
Sequencing and Phylogenetic Analysis of the Chloroplast Genome of Three Apricot Species

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Material Collection, DNA Extraction, and Sequencing
2.2. Chloroplast Genome Assembly and Annotation
2.3. Repeat Sequences and Simple Sequence Repeat (SSR) Analysis
2.4. Chloroplast Genome Comparison and Analysis of Variations in IR/SC Boundaries
2.5. Phylogenetic Analysis
3. Results
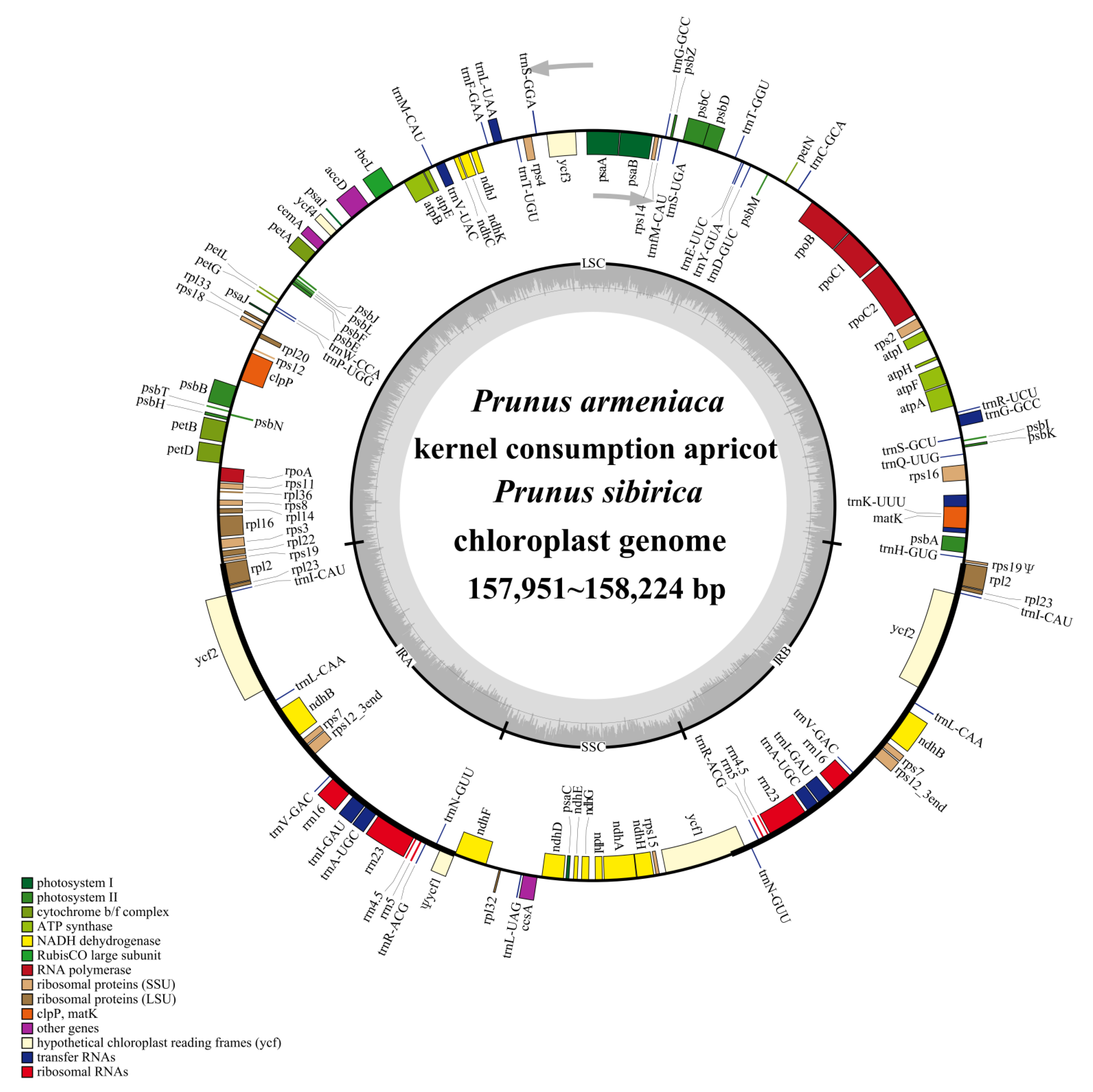
3.1. Organization and Features of the Chloroplast Genomes
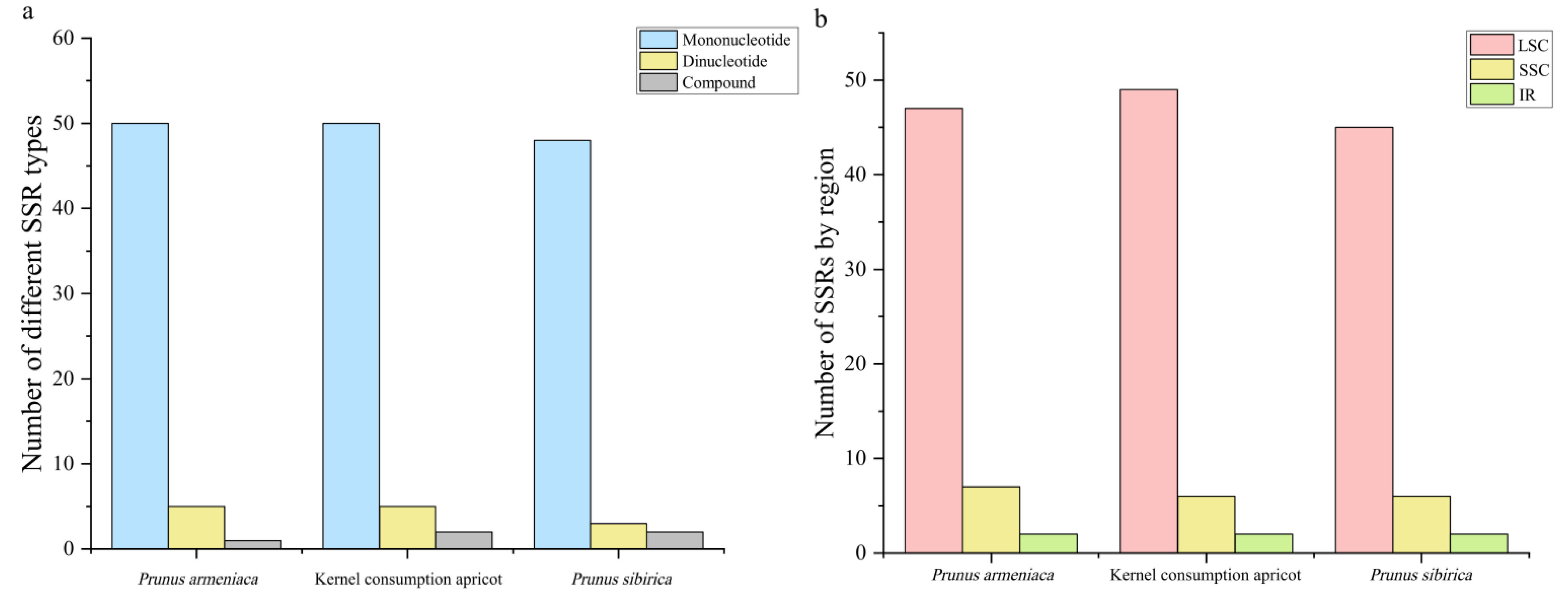
3.2. Characterization of Repeat Sequences and SSRs
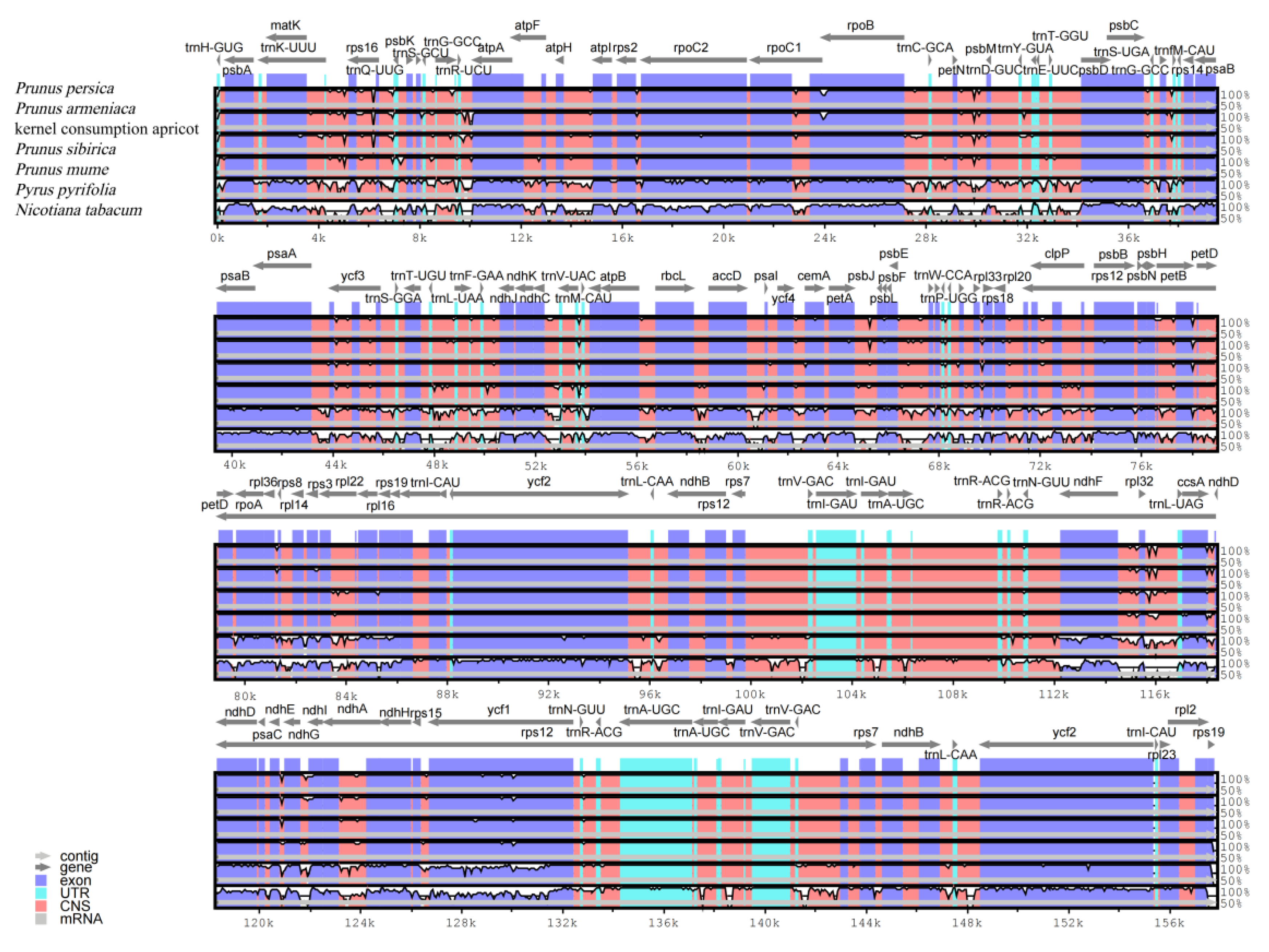
3.3. Comparative Analysis of Apricot Chloroplast Genomes
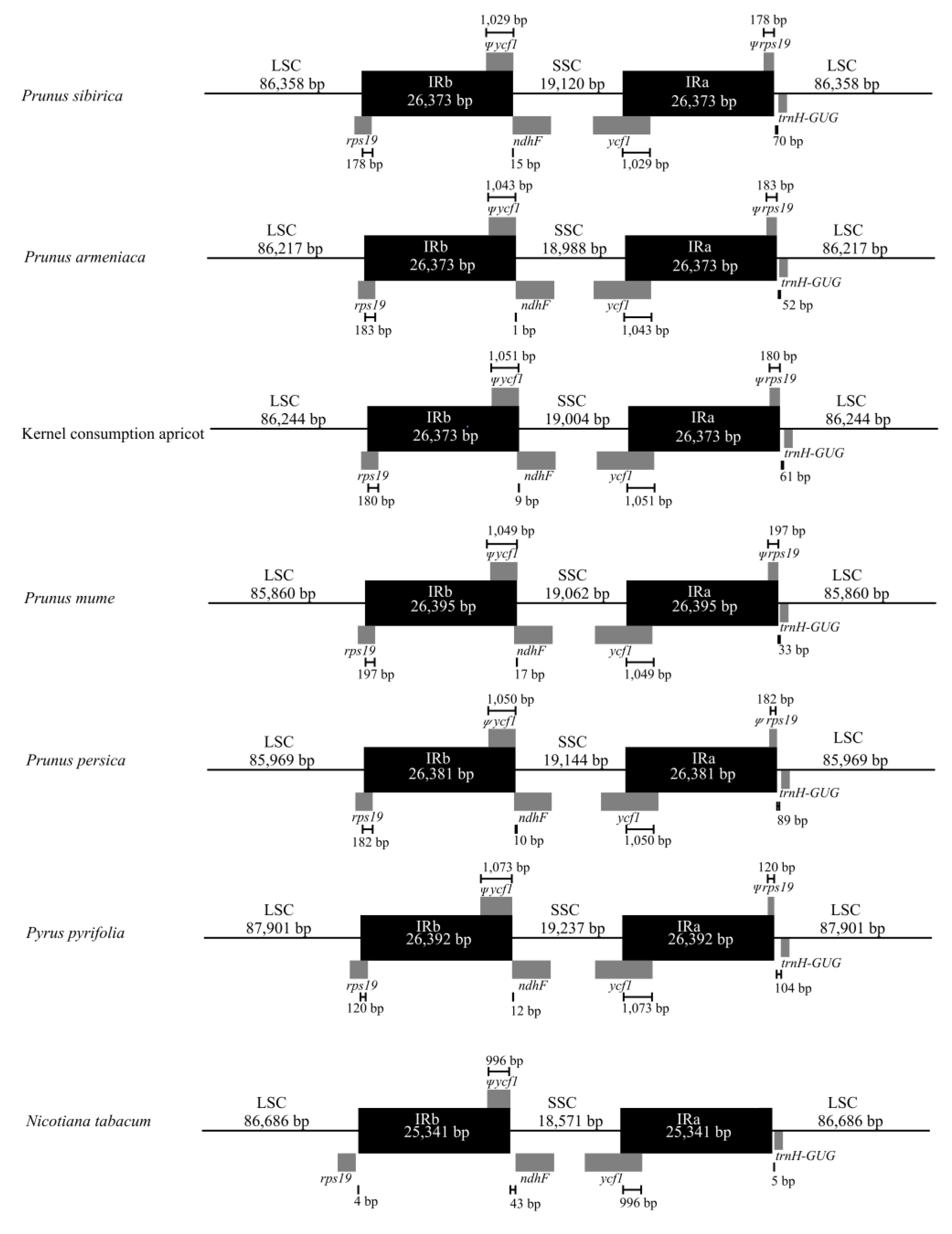
3.4. Analysis of Variations in the IR/SC Boundaries
3.5. Phylogenetic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Basile, B.; Mataffo, A.; Forlani, M.; Corrado, G. Diversity in Morphometric, Pomological, and Fruit-Quality Traits of apricots (Prunus armeniaca) Traditional Varieties: Implications for Landrace Differentiation at Regional Scale. Diversity 2022, 14, 608. [Google Scholar] [CrossRef]
- Rehder, A. Manual of Cultivated Trees and Shrubs Hardy in North America, Exclusive of the Subtropical and Warmer Temperate Regions; Macmillan: New York, NY, USA, 1940. [Google Scholar]
- Mehlenbacher, S.A.; Cociu, V.; Hough, F.L. Apricots (Prunus). Acta Hortic. 1991, 290, 65–110. [Google Scholar] [CrossRef]
- Júlia, H.; Andrzej, P.; Attila, H. Origin and dissemination of the pollen-part mutated SC haplotype which confers self-compatibility in apricot (Prunus armeniaca). New Phytol. 2007, 176, 792–803. [Google Scholar]
- Hedia, S.I.; Christopher, S.; Zhebentyayeva, T.; Ledbetter, C.; Krška, B.; Remay, A.; D’Onofrio, C.; Iketani, H.; Christen, D.; Krichen, L.; et al. Genetic Structure of a Worldwide Germplasm Collection of Prunus armeniaca L. Reveals Three Major Diffusion Routes for Varieties Coming from the Species’ Center of Origin. Bourguiba. Front. Plant Sci. 2020, 11, 638. [Google Scholar]
- Karatas, N. Evaluation of Nutritional Content in Wild Apricot Fruits for Sustainable Apricot Production. Sustainability 2022, 14, 1063. [Google Scholar] [CrossRef]
- Cirillo, A.; De Luca, L.; Izzo, L.; Cepparulo, M.; Graziani, G.; Ritieni, A.; Romano, R.; Di Vaio, C. Biochemical and nutraceutical characterization of different accessions of the apricot (Prunus armeniaca L.). Horticulturae 2023, 9, 546. [Google Scholar] [CrossRef]
- Wang, X.; Wang, L.; Sun, Y.; Chen, J.; Liu, Q.; Dong, S. Genetic diversity and conservation of Siberian apricot (Prunus sibirica L.) based on microsatellite markers. Sci. Rep. 2023, 13, 11245. [Google Scholar] [CrossRef]
- Chen, L.; Sheng, Q.; Liu, J.; Jia, Y.; Zhao, W.; Li, P.; Zhang, A. A Review on the Functional Properties, Extraction and Microencapsulation of Almond Oil. Sci. Technol. Food Ind. 2023. [Google Scholar] [CrossRef]
- Daniell, H.; Jin, S.; Zhu, X.; Gitzendanner, M.A.; Soltis, D.E.; Soltis, P.S. Green giant—A tiny chloroplast genome with mighty power to produce high-value proteins: History and phylogeny. Plant Biotechnol. J. 2021, 19, 430–447. [Google Scholar] [CrossRef]
- Jiang, H.; Tian, J.; Yang, J.; Dong, X.; Zhong, Z.; Mwachala, G.; Zhang, C.; Hu, G.; Wang, Q. Comparative and phylogenetic analyses of six Kenya Polystachya (Orchidaceae) species based on the complete chloroplast genome sequences. BMC Plant Biol. 2022, 22, 177. [Google Scholar] [CrossRef] [PubMed]
- Xiao, W.; Fei, S.; Qiuping, Z.; Liu, N.; Zhang, Y.; Xu, M.; Liu, S.; Zhang, Y.; Ma, X.; Liu, W. Genetic diversity analysis of Chinese plum (Prunus salicina L.) based on whole-genome resequencing. Tree Genet. Genomes 2021, 17, 26. [Google Scholar]
- Oh, S.H.; Park, J. The complete chloroplast genome of Euscaphis japonica (Thunb.) Kanitz (Staphyleaceae) isolated in Korea. Mitochondr. DNA B Resour. 2020, 5, 3769–3771. [Google Scholar] [CrossRef] [PubMed]
- Zha, X.; Wang, X.; Li, J.; Gao, F.; Zhou, Y. Complete chloroplast genome of Sophora alopecuroides (Papilionoideae): Molecular structures, comparative genome analysis and phylogenetic analysis. J. Genet. 2020, 99, 1–14. [Google Scholar] [CrossRef]
- Du, X.; Zeng, T.; Feng, Q.; Hu, L.; Luo, X.; Weng, Q.; He, J.; Zhu, B. The complete chloroplast genome sequence of yellow mustard (Sinapis alba L.) and its phylogenetic relationship to other Brassicaceae species. Gene 2020, 731, 144340. [Google Scholar] [CrossRef] [PubMed]
- Bai, G.Q.; Zhou, T.; Zhao, J.X.; Li, W.M.; Han, G.J.; Li, S.F. The complete chloroplast genome of Kolkwitzia amabilis (Caprifoliaceae), an endangered horticultural plant in China. Mitochondr. DNA Part A 2017, 28, 296–297. [Google Scholar] [CrossRef]
- Shinozaki, K.; Ohme, M.; Tanaka, M.; Wakasugi, T.; Hayashida, N.; Matsubayashi, T.; Zaita, N.; Chunwongse, J.; Obokata, J.; Yamaguchi-Shinozaki, K.; et al. The complete nucleotide sequence of the tobacco chloroplast genome: Its gene organization and expression. EMBO J. 1986, 5, 2043–2049. [Google Scholar] [CrossRef]
- Song, Y.F.; Yang, Q.H.; Yi, X.G.; Zhu, Z.Q.; Wang, X.R.; Li, M. Comparative analysis of codon usage patterns in chloroplast genomes of cherries. Forests 2022, 13, 1891. [Google Scholar] [CrossRef]
- Wu, J.Q.; Wang, Y.; Sun, P.; Sun, Z.S.; Shen, J.S. The complete chloroplast genome of Prunus phaeosticta (Hance) Maxim. (Rosaceae) and its phylogenetic implications. Mitochondr. DNA B 2023, 8, 136–140. [Google Scholar] [CrossRef]
- Zhongyu, D.; Ke, L.; Kai, Z.; He, Y.; Wang, H.; Chai, G.; Shi, J.; Duan, Y. The chloroplast genome of Amygdalus L. (Rosaceae) reveals the phylogenetic relationship and divergence time. BMC Genom. 2021, 22, 645. [Google Scholar]
- Mu, J.; Zhao, Y.; He, Y.; Sun, J.; Yuan, Q. The complete chloroplast genome of Prunus japonica thunb. (Rosaceae), an ornamental and medicinal plant. Mitochondr. DNA B 2021, 6, 112–114. [Google Scholar] [CrossRef] [PubMed]
- Hoang, T.K.; Wang, Y.; Hwang, Y.-J.; Lim, J.-H. Analysis of the morphological characteristics and karyomorphology of wild Chrysanthemum species in Korea. Hortic. Environ. Biotechnol. 2020, 61, 359–369. [Google Scholar] [CrossRef]
- Wani, A.A.; Zargar, S.A.; Malik, A.H.; Kashtwari, M.; Nazir, M.; Khuroo, A.A.; Ahmad, F.; Dar, T.A. Assessment of variability in morphological characters of apricot germplasm of Kashmir, India. Sci. Hortic. 2017, 225, 630–637. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhang, X.; Yan, S.; Feng, C.; Wang, D.; Yang, W.; Daud, M.K.; Xiang, J.; Mei, L. SSR identification and phylogenetic analysis in four plant species based on complete chloroplast genome sequences. Plasmid 2023, 125, 102670. [Google Scholar] [CrossRef]
- Alzahrani, D.; Albokhari, E.; Abba, A.; Yaradua, S. The first complete chloroplast genome sequences in Resedaceae: Genome structure and comparative analysis. Sci. Prog. 2021, 104, 368504211059973. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Zhu, J.; Ma, X.; Jia, J.; Luo, D.; Ding, Q.; Wang, X.; Huang, L. Comparative analysis of switchgrass chloroplast genomes provides insights into identification, phylogenetic relationships and evolution of different ecotypes. Ind. Crops Prod. 2023, 205, 117570. [Google Scholar] [CrossRef]
- Wang, S.; Gao, C.W.; Gao, L.Z. Plastid genome sequence of an ornamental and editable fruit tree of Rosaceae, Prunus mume. Mitochondr. DNA Part A 2016, 27, 4407–4408. [Google Scholar] [CrossRef]
- Terakami, S.; Matsumura, Y.; Kurita, K.; Kanamori, H.; Katayose, Y.; Yamamoto, T.; Katayama, H. Complete sequence of the chloroplast genome from pear (Pyrus pyrifolia): Genome structure and comparative analysis. Tree Genet. Genomes 2012, 8, 841–854. [Google Scholar] [CrossRef]
- Lu, R.S.; Li, P.; Qiu, Y.X. The complete chloroplast genomes of three Cardiocrinum (Liliaceae) species: Comparative genomic and phylogenetic analyses. Front. Plant Sci. 2016, 7, 2054. [Google Scholar] [CrossRef] [PubMed]
- Daniell, H.; Lin, C.S.; Yu, M.; Chang, W.J. Chloroplast genomes: Diversity, evolution, and applications in genetic engineering. Genome Biol. 2016, 17, 134. [Google Scholar] [CrossRef]
- Wang, W.C.; Chen, S.Y.; Zhang, X.Z. Chloroplast Genome Evolution in Actinidiaceae: clpP Loss, Heterogenous Divergence and Phylogenomic Practice. PLoS ONE 2016, 11, e0162324. [Google Scholar] [CrossRef]
- Asaf, S.; Khan, A.L.; Khan, M.A.; Waqas, M.; Kang, S.M.; Yun, B.W.; Lee, I.J. Chloroplast genomes of Arabidopsis halleri ssp. gemmifera and Arabidopsis lyrata ssp. petraea: Structures and comparative analysis. Sci. Rep. 2017, 7, 7556. [Google Scholar]
- Zhang, Y.; Du, L.; Liu, A.; Chen, J.; Wu, L.; Hu, W.; Zhang, W.; Kim, K.; Lee, S.-C.; Yang, T.-J.; et al. The complete chloroplast genome sequences of five Epimedium species: Lights into phylogenetic and taxonomic analyses. Front. Plant Sci. 2016, 7, 306. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Li, J.; Yu, L.; Bai, W.; Nie, D.; Xiong, Y.; Wu, S. Comparative chloroplast genomes of Prunus subgenus Cerasus (Rosaceae): Insights into sequence variations and phylogenetic relationships. Tree Genet. Genomes 2021, 17, 50. [Google Scholar] [CrossRef]
- Jansen, R.K.; Saski, C.; Lee, S.B.; Hansen, A.K.; Daniell, H. Complete plastid genome sequences of three Rosids (Castanea, Prunus, Theobroma): Evidence for at least two independent transfers of rpl22 to the nucleus. KRJ Mol. Biol. Evol. 2011, 28, 835–847. [Google Scholar] [CrossRef] [PubMed]
- Nan, W.; Ming, C.; Yong, C.; Liu, B.; Wang, X.; Li, G.; Zhou, Y.; Luo, P.; Xi, Z.; Yong, H.; et al. Natural variations in the non-coding region of ZmNAC080308 contributes maintaining grain yield under drought stress in maize. BMC Plant Biol. 2021, 21, 305. [Google Scholar]
- Lin, C.P.; Wu, C.S.; Huang, Y.Y.; Chaw, S.M. The complete chloroplast genome of Ginkgo biloba reveals the mechanism of inverted repeat contraction. Genome Biol. Evol. 2012, 4, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Chen, X.; Geng, L.; Tang, C.; Wei, X. Phylogenetic Analysis of Sorbus ser Folgnerianae (Rosaceae). J. Nanjing For. Univ. 2023, pp. 1–15. Available online: https://kns.cnki.net/kcms/detail/32.1161.S.20230412.1754.006.html (accessed on 13 September 2023).
- Xue, S.; Shi, T.; Luo, W.; Ni, X.; Iqbal, S.; Ni, Z.; Huang, X.; Yao, D.; Shen, Z.; Gao, Z. Comparative analysis of the complete chloroplast genome among Prunus mume, P. Armeniaca, and P. salicina. Hortic. Res. 2019, 6, 89. [Google Scholar] [CrossRef]
- Jansen, R.K.; Cai, Z.; Raubeson, L.A.; Daniell, H.; Depamphilis, C.W.; Leebens-Mack, J.; Müller, K.F.; Guisinger-Bellian, M.; Haberle, R.C.; Hansen, A.K.; et al. Analysis of 81 genes from 64 plastid genomes resolves relationships in angiosperms and identifies genome-scale evolutionary patterns. Proc. Natl. Acad. Sci. USA 2007, 104, 19369–19374. [Google Scholar] [CrossRef] [PubMed]
- Raman, G.; Park, S. The complete chloroplast genome sequence of Ampelopsis: Gene organization, comparative analysis and phylogenetic relationships to other angiosperms. Front. Plant Sci. 2016, 7, 341. [Google Scholar] [CrossRef] [PubMed]
- Angiosperm Phylogeny Group. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Bot. J. Linn. Soc. 2009, 161, 105–121. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, W.; Liu, W.; Liu, N.; Zhang, Y.; Liu, S. Palynological Study on the Origin and Systematic Evolution of Kernel-using Apricots. Acta Hortic. Sin. 2010, 37, 1377–1387. [Google Scholar]
- Li, M.; Zheng, P.; Ni, B.; Hu, X.; Miao, X.; Zhao, Z. Genetic diversity analysis of apricot cultivars grown in China based on SSR markers. Eur. J. Hortic. Sci. 2018, 83, 18–27. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | P. armeniaca | Kernel Consumption Apricot | P. sibirica |
|---|---|---|---|
| Size, bp | 157,951 | 157,994 | 158,224 |
| LSC 1, bp (%) | 86,217 (54.58) | 86,244 (54.59) | 86,358 (54.58) |
| SSC, bp (%) | 18,988 (12.02) | 19,004 (12.03) | 19,120 (12.09) |
| IR, bp (%) | 26,373 (16.70) | 26,373 (16.69) | 26,373 (16.67) |
| Protein-coding regions, bp (%) | 78,530 (49.72) | 78,532 (49.71) | 78,532 (49.63) |
| Introns, bp (%) | 18,017 (11.41) | 18,005 (11.40) | 18,049 (11.41) |
| rRNA, bp (%) | 9,048 (5.73) | 9,048 (5.73) | 9,048 (5.72) |
| tRNA, bp (%) | 2,804 (1.78) | 2,804 (1.78) | 2,806 (1.77) |
| IGS, bp (%) | 49,552 (31.34) | 49,605 (31.40) | 49,789 (31.47) |
| Total GC content (%) | 36.74 | 36.75 | 36.7 |
| LSC GC content (%) | 34.55 | 34.56 | 34.52 |
| SSC GC content (%) | 30.5 | 30.46 | 30.3 |
| IR GC content (%) | 42.58 | 42.59 | 42.57 |
| Protein-coding regions’ GC content (%) | 37.63 | 37.63 | 37.64 |
| Introns GC content (%) | 36.68 | 36.68 | 36.61 |
| IGS GC content (%) | 30.77 | 30.83 | 30.65 |
| rRNA GC content (%) | 55.5 | 55.5 | 55.5 |
| tRNA GC content (%) | 53.35 | 53.25 | 53.28 |
| Total number of genes | 131 | 131 | 131 |
| Protein-coding genes | 86 | 86 | 86 |
| tRNA genes | 37 | 37 | 37 |
| rRNA genes | 8 | 8 | 8 |
| GenBank accession numbers | KY101151 | KY101150 | KY101154 |
| Gene Categories | Gene Groups | Gene Names |
|---|---|---|
| Self-replication | Ribosomal RNAs | rrn16 b, rrn23 b, rrn4.5 b, rrn5 b |
| Transfer RNAs | trnA-UGC a,b, trnC-GCA, trnD-GUC, trnE-UUC, trnF-GAA, trnfM-CAU, trnG-GCC a, trnH-GUG, trnI-CAU b, trnI-GAU a,b, trnK-UUU a, trnL-CAA b, trnL-UAA a, trnL-UAG, trnM-CAU, trnN-GUU b, trnP-UGG, trnQ-UUG, trnR-ACG b, trnR-UCU, trnS-GCU, trnS-GGA, trnS-UGA, trnT-GGU, trnT-UGU, trnV-GAC b, trnV-UAC a, trnW-CCA, trnY-GUA | |
| Small ribosomal subunit | rps2, rps3, rps4, rps7 b, rps8, rps11, rps12 a,b, rps14, rps15, rps16 a, rps18, rps19 b | |
| Large ribosomal subunit | rpl2 a,b, rpl14, rpl16 a, rpl20, rpl22, rpl23 b, rpl32, rpl33, rpl36 | |
| RNA polymerase | rpoA, rpoB, rpoC1 a, rpoC2 | |
| Photosynthesis | NADH-dehydrogenase | ndhA a, ndhB a,b, ndhC, ndhD, ndhE, ndhF, ndhG, ndhH, ndhI, ndhJ, ndhK |
| Photosystem Ⅰ | psaA, psaB, psaC, psaI, psaJ | |
| Photosystem Ⅱ | psbA, psbB, psbC, psbD, psbE, psbF, psbH, psbI, psbJ, psbK, psbL, psbM, psbN, psbT, psbZ | |
| RuBisCO c | rbcL | |
| Cytochrome b/f complex | petA, petB a, petD a, petG, petL, petN | |
| ATP synthase | atpA, atpB, atpE, atpF a, atpH, atpI | |
| Other | Acetyl-CoA carboxylase | accD |
| Cytochrome c biogenesis | ccsA | |
| Maturase | matK | |
| Envelope membrane protein | cemA | |
| Unknown | Conserved hypothetical reading frames | ycf1 b, ycf2 b, ycf3 a, ycf4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yi, R.; Bao, W.; Ao, D.; Bai, Y.-e.; Wang, L.; Wuyun, T.-n. Sequencing and Phylogenetic Analysis of the Chloroplast Genome of Three Apricot Species. Genes 2023, 14, 1959. https://doi.org/10.3390/genes14101959
Yi R, Bao W, Ao D, Bai Y-e, Wang L, Wuyun T-n. Sequencing and Phylogenetic Analysis of the Chloroplast Genome of Three Apricot Species. Genes. 2023; 14(10):1959. https://doi.org/10.3390/genes14101959
Chicago/Turabian StyleYi, Ru, Wenquan Bao, Dun Ao, Yu-e Bai, Lin Wang, and Ta-na Wuyun. 2023. "Sequencing and Phylogenetic Analysis of the Chloroplast Genome of Three Apricot Species" Genes 14, no. 10: 1959. https://doi.org/10.3390/genes14101959
APA StyleYi, R., Bao, W., Ao, D., Bai, Y.-e., Wang, L., & Wuyun, T.-n. (2023). Sequencing and Phylogenetic Analysis of the Chloroplast Genome of Three Apricot Species. Genes, 14(10), 1959. https://doi.org/10.3390/genes14101959