

Ozone Disinfection for Elimination of Bacteria and Degradation of SARS-CoV2 RNA for Medical Environments

 ,
, 
Abstract
1. Introduction
2. Methods
2.1. Bacterial Strains
2.2. Ozone Treatment of Bacteria
2.3. E. coli K12 Viability Based on ATP Presence
2.4. RNA Sequencing
2.5. Differential Gene Expression and Gene Ontology Analysis
2.6. Ozone Treatment of Synthetic SARS-CoV-2 RNA
2.7. Ozone Treatment of Non-Replicative Virus
2.8. Statistical Analysis
3. Results
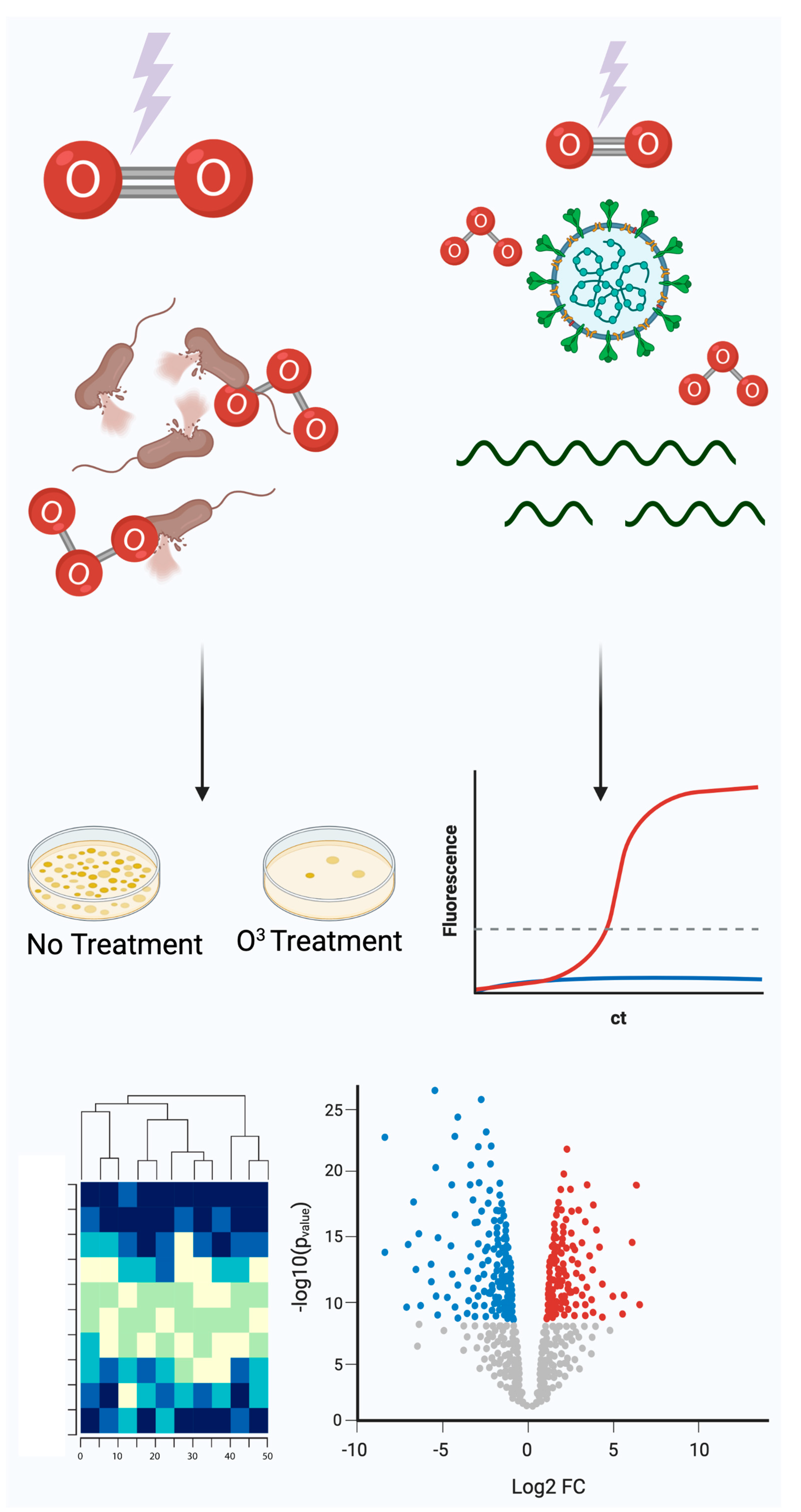
3.1. Bacteria Susceptibility to Ozone Treatment
3.2. Gene Expression Profiling of Ozone Treatment
3.3. SARS-CoV-2 RNA Susceptibility to Ozone Treatment
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Reardon, S. WHO warns against ‘post-antibiotic’ era. Nature 2014, 244, 1356–1357. [Google Scholar] [CrossRef]
- Read, A.F.; Woods, R.J. Antibiotic resistance management. Evol. Med. Public Health 2014, 2014, 147. [Google Scholar] [CrossRef] [PubMed]
- Zaman, S.B.; Hussain, M.A.; Nye, R.; Mehta, V.; Mamun, K.T.; Hossain, N. A Review on Antibiotic Resistance: Alarm Bells are Ringing. Cureus 2017, 9, e1403. [Google Scholar] [CrossRef] [PubMed]
- Lüddeke, F.; Heß, S.; Gallert, C.; Winter, J.; Güde, H.; Löffler, H. Removal of total and antibiotic resistant bacteria in advanced wastewater treatment by ozonation in combination with different filtering techniques. Water Res. 2015, 69, 243–251. [Google Scholar] [CrossRef]
- Dyas, A.; Boughton, B.J.; Das, B.C. Ozone killing action against bacterial and fungal species; microbiological testing of a domestic ozone generator. J. Clin. Pathol. 1983, 36, 1102–1104. [Google Scholar] [CrossRef]
- Environmental Protection Agency (EPA). EPA Guidance Manual, Alternative Disinfectants and Oxidants, Ozone Chemistry. April 1999, Chapter 3.1. Available online: www.epa.gov (accessed on 6 December 2022).
- National Research Council (US) Committee on Toxicology. Emergency and Continuous Exposure Limits for Selected Airborne Contaminants; National Academies Press: Washington, DC, USA, 1984; Volume 1. [Google Scholar]
- Kim, S.Y.; Kim, E.; Kim, W.J. Health Effects of Ozone on Respiratory Diseases. Tuberc. Respir. Dis. 2020, 83 (Suppl. 1), S6–S11. [Google Scholar] [CrossRef]
- Kramer, T.S.; Remschmidt, C.; Werner, S.; Behnke, M.; Schwab, F.; Werner, G.; Gastmeier, P.; Leistner, R. The importance of adjusting for enterococcus species when assessing the burden of vancomycin resistance: A cohort study including over 1000 cases of enterococcal bloodstream infections. Antimicrob. Resist. Infect. Control. 2018, 7, 133. [Google Scholar] [CrossRef]
- Salmanov, A.; Vozianov, S.; Kryzhevsky, V.; Litus, O.; Drozdova, A.; Vlasenko, I. Prevalence of healthcare-associated infections and antimicrobial resistance in acute care hospitals in Kyiv, Ukraine. J. Hosp. Infect. 2019, 102, 431–437. [Google Scholar] [CrossRef]
- Krisko, A.; Radman, M. Biology of extreme radiation resistance: The way of Deinococcus radiodurans. Cold Spring Harb. Perspect. Biol. 2013, 5, a012765. [Google Scholar] [CrossRef]
- Young, S.; Setlow, P. Mechanisms of Bacillus subtilis spore resistance to and killing by aqueous ozone. J. Appl. Microbiol. 2004, 96, 1133–1142. [Google Scholar] [CrossRef]
- Czekalski, N.; Imminger, S.; Salhi, E.; Veljkovic, M.; Kleffel, K.; Drissner, D.; Hammes, F.; Bürgmann, H.; von Gunten, U. Inactivation of Antibiotic Resistant Bacteria and Resistance Genes by Ozone: From Laboratory Experiments to Full-Scale Wastewater Treatment. Environ. Sci. Technol. 2016, 50, 11862–11871. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, R.P.; Kirit, H.A.; Predeus, A.V.; Bollback, J.P. Transcriptomic profiling of Escherichia coli K-12 in response to a compendium of stressors. Sci. Rep. 2022, 12, 8788. [Google Scholar] [CrossRef] [PubMed]
- Dennis, R.; Pourdeyhimi, B.; Cashion, A.; Emanuel, S.; Hubbard, D. Durability of Disposable N95 Mask Material When Exposed to Improvised Ozone Gas Disinfection. J. Sci. Med. 2020, 2. [Google Scholar] [CrossRef]
- Blanchard, E.L.; Lawrence, J.D.; Noble, J.A.; Xu, M.; Joo, T.; Ng, N.L.; Schmidt, B.E.; Santangelo, P.J.; Finn, M.G. Enveloped Virus Inactivation on Personal Protective Equipment by Exposure to Ozone. MedRxiv 2020. [Google Scholar] [CrossRef]
- Hudson, J.B.; Sharma, M.; Vimalanathan, S. Development of a Practical Method for Using Ozone Gas as a Virus Decontaminating Agent. Ozone Sci. Eng. 2009, 31, 216–223. [Google Scholar] [CrossRef]
- Schentag, J.J.; Akers, C.; Campagna, P.; Chirayath, P. SARS: CLEARING THE AIR. In Institute of Medicine (US) Forum on Microbial Threats; Knobler, S., Mahmoud, A., Lemon, S., Mack, A., Sivitz, L., Oberholtzer, K., Eds.; Learning from SARS: Preparing for the Next Disease Outbreak: Workshop Summary; National Academies Press (US): Washington, DC, USA, 2004. Available online: https://www.ncbi.nlm.nih.gov/books/NBK92445/ (accessed on 18 November 2021).
- Roy, D.; Wong, P.K.; Engelbrecht, R.S.; Chian, E.S. Mechanism of enteroviral inactivation by ozone. Appl. Environ. Microbiol. 1981, 41, 718–723. [Google Scholar] [CrossRef]
- Shinriki, N.; Ishizaki, K.; Ikehata, A.; Nomura, A.; Mizuno, Y. Degradation of nucleic acid with ozone. Nucleic Acids Res. 1978, 5 (Suppl. 2), s303–s308. [Google Scholar] [CrossRef]
- Gingrich, J.; Rubio, T.; Karlak, C. Effect of RNA Degradation on the Data Quality in Quantitative PCR and MICROARRAY Experiments; Bio-Rad Bulletin: Hercules, CA, USA, 2006. [Google Scholar]
- Butler, D.; Mozsary, C.; Meydan, C.; Foox, J.; Rosiene, J.; Shaiber, A.; Danko, D.; Afshinnekoo, E.; MacKay, M.; Sedlazeck, F.J.; et al. Shotgun transcriptome, spatial omics, and isothermal profiling of SARS-CoV-2 infection reveals unique host responses, viral diversification, and drug interactions. Nat. Commun. 2021, 12, 1660. [Google Scholar] [CrossRef]
- Wade, W. Unculturable bacteria--the uncharacterized organisms that cause oral infections. J. R. Soc. Med. 2002, 95, 81–83. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; 1000 Genome Project Data Processing Subgroup. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Alexa, A.; Rahnenfuhrer, J. topGO: Enrichment Analysis for Gene Ontology; R Package Version 2.40.0. 2020. Available online: https://bioconductor.org/packages/release/bioc/html/topGO.html (accessed on 5 December 2022).
- Zhao, X.; Drlica, K. Reactive oxygen species and the bacterial response to lethal stress. Curr. Opin. Microbiol. 2014, 21, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Dukan, S.; Touati, D. Hypochlorous acid stress in Escherichia coli: Resistance, DNA damage, and comparison with hydrogen peroxide stress. J. Bacteriol. 1996, 178, 6145–6150. [Google Scholar] [CrossRef] [PubMed]
- Roncarati, D.; Scarlato, V. Regulation of heat-shock genes in bacteria: From signal sensing to gene expression output. FEMS Microbiol. Rev. 2017, 41, 549–574. [Google Scholar] [CrossRef]
- Lau, C.K.; Krewulak, K.D.; Vogel, H.J. Bacterial ferrous iron transport: The Feo system. FEMS Microbiol. Rev. 2016, 40, 273–298. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, D.E.; Schultz, J.E.; Matin, A. Starvation-induced cross protection against heat or H2O2 challenge in Escherichia coli. J. Bacteriol. 1988, 170, 3910–3914. [Google Scholar] [CrossRef]
- Lindqvist, A.; Membrillo-Hernańdez, J.; Poole, R.K.; Cook, G.M. Roles of respiratory oxidases in protecting Escherichia coli K12 from oxidative stress. Antonie Van Leeuwenhoek 2000, 78, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.; O’Sullivan, D.; Pereira, R.; Whale, A.; Busby, E.; Huggett, J.; Harris, K. Comparison of SARS-CoV2 N gene real-time RT-PCR targets and commercially available mastermixes. BioRxiv 2020. [Google Scholar] [CrossRef]
- Corman, V.M.; Landt, O.; Kaiser, M.; Molenkamp, R.; Meijer, A.; Chu, D.K.; Bleicker, T.; Brünink, S.; Schneider, J.; Schmidt, M.L.; et al. Detection of 2019 novel coronavirus (2019-nCoV) by real-time RT-PCR. Euro Surveill. Bull. Eur. Sur Les Mal. Transm.=Eur. Commun. Dis. Bull. 2020, 25, 2000045. [Google Scholar] [CrossRef]
- Rangel, K.; Cabral, F.O.; Lechuga, G.C.; Carvalho, J.; Villas-Bôas, M.; Midlej, V.; De-Simone, S.G. Detrimental Effect of Ozone on Pathogenic Bacteria. Microorganisms 2021, 10, 40. [Google Scholar] [CrossRef]
- Bayarri, B.; Cruz-Alcalde, A.; López-Vinent, N.; Micó, M.M.; Sans, C. Can ozone inactivate SARS-CoV-2? A review of mechanisms and performance on viruses. J. Hazard. Mater. 2021, 415, 125658. [Google Scholar] [CrossRef]
- Tizaoui, C.; Stanton, R.; Statkute, E.; Rubina, A.; Lester-Card, E.; Lewis, A.; Holliman, P.; Worsley, D. Ozone for SARS-CoV-2 inactivation on surfaces and in liquid cell culture media. J. Hazard. Mater. 2022, 428, 128251. [Google Scholar] [CrossRef] [PubMed]
- Osha; Occupational Safety and Health Administration; U.S. Department of Health and Human Services. 1992. Available online: http://www.osha.gov/dts/chemicalsampling/data/CH_259300.html (accessed on 20 May 2022).
- Holm, S.M.; Balmes, J.R. Systematic Review of Ozone Effects on Human Lung Function, 2013 Through 2020. Chest 2022, 161, 190–201. [Google Scholar] [CrossRef] [PubMed]
- Emerson, J.B.; Adams, R.I.; Rothschild, L.J. Schrodinger’s microbes: Tools for distinguishing the living from dead in microbial ecosystems. Microbiome 2017, 5, 86. [Google Scholar] [CrossRef] [PubMed]
- Battista, J.R. Against all odds: The survival strategies of Deinococcus radiodurans. Annu. Rev. Microbiol. 1997, 51, 203–224. [Google Scholar] [CrossRef] [PubMed]
- Foerster, S.; Unemo, M.; Hathaway, L.J.; Low, N.; Althaus, C.L. Time-kill curve analysis and pharmacodynamic functions for in vitro evaluation of antimicrobials against Neisseria gonorrhoeae. BMC Microbiol. 2015, 16, 216. [Google Scholar] [CrossRef]
- Fontes, B.; Heimbecker, A.M.; Brito, G.D.; Costa, S.F.; Heijden, I.M.; Levin, A.S.; Rasslan, S. Effect of low-dose gaseous ozone on pathogenic bacteria. BMC Infect. Dis. 2012, 12, 358. [Google Scholar] [CrossRef]
- Shah, D.H. RNA Sequencing Reveals Differences between the Global Transcriptomes of Salmonella enterica Serovar Enteritidis Strains with High and Low Pathogenicities. Appl. Environ. Microbiol. 2013, 80, 896–906. [Google Scholar] [CrossRef]
- O’Hara, N.B.; Reed, H.J.; Afshinnekoo, E.; Harvin, D.; Caplan, N.; Rosen, G.; Frye, B.; Woloszynek, S.; Ounit, R.; Levy, S.; et al. Metagenomic characterization of ambulances across the United States. Microbiome 2017, 5, 125. [Google Scholar] [CrossRef]
- The MetaSUB International Consortium. The Metagenomics and Metadesign of the Subways and Urban Biomes (MetaSUB) International Consortium inaugural meeting report. Microbiome 2016, 4, 24. [Google Scholar] [CrossRef]
- Tizaoui, C. Ozone: A potential oxidant for COVID-19 virus (SARS-CoV-2). Ozone Sci. Eng. 2020, 42, 378–385. [Google Scholar] [CrossRef]
- Mendoza, E.J.; Manguiat, K.; Wood, H.; Drebot, M. Two Detailed Plaque Assay Protocols for the Quantification of Infectious SARS-CoV-2. Curr. Protoc. Microbiol. 2020, 57, ecpmc105. [Google Scholar] [CrossRef] [PubMed]
- Kitagawa, H.; Nomura, T.; Nazmul, T.; Omori, K.; Shigemoto, N.; Sakaguchi, T.; Ohge, H. Effectiveness of 222-nm ultraviolet light on disinfecting SARS-CoV-2 surface contamination. Am. J. Infect. Control. 2020, 49, 299–301. [Google Scholar] [CrossRef] [PubMed]
- Torres-Mata, L.B.; García-Pérez, O.; Rodríguez-Esparragón, F.; Blanco, A.; Villar, J.; Ruiz-Apodaca, F.; Martín-Barrasa, J.L.; González-Martín, J.M.; Serrano-Aguilar, P.; Piñero, J.E.; et al. Ozone Eliminates SARS-CoV-2 from Difficult-to-Clean Office Supplies and Clinical Equipment. Int. J. Environ. Res. Public Health 2022, 19, 8672. [Google Scholar] [CrossRef]
- Chin AW, H.; Chu JT, S.; Perera MR, A.; Hui KP, Y.; Yen, H.L.; Chan MC, W.; Peiris, M.; Poon LL, M. Stability of SARS-CoV-2 in different environmental conditions. Lancet. Microbe 2020, 1, e10. [Google Scholar] [CrossRef]
- van Doremalen, N.; Bushmaker, T.; Morris, D.H.; Holbrook, M.G.; Gamble, A.; Williamson, B.N.; Tamin, A.; Harcourt, J.L.; Thornburg, N.J.; Gerber, S.I.; et al. Aerosol and Surface Stability of SARS-CoV-2 as Compared with SARS-CoV-1. N. Engl. J. Med. 2020, 382, 1564–1567. [Google Scholar] [CrossRef]
- Carraturo, F.; Del Giudice, C.; Morelli, M.; Cerullo, V.; Libralato, G.; Galdiero, E.; Guida, M. Persistence of SARS-CoV-2 in the environment and COVID-19 transmission risk from environmental matrices and surfaces. Environ. Pollut. Barking Essex 1987 2020, 265 Pt B, 115010. [Google Scholar] [CrossRef]
- Brown, J.; O’Sullivan, D.; Pereira, R.; Whale, A.; Busby, E.; Huggett, J.; Harris, K. Comparison of SARS-CoV2 N gene real-time RT-PCR targets and commercially available mastermixes. J. Virol. Methods 2021, 295, 114215. [Google Scholar] [CrossRef]
- Solo-Gabriele, H.M.; Kumar, S.; Abelson, S.; Penso, J.; Contreras, J.; Babler, K.M.; Sharkey, M.E.; Mantero AM, A.; Lamar, W.E.; Tallon, J.J.; et al. Predicting COVID-19 cases using SARS-CoV-2 RNA in air, surface swab and wastewater samples. Sci. Total Environ. 2023, 857 Pt 1, 159188. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GO.ID | Term | Rank in Class | Classic Fisher | Classic KS |
|---|---|---|---|---|
| GO:0055114 | oxidation-reduction process | 1 | 8.7 × 109 | 5.9 × 1011 |
| GO:0045333 | cellular respiration | 2 | 5.7 × 107 | 8.4 × 109 |
| GO:0009060 | aerobic respiration | 3 | 1.8 × 106 | 2.2 × 108 |
| GO:0015980 | energy derivation by oxidation of organisms | 4 | 2.8 × 106 | 1.1 × 107 |
| GO:0006099 | tricarboxylic acid cycle | 5 | 8.2 × 106 | 1.7 × 106 |
| GO:0006101 | citrate metabolic process | 6 | 8.2 × 106 | 1.7 × 106 |
| GO:0072350 | tricarboxylic acid metabolic process | 7 | 8.2 × 106 | 1.7 × 106 |
| GO:0006091 | generation of precursor metabolites | 8 | 2.9 × 105 | 2.5 × 106 |
| GO:0072329 | monocarboxylic acid catabolic process | 9 | 5.6 × 105 | 3.0 × 105 |
| GO:0016999 | antibiotic metabolic process | 10 | 6.5 × 105 | 1.3 × 105 |
| Synthetic RNA | Ct Mean | Ct SD | △△CT Treatment-Control | % RNA Fold Change vs. Control |
| 0 h Control | 18.22 | 0.48 | 0.00 | 100.0 |
| 30 min air exposure | 18.24 | 0.37 | 0.02 | 98.7 |
| 1 h air exposure | 18.32 | 0.07 | 0.10 | 93.5 |
| 2 h air exposure | 18.43 | 0.29 | 0.21 | 86.2 |
| 3 h air exposure | 19.99 | 0.42 | 1.77 | 29.3 |
| 4 h air exposure | 20.56 | 0.12 | 2.34 | 19.7 |
| 30 min O3 | 18.68 | 0.16 | 0.46 | 72.5 |
| 1 h O3 | 18.79 | 0.14 | 0.57 | 67.2 |
| 2 h O3 | 21.37 | 0.13 | 3.15 | 11.2 |
| 3 h O3 | 21.22 | 0.44 | 3.00 | 12.5 |
| 4 h O3 | 22.24 | 0.78 | 4.02 | 6.2 |
| Rnase | 38.81 | 3.01 | 20.59 | 0.0 |
| 10,000 copies of Capsid | Ct Mean | Ct SD | △△CT Treatment-Control | % RNA fold change vs. control |
| 0 h Control | 25.26 | 0.55 | 0.00 | 100.00 |
| 30 min O3 | 27.13 | 0.87 | 1.86 | 27.53 |
| 1 h O3 | 27.81 | 2.74 | 2.55 | 17.10 |
| 2 h O3 | 29.31 | 0.75 | 4.04 | 6.07 |
| 3 h O3 | 29.08 | 2.49 | 3.82 | 7.09 |
| 4 h O3 | 28.54 | 0.67 | 3.28 | 10.33 |
| H2O2 | 27.57 | 0.53 | 2.30 | 20.29 |
| Rnase | 41.40 | 0.73 | 16.13 | 0.00 |
| 1000 copies of Capsid | Ct Mean | Ct SD | △△CT Treatment-Control | % RNA fold change vs. control |
| 0 h Control | 28.46 | 0.40 | 0.00 | 100.00 |
| 30 min O3 | 30.05 | 0.05 | 1.59 | 33.17 |
| 1 h O3 | 32.53 | 0.37 | 4.07 | 5.95 |
| 2 h O3 | 32.73 | 0.36 | 4.27 | 5.19 |
| 3 h O3 | 32.77 | 1.30 | 4.31 | 5.04 |
| 4 h O3 | 33.95 | 0.51 | 5.50 | 2.22 |
| H2O2 | 30.27 | 0.45 | 1.81 | 28.45 |
| Rnase | 41.40 | 0.73 | 12.94 | 0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Westover, C.; Rahmatulloev, S.; Danko, D.; Afshin, E.E.; O’Hara, N.B.; Ounit, R.; Bezdan, D.; Mason, C.E. Ozone Disinfection for Elimination of Bacteria and Degradation of SARS-CoV2 RNA for Medical Environments. Genes 2023, 14, 85. https://doi.org/10.3390/genes14010085
Westover C, Rahmatulloev S, Danko D, Afshin EE, O’Hara NB, Ounit R, Bezdan D, Mason CE. Ozone Disinfection for Elimination of Bacteria and Degradation of SARS-CoV2 RNA for Medical Environments. Genes. 2023; 14(1):85. https://doi.org/10.3390/genes14010085
Chicago/Turabian StyleWestover, Craig, Savlatjon Rahmatulloev, David Danko, Evan E. Afshin, Niamh B. O’Hara, Rachid Ounit, Daniela Bezdan, and Christopher E. Mason. 2023. "Ozone Disinfection for Elimination of Bacteria and Degradation of SARS-CoV2 RNA for Medical Environments" Genes 14, no. 1: 85. https://doi.org/10.3390/genes14010085
APA StyleWestover, C., Rahmatulloev, S., Danko, D., Afshin, E. E., O’Hara, N. B., Ounit, R., Bezdan, D., & Mason, C. E. (2023). Ozone Disinfection for Elimination of Bacteria and Degradation of SARS-CoV2 RNA for Medical Environments. Genes, 14(1), 85. https://doi.org/10.3390/genes14010085