu. Rev. Genomics Hum. Genet. 2008, 9, 259–282. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML Version 8: A tool for phylogenetic analysis and postanalysis of large phylogenies. Bioinformatics 2014, 20, 1312–1313. [Google Scholar] [CrossRef]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A rapid bootstrap algorithm for the RAxML web servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef]
- Foley, N.M.; Mason, V.C.; Harris, A.J.; Bredemeyer, K.R.; Damas, J.; Lewin, H.A.; Eizirik, E.; Gatesy, J.; Karlsson, E.K.; Lindblad-Toh, K.; et al. A genomic timescale for placental mammal evolution. Science 2023, 380, eabl8189. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z. PAML 4: Phylogenetic analysis by maximum likelihood. Mol. Biol. Evol. 2007, 24, 1586–1591. [Google Scholar] [CrossRef] [PubMed]
- Randall, J.G.; Gatesy, J.; Springer, M.S. Molecular evolutionary analyses of tooth genes support sequential loss of enamel and teeth in baleen whales (Mysticeti). Mol. Phylogenet. Evol. 2022, 171, 107463. [Google Scholar] [CrossRef] [PubMed]
- Duchêne, D.A.; Bragg, J.G.; Duchêne, S.; Neaves, L.E.; Potter, S.; Moritz, C.; Johnson, R.N.; Ho, S.Y.; Eldridge, M.D. Analysis of phylogenomic tree space resolves relationships among marsupial families. Syst. Biol. 2018, 67, 400–412. [Google Scholar] [CrossRef] [PubMed]
- Springer, M.S.; Emerling, C.A.; Fugate, N.; Patel, R.; Starrett, J.; Morin, P.A.; Hayashi, C.; Gatesy, J. Inactivation of cone-specific phototransduction genes in rod monochromatic cetaceans. Front. Ecol. Evol. 2016, 4, 61. [Google Scholar] [CrossRef]
- Springer, M.S.; Guerrero-Juarez, C.F.; Huelsmann, M.; Collin, M.A.; Danil, K.; McGowen, M.R.; Oh, J.W.; Ramos, R.; Hiller, M.; Plikus, M.V.; et al. Genomic and anatomical comparisons of skin support independent adaptation to life in water by cetaceans and hippos. Curr. Biol. 2021, 31, 2124–2139. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.; Michaelides, M.; Aligianis, I.A.; Ainsworth, J.R.; Mollon, J.D.; Maher, E.R.; Moore, A.T.; Hunt, D.M. Achromatopsia caused by novel mutations in both CNGA3 and CNGB3. J. Med. Genet. 2004, 41, e20. [Google Scholar] [CrossRef] [PubMed]
- Perng, M.D.; Quinlan, R.A. Seeing is believing! The optical properties of the eye lens are dependent upon a functional intermediate filament cytoskeleton. Exp. Cell Res. 2005, 305, 1–9. [Google Scholar] [CrossRef]
- Yi, J.; Yun, J.; Li, Z.K.; Xu, C.T.; Pan, B.R. Epidemiology and molecular genetics of congenital cataracts. Int. J. Ophthalmol. 2011, 4, 422–432. [Google Scholar]
- Deng, H.; Yuan, L. Molecular genetics of congenital nuclear cataract. Eur. J. Med. Genet. 2014, 57, 113–122. [Google Scholar] [CrossRef]
- Heavner, W.; Pevny, L. Eye development and retinogenesis. Cold Spring Harb. Perspect. Biol. 2012, 4, a008391. [Google Scholar] [CrossRef]
- Chase, H.B. Studies on an anophthalmic strain of mice. III. Results of crosses with other strains. Genetics 1942, 27, 339–348. [Google Scholar] [CrossRef]
- Mitchell, K.J.; Pratt, R.C.; Watson, L.N.; Gibb, G.C.; Llamas, B.; Kasper, M.; Edson, J.; Hopwood, B.; Male, D.; Armstrong, K.N.; et al. Molecular phylogeny, biogeography, and habitat preference evolution of marsupials. Mol. Biol. Evol. 2014, 31, 2322–2330. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, M.E.; Ehrhart, T.L.; Berson, E.L.; Dryja, T.P. Mutation spectrum of the gene encoding the beta subunit of rod phosphodiesterase among patients with autosomal recessive retinitis pigmentosa. Proc. Natl. Acad. Sci. USA 1995, 92, 3249–3253. [Google Scholar] [CrossRef]
- Han, J.; Dinculescu, A.; Dai, X.; Du, W.; Smith, W.C.; Pang, J. The history and role of naturally occurring mouse models with Pde6b mutations. Mol. Vis. 2013, 19, 2579–2589. [Google Scholar]
- Marconi, S.; Stout, J.T. PDE6B Mutation-associated inherited retinal disease. Int. Ophthalmol. Clin. 2021, 61, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Johnson, G.L.I. Contributions to the comparative anatomy of the mammalian eye, chiefly based on ophthalmoscopic examination. Philos. Trans. Roy. Soc. B 1901, 194, 1–82. [Google Scholar]
- Watson, G.A. The mammalian cerebral cortex, with special reference to its comparative histology. 1. Order Insectivora.—Preliminary communication. Proc. Roy. Soc. Lond. B 1906, 77, 150–160. [Google Scholar]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef]
- Da Silva, J.D.; Costa, M.D.; Almeida, B.; Lopes, F.; Maciel, P.; Teixeira-Castro, A. Case report: A novel GNB1 mutation causes global developmental delay with intellectual disability and behavioral disorders. Front. Neurol. 2021, 12, 735549. [Google Scholar] [CrossRef]
- Moraes, M.N.; de Assis, L.V.M.; Provencio, I.; Castrucci, A.M.D.L. Opsins outside the eye and the skin: A more complex scenario than originally thought for a classical light sensor. Cell Tissue Res. 2021, 385, 519–538. [Google Scholar] [CrossRef]
- Breazzano, M.P.; Grewal, M.R.; Tsang, S.H.; Chen, R.W. Etiology of retinitis pigmentosa. In Retinitis Pigmentosa; Tsang, S.H., Quinn, P.M., Eds.; Springer: New York, NY, USA, 2022; pp. 15–30. [Google Scholar]
- McLaughlin, S.K.; McKinnon, P.J.; Spickofsky, N.; Danho, W.; Margolskee, R.F. Molecular cloning of G proteins and phosphodiesterases from rat taste cells. Physiol. Behav. 1994, 56, 1157–1164. [Google Scholar] [CrossRef] [PubMed]
- Governini, L.; Semplici, B.; Pavone, V.; Crifasi, L.; Marrocco, C.; De Leo, V.; Arlt, E.; Gudermann, T.; Boekhoff, I.; Luddi, A.; et al. Expression of taste receptor 2 subtypes in human testis and sperm. J. Clin. Med. 2020, 9, 264. [Google Scholar] [CrossRef] [PubMed]
- Vinberg, F.; Wang, T.; Molday, R.S.; Chen, J.; Kefalov, V.J. A new mouse model for stationary night blindness with mutant Slc24a1 explains the pathophysiology of the associated human disease. Hum. Mol. Genet. 2015, 24, 5915–5929. [Google Scholar] [CrossRef]
- Nikonov, S.S.; Brown, B.M.; Davis, J.A.; Zuniga, F.I.; Bragin, A.; Pugh, E.N.; Craft, C.M. Mouse cones require an arrestin for normal inactivation of phototransduction. Neuron 2008, 59, 462–474. [Google Scholar] [CrossRef] [PubMed]
- Chaves, J.M.; Gupta, R.; Srivastava, K.; Srivastava, O. Human alpha A-crystallin missing N-terminal domain poorly complexes with filensin and phakinin. Biochem. Biophys. Res. Commun. 2017, 494, 402–408. [Google Scholar] [CrossRef]
- Beck, R.M.; Warburton, N.M.; Archer, M.; Hand, S.J.; Aplin, K.P. Going underground: Postcranial morphology of the early Miocene marsupial mole Naraboryctes philcreaseri and the evolution of fossoriality in notoryctemorphians. Mem. Mus. Vic. 2016, 74, 151–171. [Google Scholar] [CrossRef][Green Version]
- Archer, M.; Beck, R.; Gott, M.; Hand, S.; Godthelp, H.; Black, K. Australia’s first fossil mole (Notoryctemorphia) resolves controversies about their evolution and palaeoenvironmental origins. Proc. R. Soc. B 2011, 278, 1498–1506. [Google Scholar] [CrossRef]
- Strömberg, C.A.E. Evolution of grasses and grassland ecosystems. Annu. Rev. Earth Planet. Sci. 2011, 39, 517–544. [Google Scholar] [CrossRef]
- Fujioka, T.; Chappell, J. History of Australian aridity: Chronology in the evolution of arid landscapes. Geol. Soc. Lond. Spec. Publ. 2010, 346, 121–139. [Google Scholar] [CrossRef]
- Fujioka, T.; Chappell, J.; Fifield, L.K.; Rhodes, E.J. Australian desert dune fields initiated with Pliocene–Pleistocene global climatic shift. Geology 2006, 37, 51–54. [Google Scholar] [CrossRef]
- Wyrwoll, K.-H.; Turner, B.L.; Findlater, P. On the origins, geomorphology and soils on the sandplains of south-western Australia. In Plant Life on the Sandplains in Southwest Australia, a Global Biodiversity Hotspot; Lambers, H., Ed.; University of Western Australia Publishing: Crawley, UK, 2014; pp. 3–23. [Google Scholar]
- Pickford, M. Chrysochloridae (Mammalia) from the Lutetian (Middle Eocene) of Black Crow, Namibia. Commun. Geol. Surv. Namib. 2015, 16, 112–120. [Google Scholar]
- Pickford, M. Tiny Tenrecomorpha (Mammalia) from the Eocene of Black Crow, Namibia. Commun. Geol. Surv. Namib. 2019, 21, 15–25. [Google Scholar]
- Pickford, M. Late Eocene Chrysochloridae (Mammalia) from the Sperrgebiet, Namibia. Commun. Geol. Surv. Namib. 2015, 16, 160–199. [Google Scholar]
- Mason, M.J.; Bennett, N.C.; Pickford, M. The middle and inner ears of the Palaeogene golden mole Namachloris: A comparison with extant species. J. Morphol. 2017, 279, 375–395. [Google Scholar] [CrossRef] [PubMed]
- Asher, R.J. Recent additions to the fossil record of tenrecs and golden moles. Afrotherian Conserv. 2019, 15, 4–13. [Google Scholar]
- Crumpton, N.; Kardjilov, N.; Asher, R.J. Convergence vs. specialization in the ear region of moles (Mammalia). J. Morphol. 2015, 276, 900–914. [Google Scholar] [CrossRef] [PubMed]
- Emerling, C.A.; Springer, M.S. Genomic evidence for rod monochromacy in sloths and armadillos suggests early subterranean history for Xenarthra. Proc. Roy. Soc. B 2015, 282, 20142192. [Google Scholar] [CrossRef] [PubMed]
- Buhr, E.D.; Vemaraju, S.; Diaz, N.; Lang, R.A.; Van Gelder, R.N. Neuropsin (OPN5) mediates local light-dependent induction of circadian clock genes and circadian photoentrainment in exposed murine skin. Curr. Biol. 2019, 29, 3478–3487. [Google Scholar] [CrossRef] [PubMed]
- Foster, R.G.; Hughes, S.; Peirson, S.N. Circadian photoentrainment in mice and humans. Biology 2020, 9, 180. [Google Scholar] [CrossRef] [PubMed]
- Gegnaw, S.T.; Sandu, C.; Mazzaro, N.; Mendoza, J.; Bergen, A.A.; Felder-Schmittbuhl, M.P. Enhanced robustness of the mouse retinal circadian clock upon inherited retina degeneration. J. Biol. Rhythms 2022, 37, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Altimus, C.M.; Güler, A.D.; Alam, N.M.; Arman, A.C.; Prusky, G.T.; Sampath, A.P.; Hattar, S. Rod photoreceptors drive circadian photoentrainment across a wide range of light intensities. Nat. Neurosci. 2010, 13, 1107–1112. [Google Scholar] [CrossRef]
- Do, M.T.H.; Kang, S.H.; Xue, T.; Zhong, H.; Liao, H.W.; Bergles, D.E.; Yau, K.W. Photon capture and signalling by melanopsin retinal ganglion cells. Nature 2009, 457, 281–287. [Google Scholar] [CrossRef]
- Sanyal, S.; Jansen, H.G.; De Grip, W.J.; Nevo, E.; de Jong, W.W. The eye of the blind mole rat, Spalax ehrenbergi. Investig. Ophthalmol. Vis. Sci. 1990, 31, 1398–1404. [Google Scholar]
- Goldman, B.D.; Goldman, S.L.; Riccio, A.P.; Terkel, J. Circadian patterns of locomotor activity and body temperature in blind mole-rats, Spalax ehrenbergi. J. Biol. Rhythms 1997, 12, 48–361. [Google Scholar] [CrossRef]
- Erkert, H.G.; Rothmund, E. Differences in temperature sensitivity of the circadian systems of homoiothermic and heterothermic neotropical bats. Comp. Biochem. Physiol. A Physiol. 1981, 68, 383–390. [Google Scholar] [CrossRef]
- Tokura, H.; Aschoff, J. Effects of temperature on the circadian rhythm of pig-tailed macaques Macaca nemestrina. Am. J. Physiol. 1983, 245, R800–R804. [Google Scholar] [CrossRef] [PubMed]
- Aschoff, J.; Tokura, H. Circadian activity rhythms in squirrel monkeys: Entrainment by temperature cycles. J. Biol. Rhythms 1986, 1, 91–99. [Google Scholar] [CrossRef] [PubMed]
- McKechnie, A.E.; Mzilikazi, N. Heterothermy in Afrotropical mammals and birds: A review. Integr. Comp. Biol. 2011, 51, 349–363. [Google Scholar] [CrossRef] [PubMed]
- Riek, A.; Geiser, F. Heterothermy in pouched mammals—A review. J. Zool. 2014, 292, 74–85. [Google Scholar] [CrossRef]





| Feature | Chrysochloridae | Notoryctes typhlops |
|---|---|---|
| Not visible externally; located within the dermis and covered by outer layers of skin and fur | Not visible externally; located beneath the temporalis muscle and covered by skin and fur |
| Absent | Degenerative, abnormal in position, non-striated |
| III and IV absent; no information on VI | Absent |
| Oculomotor and trochlear nuclei greatly reduced and neurons lack clear dendrites; abducens nucleus absent | No information |
| Optic nerve and optic chiasma absent except for remnants of nerve exiting eye in some specimens | Optic nerve and optic chiasma absent except for remnants of nerve or nerve sheath exiting eye in one specimen |
| Degenerate but recognizable | Absent or represented by only a few nuclei |
| Small space that may be pupil | Absent |
| Indistinguishable from sclera and choroid | Indistinguishable from sclera and choroid |
| Degenerate but recognizable; adult lens is an irregular mass of cells without lens fibers | Absent |
| Not well developed or absent | Practically absent or absent |
| Some degeneration but layers are clearly distinguishable in most cases | Undifferentiated mass of cells |
| Well developed | Eye is hollow ball of pigment that may be “greatly changed” retinal pigment layer |
| Most degenerate layer of retina | Absent |
| Layer of cones and rods is recognizable in some, but in other cases all that remains are rod-like structures | Absent |
| Eye Phenotype and Associated Genes | Inactivation Time in Chrysochloridae | Inactivation Time in Notoryctes typhlops |
|---|---|---|
| Cone phototransduction activation (CNGA3, CNGB3, GNAT2, GNGT2, OPN1LW, OPN1SW, PDE6C, PDE6H) | 16.21 | 5.38 |
| Cone phototransduction recovery (ARR3 *, GRK7) | 40.31 | 38.85 |
| Rod phototransduction activation (CNGA1, GNAT1, GNGT1, PDE6A, PDE6B, PDE6G) | No inactivated genes | 3.39 |
| Rod phototransduction recovery (GRK1, SAG, SLC24A1) | 31.43 | 1.52 |
| Lens/cornea development and structure (CRYAA, CRYBA1, CRYBA4 #, CRYBB1, CRYBB2, CRYBB3, CRYGS, BFSP1, BFSP2, GJA8, LIM2, MIP, KRT12) | 26.03 | 17.84 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Springer, M.S.; Emerling, C.A.; Gatesy, J. Three Blind Moles: Molecular Evolutionary Insights on the Tempo and Mode of Convergent Eye Degeneration in Notoryctes typhlops (Southern Marsupial Mole) and Two Chrysochlorids (Golden Moles). Genes 2023, 14, 2018. https://doi.org/10.3390/genes14112018
Springer MS, Emerling CA, Gatesy J. Three Blind Moles: Molecular Evolutionary Insights on the Tempo and Mode of Convergent Eye Degeneration in Notoryctes typhlops (Southern Marsupial Mole) and Two Chrysochlorids (Golden Moles). Genes. 2023; 14(11):2018. https://doi.org/10.3390/genes14112018
Chicago/Turabian StyleSpringer, Mark S., Christopher A. Emerling, and John Gatesy. 2023. "Three Blind Moles: Molecular Evolutionary Insights on the Tempo and Mode of Convergent Eye Degeneration in Notoryctes typhlops (Southern Marsupial Mole) and Two Chrysochlorids (Golden Moles)" Genes 14, no. 11: 2018. https://doi.org/10.3390/genes14112018
APA StyleSpringer, M. S., Emerling, C. A., & Gatesy, J. (2023). Three Blind Moles: Molecular Evolutionary Insights on the Tempo and Mode of Convergent Eye Degeneration in Notoryctes typhlops (Southern Marsupial Mole) and Two Chrysochlorids (Golden Moles). Genes, 14(11), 2018. https://doi.org/10.3390/genes14112018