
Genetic Diversity of Cytochrome P450s CYP6M2 and CYP6P4 Associated with Pyrethroid Resistance in the Major Malaria Vectors Anopheles coluzzii and Anopheles gambiae from Yaoundé, Cameroon

 , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Clearance
2.2. Mosquito Samples Processing
2.2.1. Mosquito Collection
2.2.2. Insecticide Bioassays and PBO-Based Synergist Test
2.2.3. Mosquito Selection
2.3. Molecular Analysis
2.3.1. Genotyping of Kdr-Mutation Alleles
2.3.2. Investigation of the Expression Profile of Six Main Candidate-Resistance Genes
2.3.3. Amplification and Direct Sequencing of Upstream and Full Gene Regions of CYP6M2 and CYP6P4
2.3.4. In Silico Prediction of Promoter Region of CYP6M2 and CYP6P4
2.3.5. Polymorphism-Based Analysis of Genomic DNA Sequence
2.3.6. Detection of Key Polymorphisms in An. gambiae CYP6M2 and CYP6P4
2.4. Data Analysis
3. Results
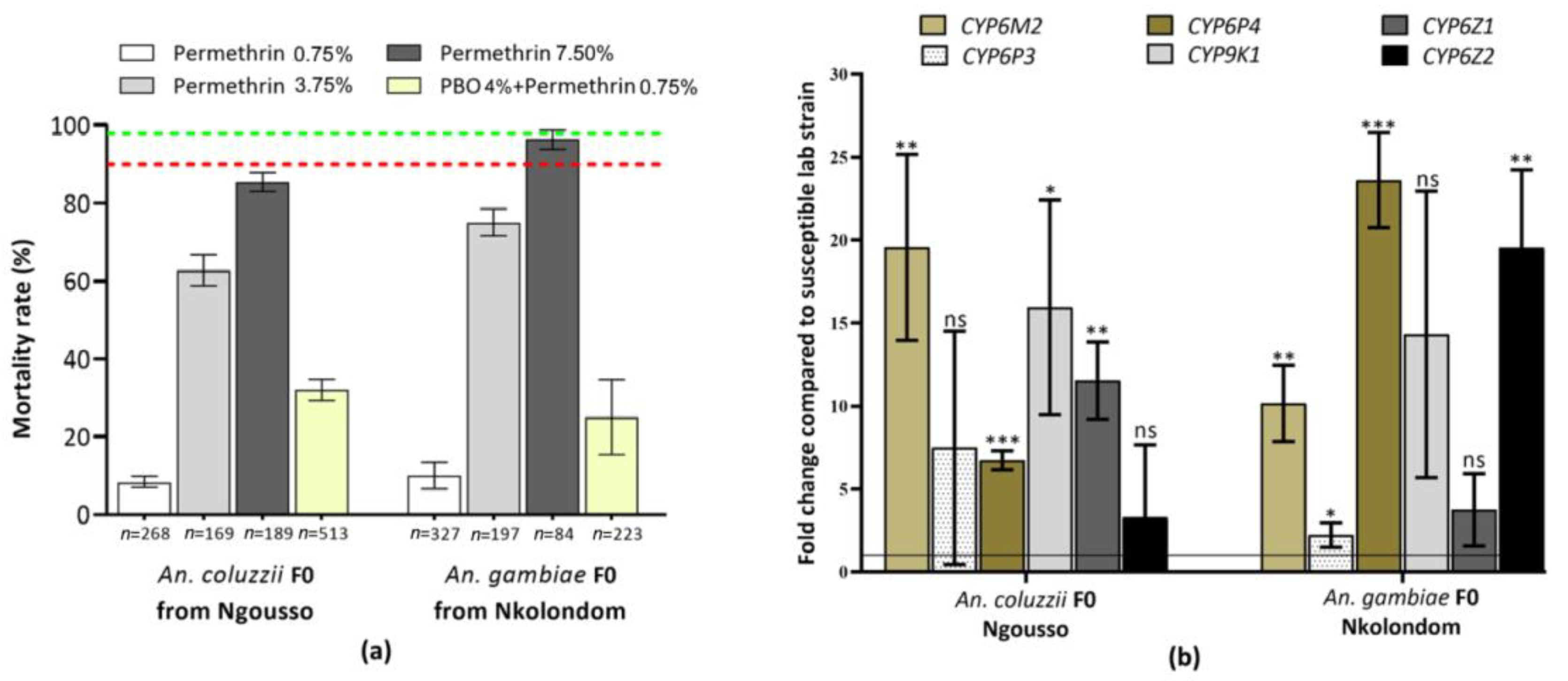
3.1. Species Identification and Susceptibility to Insecticides
3.2. Expression Profile of CYP6M2 and CYP6P4
3.3. Detection of the Knockdown Resistance Mutations
3.4. Association of Polymorphisms in CYP6M2 and Pyrethroid Resistance
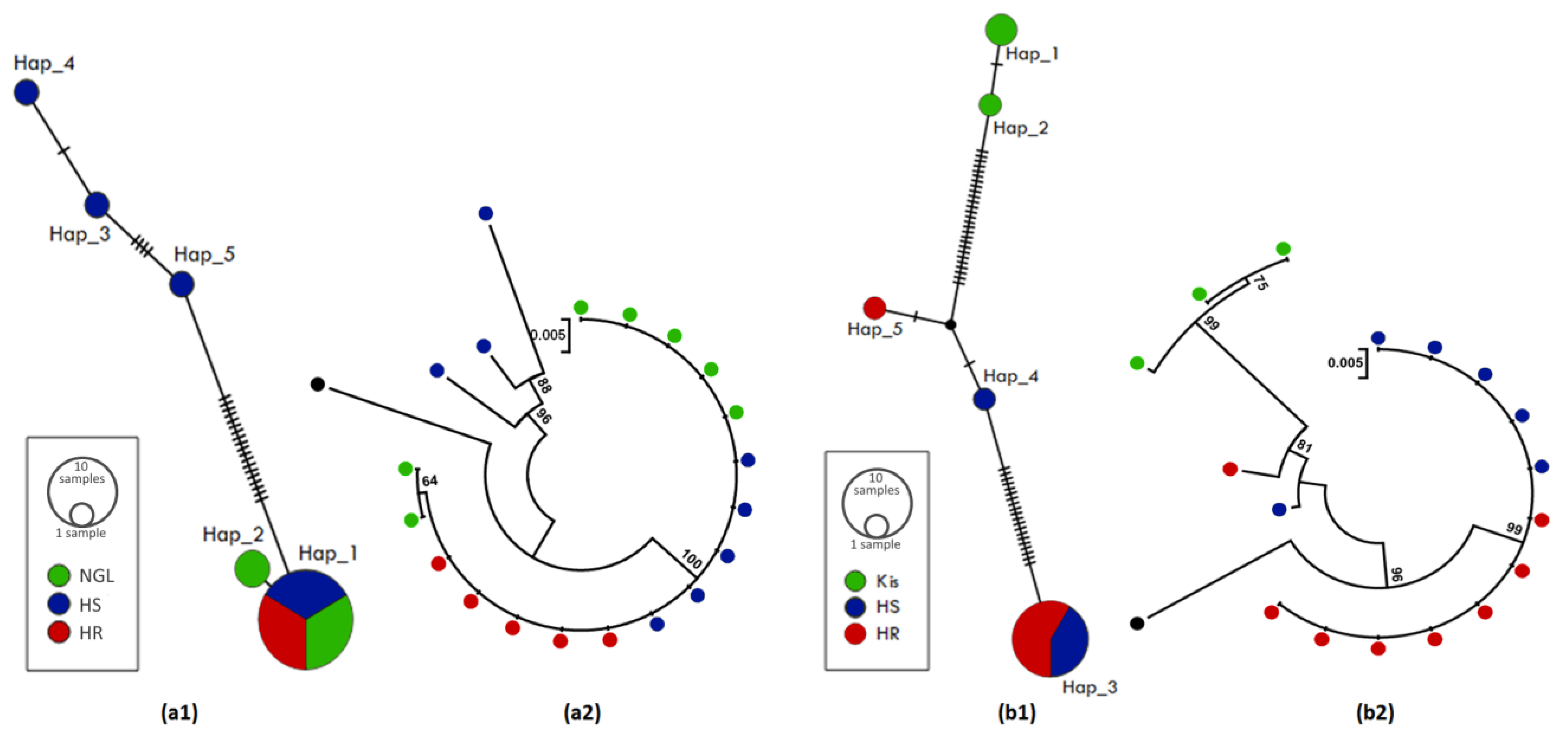
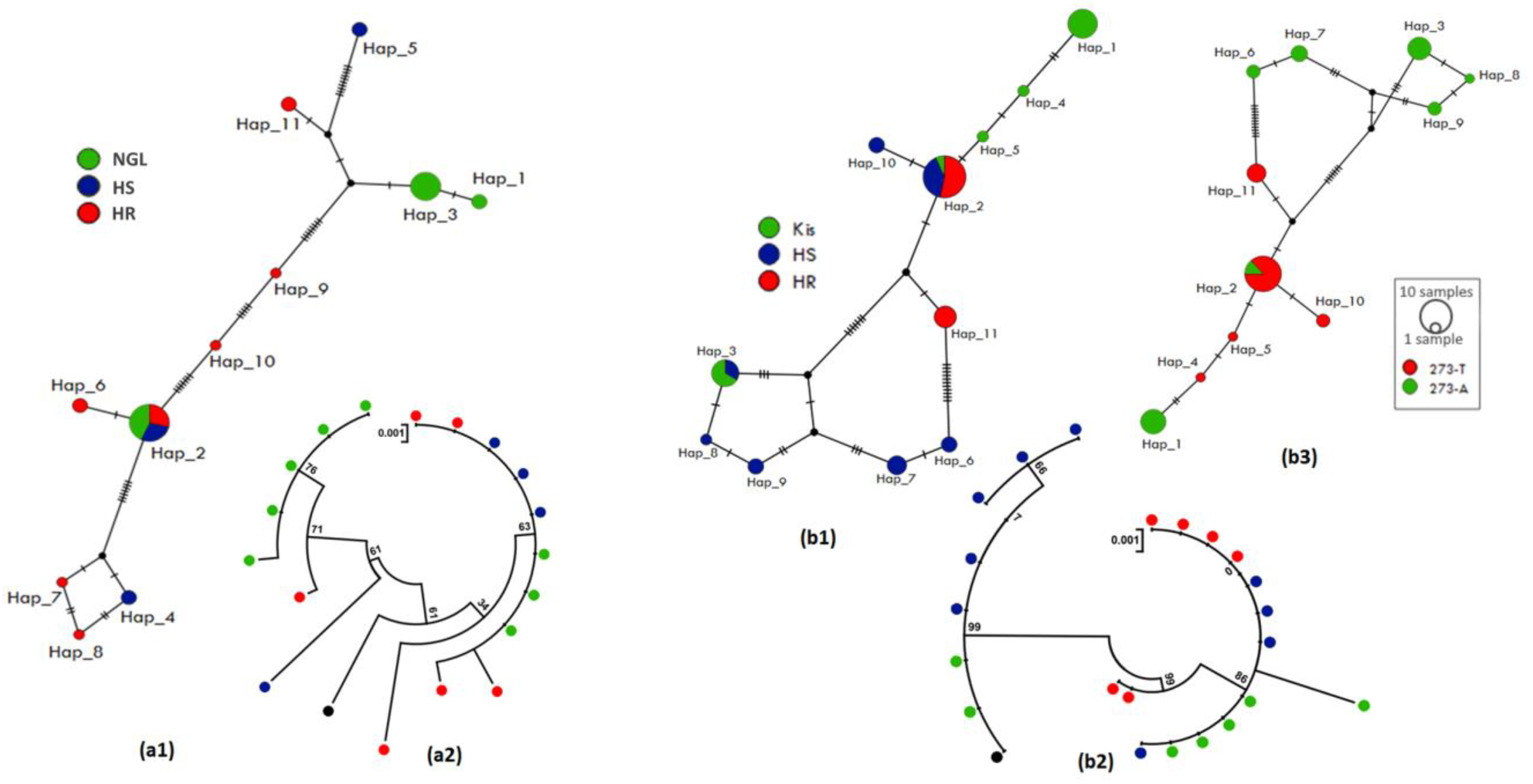
3.4.1. Polymorphism Analyses of the Upstream Region of CYP6M2
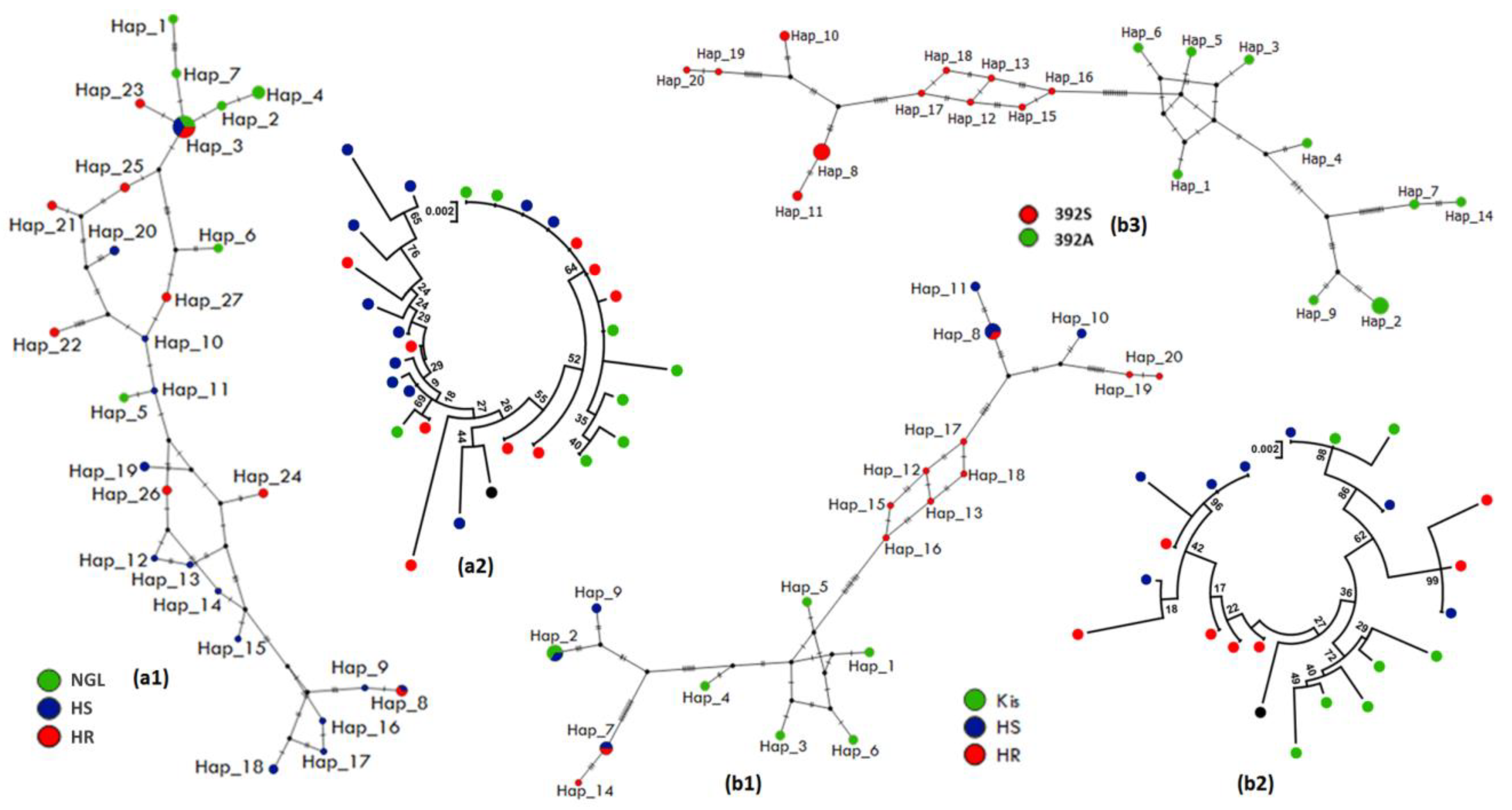
3.4.2. Polymorphism Analyses of the Full-Gene Region of CYP6M2
3.5. Genetic Polymorphisms Analysis of CYP6P4 Associated to Pyrethroid Resistance
3.5.1. Polymorphism Analyses of the Upstream Region of CYP6P4
3.5.2. Polymorphism Analyses of the Full-Gene Region of CYP6P4
3.6. Distribution of Key Mutations Found in CYP6M2 and CYP6P4 Genes among the Populations of An. gambiae
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bhatt, S.; Weiss, D.J.; Cameron, E.; Bisanzio, D.; Mappin, B.; Dalrymple, U.; Battle, K.E.; Moyes, C.L.; Henry, A.; Eckhoff, P.A.; et al. The effect of malaria control on Plasmodium falciparum in Africa between 2000 and 2015. Nature 2015, 526, 207–211. [Google Scholar] [CrossRef]
- WHO. World Malaria Report 2021; World Health Organization: Geneva, Switzerland, 2021. [Google Scholar]
- Antonio-Nkondjio, C.; Sonhafouo-Chiana, N.; Ngadjeu, C.S.; Doumbe-Belisse, P.; Talipouo, A.; Djamouko-Djonkam, L.; Kopya, E.; Bamou, R.; Awono-Ambene, P.; Wondji, C.S. Review of the evolution of insecticide resistance in main malaria vectors in Cameroon from 1990 to 2017. Parasites Vectors 2017, 10, 472. [Google Scholar] [CrossRef]
- Djouaka, R.F.; Bakare, A.A.; Coulibaly, O.N.; Akogbeto, M.C.; Ranson, H.; Hemingway, J.; Strode, C. Expression of the cytochrome P450s, CYP6P3 and CYP6M2 are significantly elevated in multiple pyrethroid resistant populations of Anopheles gambiae s.s. from Southern Benin and Nigeria. BMC Genom. 2008, 9, 538. [Google Scholar] [CrossRef]
- Hemingway, J. The role of vector control in stopping the transmission of malaria: Threats and opportunities. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20130431. [Google Scholar] [CrossRef]
- Ranson, H.; Lissenden, N. Insecticide Resistance in African Anopheles Mosquitoes: A Worsening Situation that Needs Urgent Action to Maintain Malaria Control. Trends Parasitol. 2016, 32, 187–196. [Google Scholar] [CrossRef]
- WHO. World Malaria Report 2019; World Health Organization: Geneva, Switzerland, 2019. [Google Scholar]
- Zhu, F.; Lavine, L.; O’Neal, S.; Lavine, M.; Foss, C.; Walsh, D. Insecticide resistance and management strategies in urban ecosystems. Insects 2016, 7, 2. [Google Scholar] [CrossRef]
- Hemingway, J.; Hawkes, N.J.; McCarroll, L.; Ranson, H. The molecular basis of insecticide resistance in mosquitoes. Insect Biochem. Mol. Biol. 2004, 34, 653–665. [Google Scholar] [CrossRef]
- WHO. Global Plan for Insecticide Resistance Management in Malaria Vectors: Executive Summary; World Health Organization: Geneva, Switzerland, 2012. [Google Scholar]
- Protopopoff, N.; Mosha, J.F.; Lukole, E.; Charlwood, J.D.; Wright, A.; Mwalimu, C.D.; Manjurano, A.; Mosha, F.W.; Kisinza, W.; Kleinschmidt, I. Effectiveness of a long-lasting piperonyl butoxide-treated insecticidal net and indoor residual spray interventions, separately and together, against malaria transmitted by pyrethroid-resistant mosquitoes: A cluster, randomised controlled, two-by-two factorial design trial. Lancet 2018, 391, 1577–1588. [Google Scholar]
- Staedke, S.G.; Gonahasa, S.; Dorsey, G.; Kamya, M.R.; Maiteki-Sebuguzi, C.; Lynd, A.; Katureebe, A.; Kyohere, M.; Mutungi, P.; Kigozi, S.P. Effect of long-lasting insecticidal nets with and without piperonyl butoxide on malaria indicators in Uganda (LLINEUP): A pragmatic, cluster-randomised trial embedded in a national LLIN distribution campaign. Lancet 2020, 395, 1292–1303. [Google Scholar] [CrossRef]
- Adolfi, A.; Poulton, B.; Anthousi, A.; Macilwee, S.; Ranson, H.; Lycett, G.J. Functional genetic validation of key genes conferring insecticide resistance in the major African malaria vector, Anopheles gambiae. Proc. Natl. Acad. Sci. USA 2019, 116, 25764–25772. [Google Scholar] [CrossRef]
- Ibrahim, S.S.; Riveron, J.M.; Stott, R.; Irving, H.; Wondji, C.S. The cytochrome P450 CYP6P4 is responsible for the high pyrethroid resistance in knockdown resistance-free Anopheles arabiensis. Insect Biochem. Mol. Biol. 2016, 68, 23–32. [Google Scholar] [CrossRef]
- Stevenson, B.J.; Bibby, J.; Pignatelli, P.; Muangnoicharoen, S.; O’Neill, P.M.; Lian, L.Y.; Muller, P.; Nikou, D.; Steven, A.; Hemingway, J.; et al. Cytochrome P450 6M2 from the malaria vector Anopheles gambiae metabolizes pyrethroids: Sequential metabolism of deltamethrin revealed. Insect Biochem. Mol. Biol. 2011, 41, 492–502. [Google Scholar] [CrossRef]
- Mitchell, S.N.; Stevenson, B.J.; Muller, P.; Wilding, C.S.; Egyir-Yawson, A.; Field, S.G.; Hemingway, J.; Paine, M.J.; Ranson, H.; Donnelly, M.J. Identification and validation of a gene causing cross-resistance between insecticide classes in Anopheles gambiae from Ghana. Proc. Natl. Acad. Sci. USA 2012, 109, 6147–6152. [Google Scholar] [CrossRef]
- Edi, C.V.; Djogbenou, L.; Jenkins, A.M.; Regna, K.; Muskavitch, M.A.; Poupardin, R.; Jones, C.M.; Essandoh, J.; Ketoh, G.K.; Paine, M.J.; et al. CYP6 P450 enzymes and ACE-1 duplication produce extreme and multiple insecticide resistance in the malaria mosquito Anopheles gambiae. PLoS Genet. 2014, 10, e1004236. [Google Scholar] [CrossRef]
- Wagah, M.G.; Korlević, P.; Clarkson, C.; Miles, A.; Lawniczak, M.K.; Makunin, A. Genetic variation at the Cyp6m2 putative insecticide resistance locus in Anopheles gambiae and Anopheles coluzzii. Malar. J. 2021, 20, 234. [Google Scholar] [CrossRef]
- Witzig, C.; Parry, M.; Morgan, J.C.; Irving, H.; Steven, A.; Cuamba, N.; Kerah-Hinzoumbé, C.; Ranson, H.; Wondji, C.S. Genetic mapping identifies a major locus spanning P450 clusters associated with pyrethroid resistance in kdr-free Anopheles arabiensis from Chad. Heredity 2013, 110, 389–397. [Google Scholar] [CrossRef][Green Version]
- Njoroge, H.; Van’t Hof, A.; Oruni, A.; Pipini, D.; Nagi, S.C.; Lynd, A.; Lucas, E.R.; Tomlinson, S.; Grau-Bove, X.; McDermott, D.; et al. Identification of a rapidly-spreading triple mutant for high-level metabolic insecticide resistance in Anopheles gambiae provides a real-time molecular diagnostic for antimalarial intervention deployment. Mol. Ecol. 2022, 31, 4307–4318. [Google Scholar] [CrossRef]
- Matowo, J.; Weetman, D.; Pignatelli, P.; Wright, A.; Charlwood, J.D.; Kaaya, R.; Shirima, B.; Moshi, O.; Lukole, E.; Mosha, J. Expression of pyrethroid metabolizing P450 enzymes characterizes highly resistant Anopheles vector species targeted by successful deployment of PBO-treated bednets in Tanzania. PLoS ONE 2022, 17, e0249440. [Google Scholar] [CrossRef]
- Tene-Fossog, B.; Poupardin, R.; Costantini, C.; Awono-Ambene, P.; Wondji, C.S.; Ranson, H. Resistance to DDT in an urban setting: Common mechanisms implicated in both M and S forms of Anopheles gambiae in the City of Yaoundé, Cameroon. PLoS ONE 2013, 8, e61408. [Google Scholar] [CrossRef]
- Bamou, R.; Sonhafouo-Chiana, N.; Mavridis, K.; Tchuinkam, T.; Wondji, C.S.; Vontas, J.; Antonio-Nkondjio, C. Status of insecticide resistance and its mechanisms in Anopheles gambiae and Anopheles coluzzii populations from forest settings in South-Cameroon. Genes 2019, 10, 741. [Google Scholar] [CrossRef]
- Piameu, M.; Nwane, P.; Toussile, W.; Mavridis, K.; Wipf, N.C.; Kouadio, P.F.; Mbakop, L.R.; Mandeng, S.; Ekoko, W.E.; Toto, J.C.; et al. Pyrethroid and Etofenprox Resistance in Anopheles gambiae and Anopheles coluzzii from Vegetable Farms in Yaoundé, Cameroon: Dynamics, Intensity and Molecular Basis. Molecules 2021, 26, 5543. [Google Scholar] [CrossRef]
- Menze, B.D.; Wondji, M.J.; Tchapga, W.; Tchoupo, M.; Riveron, J.M.; Wondji, C.S. Bionomics and insecticides resistance profiling of malaria vectors at a selected site for experimental hut trials in central Cameroon. Malar. J. 2018, 17, 317. [Google Scholar] [CrossRef]
- Liu, N. Insecticide resistance in mosquitoes: Impact, mechanisms, and research directions. Annu. Rev. Entomol. 2015, 60, 537–559. [Google Scholar] [CrossRef]
- Ibrahim, S.S.; Riveron, J.M.; Bibby, J.; Irving, H.; Yunta, C.; Paine, M.J.; Wondji, C.S. Allelic variation of cytochrome P450s drives resistance to bednet insecticides in a major malaria vector. PLoS Genet. 2015, 11, e1005618. [Google Scholar] [CrossRef]
- Mugenzi, L.M.; Menze, B.D.; Tchouakui, M.; Wondji, M.J.; Irving, H.; Tchoupo, M.; Hearn, J.; Weedall, G.D.; Riveron, J.M.; Wondji, C.S. Cis-regulatory CYP6P9b P450 variants associated with loss of insecticide-treated bed net efficacy against Anopheles funestus. Nat. Commun. 2019, 10, 4652. [Google Scholar] [CrossRef]
- Weedall, G.D.; Mugenzi, L.M.; Menze, B.D.; Tchouakui, M.; Ibrahim, S.S.; Amvongo-Adjia, N.; Irving, H.; Wondji, M.J.; Tchoupo, M.; Djouaka, R. A cytochrome P450 allele confers pyrethroid resistance on a major African malaria vector, reducing insecticide-treated bednet efficacy. Sci. Transl. Med. 2019, 11, eaat7386. [Google Scholar] [CrossRef]
- Yunta, C.; Hemmings, K.; Stevenson, B.; Koekemoer, L.L.; Matambo, T.; Pignatelli, P.; Voice, M.; Nász, S.; Paine, M.J.I. Cross-resistance profiles of malaria mosquito P450s associated with pyrethroid resistance against WHO insecticides. Pestic. Biochem. Physiol. 2019, 161, 61–67. [Google Scholar] [CrossRef]
- Ag1000G Anopheles gambiae Genomes Consortium: Genetic diversity of the African malaria vector Anopheles gambiae. Nature 2017, 552, 96. [CrossRef]
- Tene-Fossog, B.; Fotso-Toguem, Y.G.; Amvongo-Adjia, N.; Ranson, H.; Wondji, C.S.J.M.; Entomology, V. Temporal variation of high-level pyrethroid resistance in the major malaria vector Anopheles gambiae sl in Yaoundé, Cameroon, is mediated by target-site and metabolic resistance. Med. Vet. Entomol. 2022, 36, 247–259. [Google Scholar] [CrossRef]
- Kamdem, C.; Tene Fossog, B.; Simard, F.; Etouna, J.; Ndo, C.; Kengne, P.; Bousses, P.; Etoa, F.X.; Awono-Ambene, P.; Fontenille, D.; et al. Anthropogenic habitat disturbance and ecological divergence between incipient species of the malaria mosquito Anopheles gambiae. PLoS ONE 2012, 7, e39453. [Google Scholar] [CrossRef]
- Service, M. Sampling the larval population. Mosq. Ecol. Field Sampl. Methods 1993, 75–209. [Google Scholar] [CrossRef]
- WHO. Test Procedures for Insecticide Resistance Monitoring in Malaria Vectors; World Health Organization: Geneva, Switzerland, 2016; Volume 55. [Google Scholar]
- WHO. Standard Operating Procedure for Determining the Ability of PBO to Restore Susceptibility of Adult Mosquitoes to Pyrethroid Insecticides in WHO Tube Tests; 9240043853; World Health Organization: Geneva, Switzerland, 2022. [Google Scholar]
- Wondji, C.S.; Morgan, J.; Coetzee, M.; Hunt, R.H.; Steen, K.; Black, W.C.t.; Hemingway, J.; Ranson, H. Mapping a quantitative trait locus (QTL) conferring pyrethroid resistance in the African malaria vector Anopheles funestus. BMC Genom. 2007, 8, 34. [Google Scholar] [CrossRef]
- Cattel, J.; Faucon, F.; Le Péron, B.; Sherpa, S.; Monchal, M.; Grillet, L.; Gaude, T.; Laporte, F.; Dusfour, I.; Reynaud, S. Combining genetic crosses and pool targeted DNA-seq for untangling genomic variations associated with resistance to multiple insecticides in the mosquito Aedes aegypti. Evol. Appl. 2020, 13, 303–317. [Google Scholar] [CrossRef]
- Livak, K.J. Organization and mapping of a sequence on the Drosophila melanogaster X and Y chromosomes that is transcribed during spermatogenesis. Genetics 1984, 107, 611–634. [Google Scholar] [CrossRef]
- Gillies, M.; Coetzee, M. A supplement to the Anophelinae of Africa south of the Sahara (Afrotropical region). Publ. S Afr. Inst. Med. Res. 1987, 55, 1–143. [Google Scholar]
- Gillies, M.T.; De Meillon, B. The anophelinae of Africa south of the Sahara (Ethiopian zoogeographical region). Anophelinae Afr. South Sahara (Ethiop. Zoogeogr. Reg.) 1968, 54, 1–343. [Google Scholar]
- Santolamazza, F.; Mancini, E.; Simard, F.; Qi, Y.; Tu, Z.; della Torre, A. Insertion polymorphisms of SINE200 retrotransposons within speciation islands of Anopheles gambiae molecular forms. Malar. J. 2008, 7, 163. [Google Scholar] [CrossRef]
- Bass, C.; Nikou, D.; Donnelly, M.J.; Williamson, M.S.; Ranson, H.; Ball, A.; Vontas, J.; Field, L.M. Detection of knockdown resistance (kdr) mutations in Anopheles gambiae: A comparison of two new high-throughput assays with existing methods. Malar. J. 2007, 6, 111. [Google Scholar] [CrossRef]
- Jones, C.M.; Liyanapathirana, M.; Agossa, F.R.; Weetman, D.; Ranson, H.; Donnelly, M.J.; Wilding, C.S. Footprints of positive selection associated with a mutation (N1575Y) in the voltage-gated sodium channel of Anopheles gambiae. Proc. Natl. Acad. Sci. USA 2012, 109, 6614–6619. [Google Scholar] [CrossRef]
- Riveron, J.M.; Irving, H.; Ndula, M.; Barnes, K.G.; Ibrahim, S.S.; Paine, M.J.I.; Wondji, C.S. Directionally selected cytochrome P450 alleles are driving the spread of pyrethroid resistance in the major malaria vector Anopheles funestus. Proc. Natl. Acad. Sci. USA 2013, 110, 252–257. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C (T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Mavridis, K.; Wipf, N.; Medves, S.; Erquiaga, I.; Muller, P.; Vontas, J. Rapid multiplex gene expression assays for monitoring metabolic resistance in the major malaria vector Anopheles gambiae. Parasites Vectors 2019, 12, 9. [Google Scholar] [CrossRef]
- Lee, T.Y.; Chang, W.C.; Hsu, J.B.K.; Chang, T.H.; Shien, D.M. GPMiner: An integrated system for mining combinatorial cis-regulatory elements in mammalian gene group. BMC Genom. 2012, 13 (Suppl. 1), S3. [Google Scholar] [CrossRef]
- Farré, D.; Roset, R.; Huerta, M.; Adsuara, J.E.; Roselló, L.; Albà, M.M.; Messeguer, X. Identification of patterns in biological sequences at the ALGGEN server: PROMO and MALGEN. Nucleic Acids Res. 2003, 31, 3651–3653. [Google Scholar] [CrossRef]
- Messeguer, X.; Escudero, R.; Farré, D.; Núñez, O.; Martínez, J.; Albà, M.M. PROMO: Detection of known transcription regulatory elements using species-tailored searches. Bioinformatics 2002, 18, 333–334. [Google Scholar] [CrossRef]
- Kel-Margoulis, O.V.; Kel, A.E.; Reuter, I.; Deineko, I.V.; Wingender, E. TRANSCompel®: A database on composite regulatory elements in eukaryotic genes. Nucleic Acids Res. 2002, 30, 332–334. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Tchouakui, M.; Chiang, M.-C.; Ndo, C.; Kuicheu, C.K.; Amvongo-Adjia, N.; Wondji, M.J.; Tchoupo, M.; Kusimo, M.O.; Riveron, J.M.; Wondji, C.S. A marker of glutathione S-transferase-mediated resistance to insecticides is associated with higher Plasmodium infection in the African malaria vector Anopheles funestus. Sci. Rep. 2019, 9, 5772. [Google Scholar] [CrossRef]
- Kwiatkowska, R.; Platt, N.; Poupardin, R.; Irving, H.; Dabire, R.; Mitchell, S.; Jones, C.; Diabate, A.; Ranson, H.; Wondji, C. Dissecting the mechanisms responsible for the multiple insecticide resistance phenotype in Anopheles gambiae s.s., M form, from Vallee du Kou, Burkina Faso. Genes 2013, 519, 98–106. [Google Scholar] [CrossRef]
- Antonio-Nkondjio, C.; Fossog, B.T.; Ndo, C.; Djantio, B.M.; Togouet, S.Z.; Awono-Ambene, P.; Costantini, C.; Wondji, C.S.; Ranson, H. Anopheles gambiae distribution and insecticide resistance in the cities of Douala and Yaounde (Cameroon): Influence of urban agriculture and pollution. Malar. J. 2011, 10, 154. [Google Scholar] [CrossRef]
- Ndo, C.; Kopya, E.; Donbou, M.A.; Njiokou, F.; Awono-Ambene, P.; Wondji, C. Elevated Plasmodium infection rates and high pyrethroid resistance in major malaria vectors in a forested area of Cameroon highlight challenges of malaria control. Parasites Vectors 2018, 11, 157. [Google Scholar] [CrossRef]
- Subra, R.; Mosha, F. The effect of domestic detergents on the population dynamics of the immature stages of two competitor mosquitoes, Culex cinereus Theobald and Culex quinquefasciatus Say (Diptera, Culicidae) in Kenya. Acta Trop. 1984, 41, 69–75. [Google Scholar]
- Antonio-Nkondjio, C.; Youmsi-Goupeyou, M.; Kopya, E.; Tene-Fossog, B.; Njiokou, F.; Costantini, C.; Awono-Ambene, P. Exposure to disinfectants (soap or hydrogen peroxide) increases tolerance to permethrin in Anopheles gambiae populations from the city of Yaoundé, Cameroon. Malar. J. 2014, 13, 296. [Google Scholar] [CrossRef]
- Maitra, S.; Dombrowski, S.M.; Basu, M.; Raustol, O.; Waters, L.C.; Ganguly, R. Factors on the third chromosome affect the level of Cyp6a2 and Cyp6a8 expression in Drosophila melanogaster. Gene 2000, 248, 147–156. [Google Scholar] [CrossRef]
- Li, X.; Schuler, M.A.; Berenbaum, M.R. Molecular mechanisms of metabolic resistance to synthetic and natural xenobiotics. Annu. Rev. Entomol. 2007, 52, 231–253. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species (Size) | Populations | 2n | S | H (Hd) | Π | D | D * |
|---|---|---|---|---|---|---|---|
| An. coluzzii (932 bp) | HR F4 | 10 | 0 | 1 (0.000) | 0.00000 | n.a | n.a |
| HS F4 | 16 | 23 | 4 (0.600) | 0.01221 | 2.387 * | 1.592 ** | |
| Ngousso | 14 | 1 | 2 (0.440) | 0.00049 | 0.842 | 1.581 | |
| All | 40 | 24 | 5 (0.431) | 0.00645 | 0.103 | 1.722 ** | |
| An. gambiae (932 bp) | HR F4 | 16 | 23 | 2 (0.233) | 0.00583 | −0.917 | 1.623 ** |
| HS F4 | 12 | 20 | 2 (0.303) | 0.00659 | −0.373 | 1.593 ** | |
| Kisumu | 6 | 1 | 2 (0.533) | 0.00057 | 0.851 | 1.053 | |
| All | 34 | 49 | 5 (0.492) | 0.01831 | 1.474 | 1.859 ** |
| Species (Size) | Populations | 2n | S | H (Hd) | Syn | NSyn | π | D | D * | pKa/pKs |
|---|---|---|---|---|---|---|---|---|---|---|
| An. coluzzii (1104 bp) | HR F4 | 18 | 20 | 8 (0.915) | 11 | 7 | 0.0064 | 0.799 | 1.497 ** | 0.46 |
| HS F4 | 20 | 27 | 14 (0.953) | 13 | 10 | 0.0099 | 1.715 | 1.592 ** | 0.38 | |
| Ngousso | 18 | 17 | 7 (0.889) | 9 | 7 | 0.0045 | 0.059 | 1.4973 ** | 0.37 | |
| All | 56 | 32 | 27 (0.944) | 15 | 13 | 0.0097 | 1.238 | 1.8745 ** | 0.34 | |
| An. gambiae (1528 bp) | HR F4 | 14 | 36 | 11 (0.967) | 17 | 14 | 0.0102 | 1.621 | 1.493 ** | 0.26 |
| HS F4 | 14 | 44 | 6 (0.879) | 23 | 14 | 0.0122 | 1.515 | 1.493 ** | 0.19 | |
| Kisumu | 14 | 22 | 6 (0.879) | 14 | 5 | 0.0064 | 1.728 | 1.493 ** | 0.11 | |
| All | 42 | 51 | 20 (0.948) | 26 | 23 | 0.0124 | 2.124 * | 1.929 ** | 0.24 |
| Species (Size) | Population | 2n | S | H (Hd) | π | D | D * |
|---|---|---|---|---|---|---|---|
| An. coluzzii (870 bp) | HR F4 | 12 | 22 | 7 (0.879) | 0.00935 | 0.3557 | 1.509 |
| HS F4 | 8 | 20 | 3 (0.714) | 0.01071 | 1.1576 | 1.558 ** | |
| Ngousso | 16 | 15 | 3 (0.630) | 0.00822 | 2.3509 * | 1.509 ** | |
| All | 36 | 29 | 11 (0.803) | 0.01015 | 0.8198 | 1.749 * | |
| An. gambiae (880 bp) | HR F4 | 12 | 2 | 2 (0.489) | 0.00112 | 1.356 | 1.195 |
| HS F4 | 18 | 17 | 7 (0.856) | 0.00952 | 2.529 ** | 1.348 ** | |
| Kisumu | 14 | 15 | 5 (0.703) | 0.00754 | 1.537 | 1.288 ** | |
| All | 44 | 19 | 11 (0.838) | 0.00828 | 2.047 * | 1.655 ** |
| Species (Size) | Population | 2n | S | H (Hd) | Syn | NSyn | π | D | D * | pKa/pKs |
|---|---|---|---|---|---|---|---|---|---|---|
| An. coluzzii (1037 bp) | HR F4 | 18 | 21 | 8 (0.915) | 19 | 2 | 0.0076 | 1.114 | 1.246 ** | 0.002 |
| HS F4 | 20 | 20 | 9 (0.921) | 21 | 0 | 0.0078 | 1.365 | 1.268 | 0.000 | |
| Ngousso | 20 | 25 | 9 (0.922) | 19 | 7 | 0.0076 | 0.095 | 1.246 * | 0.078 | |
| All | 58 | 33 | 25 (0.963) | 25 | 9 | 0.0084 | 0.610 | 1.614 ** | 0.003 | |
| An. gambiae (1051 bp) | HR F4 | 20 | 11 | 12 (0.905) | 10 | 1 | 0.0041 | 1.324 | 1.007 | 0.030 |
| HS F4 | 18 | 13 | 13 (0.967) | 11 | 2 | 0.0048 | 1.275 | 0.667 * | 0.055 | |
| Kisumu | 20 | 16 | 13 (0.947) | 13 | 3 | 0.,0059 | 1.415 | 1.007 ** | 0.070 | |
| All | 58 | 17 | 33 (0.969) | 14 | 3 | 0.0062 | 2.385 * | 1.632 ** | 0.065 |
| Groups | Indel of 8 bp in Upstream of CYP6M2 | N | 2N | Allele | OR (CI) | p Value | |||
| D/D | D/I | I/I | D | I | |||||
| Kisumu | 0 | 3 | 14 | 17 | 34 | 0.09 | 0.91 | 11.40 (5.18–25.11) | <0.0001 |
| F0 | 6 | 8 | 5 | 19 | 38 | 0.53 | 0.47 | ||
| HR F4 | 18 | 21 | 15 | 54 | 108 | 0.53 | 0.47 | 0.73 (0.42–1.27) | 0.32 |
| HS F4 | 10 | 32 | 16 | 58 | 116 | 0.45 | 0.55 | ||
| Groups | Amino Acid Residue at 392 CYP6M2-Gene | N | 2N | Allele | OR (CI) | p Value | |||
| S/S | S/A | A/A | S | A | |||||
| Kisumu | 0 | 8 | 22 | 30 | 60 | 0.13 | 0.87 | 0.50 (0.19–1.32) | 0.23 |
| F0 | 1 | 6 | 49 | 56 | 112 | 0.07 | 0.93 | ||
| HR F4 | 13 | 23 | 14 | 50 | 100 | 0.49 | 0.51 | 0.59 (0.33–1.03) | 0.09 |
| HS F4 | 8 | 21 | 22 | 51 | 102 | 0.36 | 0.64 | ||
| Groups | Indel of 7 bp in Upstream of CYP6P4 | N | 2N | Allele | OR (CI) | p Value | |||
| t/t | t/a | a/a | t | a | |||||
| Kisumu | 0 | 6 | 6 | 12 | 24 | 0.25 | 0.75 | 1.46 (0.76–2.80) | 0.32 |
| F0 | 3 | 5 | 11 | 20 | 40 | 0.29 | 0.71 | ||
| HR F4 | 5 | 25 | 16 | 46 | 92 | 0.38 | 0.62 | 1.39 (0.79–2.44) | 0.32 |
| HS F4 | 8 | 21 | 11 | 40 | 80 | 0.46 | 0.54 | ||
| Groups | t/c Mutation at Codon 144 CYP6P4-Gene | N | 2N | Allele | OR (CI) | p Value | |||
| t/t | t/c | c/c | t | c | |||||
| Kisumu | 0 | 8 | 16 | 24 | 48 | 0.21 | 0.79 | 1.46 (0.76–2.80) | 0.32 |
| F0 | 3 | 5 | 12 | 20 | 40 | 0.28 | 0.72 | ||
| HR F4 | 25 | 19 | 0 | 44 | 88 | 0.78 | 0.22 | 0.52 (0.28–0.98) | 0.06 |
| HS F4 | 16 | 23 | 3 | 42 | 84 | 0.65 | 0.35 | ||
| Groups | Amino Acid Residue at 168 CYP6P4-Gene | N | 2N | Allele | OR (CI) | p Value | |||
| C/C | C/S | S/S | C | S | |||||
| Kisumu | 0 | 8 | 12 | 20 | 40 | 0.20 | 0.80 | 1.00 (0.50–1.99) | 1.00 |
| F0 | 0 | 10 | 15 | 25 | 50 | 0.20 | 0.80 | ||
| HR F4 | 3 | 22 | 5 | 30 | 60 | 0.47 | 0.53 | 1.00 (0.57–1.74) | 1.00 |
| HS F4 | 6 | 16 | 8 | 30 | 60 | 0.47 | 0.53 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fotso-Toguem, Y.; Tene-Fossog, B.; Mugenzi, L.M.J.; Wondji, M.J.; Njiokou, F.; Ranson, H.; Wondji, C.S. Genetic Diversity of Cytochrome P450s CYP6M2 and CYP6P4 Associated with Pyrethroid Resistance in the Major Malaria Vectors Anopheles coluzzii and Anopheles gambiae from Yaoundé, Cameroon. Genes 2023, 14, 52. https://doi.org/10.3390/genes14010052
Fotso-Toguem Y, Tene-Fossog B, Mugenzi LMJ, Wondji MJ, Njiokou F, Ranson H, Wondji CS. Genetic Diversity of Cytochrome P450s CYP6M2 and CYP6P4 Associated with Pyrethroid Resistance in the Major Malaria Vectors Anopheles coluzzii and Anopheles gambiae from Yaoundé, Cameroon. Genes. 2023; 14(1):52. https://doi.org/10.3390/genes14010052
Chicago/Turabian StyleFotso-Toguem, Yvan, Billy Tene-Fossog, Leon M. J. Mugenzi, Murielle J. Wondji, Flobert Njiokou, Hilary Ranson, and Charles S. Wondji. 2023. "Genetic Diversity of Cytochrome P450s CYP6M2 and CYP6P4 Associated with Pyrethroid Resistance in the Major Malaria Vectors Anopheles coluzzii and Anopheles gambiae from Yaoundé, Cameroon" Genes 14, no. 1: 52. https://doi.org/10.3390/genes14010052
APA StyleFotso-Toguem, Y., Tene-Fossog, B., Mugenzi, L. M. J., Wondji, M. J., Njiokou, F., Ranson, H., & Wondji, C. S. (2023). Genetic Diversity of Cytochrome P450s CYP6M2 and CYP6P4 Associated with Pyrethroid Resistance in the Major Malaria Vectors Anopheles coluzzii and Anopheles gambiae from Yaoundé, Cameroon. Genes, 14(1), 52. https://doi.org/10.3390/genes14010052