
Association of CX36 Protein Encoding Gene GJD2 with Refractive Errors

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Study Samples
2.3. Refractive Error Measurement
2.4. DNA Extraction
2.5. Genotyping
2.6. Verification of Zygosity
2.7. Twin Method
2.8. Statistical Analysis
3. Results
3.1. Sample Characteristics for Heritability Assessment of Hyperopia and Astigmatism
3.2. Heritability of Hyperopia and Astigmatism using the Twin Method
3.3. Sample Characteristics for SNPs Studies
3.4. Associations of SNPs with Refractive Errors
3.5. Haplotype Analysis
3.6. Associations of SNPs with Degrees of Myopia and Hyperopia
4. Discussion
5. Limitation(s)
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Krishnaiah, S.; Srinivas, M.; Khanna, R.C.; Rao, G.N. Prevalence and risk factors for refractive errors in the South Indian adult population: The Andhra Pradesh Eye disease study. Clin. Ophthalmol. 2009, 3, 17–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, K.M.; Verhoeven, V.J.M.; Cumberland, P.; Bertelsen, G.; Wolfram, C.; Buitendijk, G.H.S.; Hofman, A.; van Duijn, C.M.; Vingerling, J.R.; Kuijpers, R.W.A.M.; et al. Prevalence of refractive error in Europe: The European Eye Epidemiology (E3) Consortium. Eur. J. Epidemiol. 2015, 30, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Verhoeven, V.J.M.; Hysi, P.G.; Wojciechowski, R.; Fan, Q.; Guggenheim, J.A.; Höhn, R.; Macgregor, S.; Hewitt, A.W.; Buitendijk, G.H.S.; Mcmahon, G.; et al. NIH Public Access multiple new susceptibility loci for refractive error and myopia. Nature Genetics 2013, 45, 314–318. [Google Scholar] [CrossRef] [PubMed]
- Stambolian, D. Genetic susceptibility and mechanisms for refractive error. Clin. Genet. 2013, 84, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Morgan, I.G.; Wu, P.C.; Ostrin, L.A.; Tideman, J.W.; Yam, J.C.; Lan, W.; Baraas, R.C.; He, X.; Sankaridurg, P.; Saw, S.M.; et al. IMI risk factors for myopia. Investig. Ophthalmol. Vis. Sci. 2021, 62, 12–15. [Google Scholar] [CrossRef]
- Wojciechowski, R. Nature and nurture: The complex genetics of myopia and refractive error. Clin. Genet. 2010, 79, 301–320. [Google Scholar] [CrossRef]
- Foster, P.J.; Jiang, Y. Epidemiology of myopia. Eye 2014, 28, 202–208. [Google Scholar] [CrossRef] [Green Version]
- Castagno, V.D.; Fassa, A.G.; Carret, M.L.V.; Vilela, M.A.P.; Meucci, R.D. Hyperopia: A meta-analysis of prevalence and a review of associated factors among school-aged children. BMC Ophthalmol. 2014, 14, 163. [Google Scholar] [CrossRef] [Green Version]
- Quint, W.H.; Tadema, K.C.D.; de Vrieze, E.; Lukowicz, R.M.; Broekman, S.; Winkelman, B.H.J.; Hoevenaars, M.; de Gruiter, H.M.; van Wijk, E.; Schaeffel, F.; et al. Loss of Gap Junction Delta-2 (GJD2) gene orthologs leads to refractive error in zebrafish. Commun. Biol. 2021, 4, 676. [Google Scholar] [CrossRef]
- van der Sande, E.; Haarman, A.E.G.; Quint, W.H.; Tadema, K.C.D.; Meester-smoor, M.A.; Kamermans, M.; Zeeuw, C.I.; De Klaver, C.C.W.; Winkelman, B.H.J.; Iglesias, A.I. The Role of GJD2 ( Cx36 ) in Refractive Error Development. Investig. Opthalmology Vis. Sci. 2022, 63, 5. [Google Scholar] [CrossRef]
- Kántor, O.; Varga, A.; Nitschke, R.; Naumann, A.; Énzsöly, A.; Lukáts, A.; Szabó, A.; Németh, J.; Völgyi, B. Bipolar cell gap junctions serve major signaling pathways in the human retina. Anat. Embryol. 2017, 222, 2603–2624. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Wong, K.Y.; Zhang, D.Q. Mapping physiological inputs from multiple photoreceptor systems to dopaminergic amacrine cells in the mouse retina. Sci. Rep. 2017, 7, 7920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srinivas, M.; Verselis, V.K.; White, T.W. Human diseases associated with connexin mutations. Biochim. Biophys. Acta-Biomembr. 2018, 1860, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Fernández, V.; Milosavljevic, N.; Allen, A.E.; Vessey, K.A.; Jobling, A.I.; Fletcher, E.L.; Breen, P.P.; Morley, J.W.; Cameron, M.A. Rod Photoreceptor Activation Alone Defines the Release of Dopamine in the Retina. Curr. Biol. 2019, 29, 763–774.e5. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.H.; Qian, K.W.; Xu, G.Z.; Li, Y.Y.; Ma, Y.Y.; Huang, F.; Wang, Y.Q.; Zhou, X.; Qu, J.; Yang, X.L.; et al. The role of retinal dopamine in C57BL/6 mouse refractive development as revealed by intravitreal administration of 6-hydroxydopamine. Investig. Ophthalmol. Vis. Sci. 2016, 57, 5393–5404. [Google Scholar] [CrossRef]
- Chuang, K.K.; Rucker, F.J. The role of dopamine in eye growth responses to color and luminance flicker in chicks. Exp. Eye Res. 2019, 189, 107822. [Google Scholar] [CrossRef]
- Francisco, B.M.; Salvador, M.; Amparo, N. Oxidative stress in myopia. Oxid. Med. Cell. Longev. 2015, 2015, 750637. [Google Scholar] [CrossRef] [Green Version]
- Carr, B.J.; Stell, W.K. Nitric Oxide (NO) Mediates the Inhibition of Form-Deprivation Myopia by Atropine in Chicks. Sci. Rep. 2016, 6, 9. [Google Scholar] [CrossRef] [Green Version]
- Simpson, C.L.; Wojciechowski, R.; Oexle, K.; Murgia, F.; Portas, L.; Li, X.; Virginie, J.M.V.; Vitart, V.; Schache, M.; Mohsen Hosseini, S.; et al. Genome-wide meta-analysis of myopia and hyperopia provides evidence for replication of 11 loci. PLoS ONE 2014, 9, e107110. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Pardue, M.T.; Iuvone, P.M.; Qu, J. Dopamine signaling and myopia development: What are the key challenges. Prog. Retin. Eye Res. 2017, 61, 60–71. [Google Scholar] [CrossRef]
- Kunceviciene, E.; Sriubiene, M.; Liutkeviciene, R.; Miceikiene, I.T.; Smalinskiene, A. Heritability of myopia and its relation with GDJ2 and RASGRF1 genes in Lithuania. BMC Ophthalmol. 2018, 18, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, G.Y.; Liu, F.Y.; Li, X.; Zhu, Q.R.; Chen, B.J.; Liu, L.Q. Decreased expression of gap junction delta-2 (GJD2) messenger RNA and connexin 36 protein in form-deprivation myopia of guinea pigs. Chin. Med. J. 2019, 132, 1700–1705. [Google Scholar] [CrossRef] [PubMed]
- Akopian, A.; Kumar, S.; Ramakrishnan, H.; Roy, K.; Viswanathan, S.; Bloomfield, S.A. Targeting neuronal gap junctions in mouse retina offers neuroprotection in glaucoma. J. Clin. Investig. 2017, 127, 2647–2661. [Google Scholar] [CrossRef] [PubMed]
- Kiefer, A.K.; Tung, J.Y.; Do, C.B.; Hinds, D.A.; Mountain, J.L.; Francke, U.; Eriksson, N. Genome-Wide Analysis Points to Roles for Extracellular Matrix Remodeling, the Visual Cycle, and Neuronal Development in Myopia. PLoS Genet. 2013, 9, e1003299. [Google Scholar] [CrossRef] [Green Version]
- Oishi, M.; Yamashiro, K.; Miyake, M.; Akagi-Kurashige, Y.; Kumagai, K.; Nakata, I.; Nakanishi, H.; Yoshikawa, M.; Oishi, A.; Gotoh, N.; et al. Association between ZIC2, RASGRF1, and SHISA6 genes and high myopia in Japanese subjects. Investig. Ophthalmol. Vis. Sci. 2013, 54, 7492–7497. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.; Shan, G.; Ma, J.; Zhong, Y. Polymorphism in the RASGRF1 gene with high myopia: A meta-analysis. Mol. Vis. 2015, 21, 1272–1280. [Google Scholar]
- Shah, R.L.; Li, Q.; Zhao, W.; Tedja, M.S.; Tideman, J.W.L.; Khawaja, A.P.; Fan, Q.; Yazar, S.; Williams, K.M.; Verhoeven, V.J.M.; et al. A genome-wide association study of corneal astigmatism: The CREAM consortium. Mol. Vis. 2018, 24, 127–142. [Google Scholar]
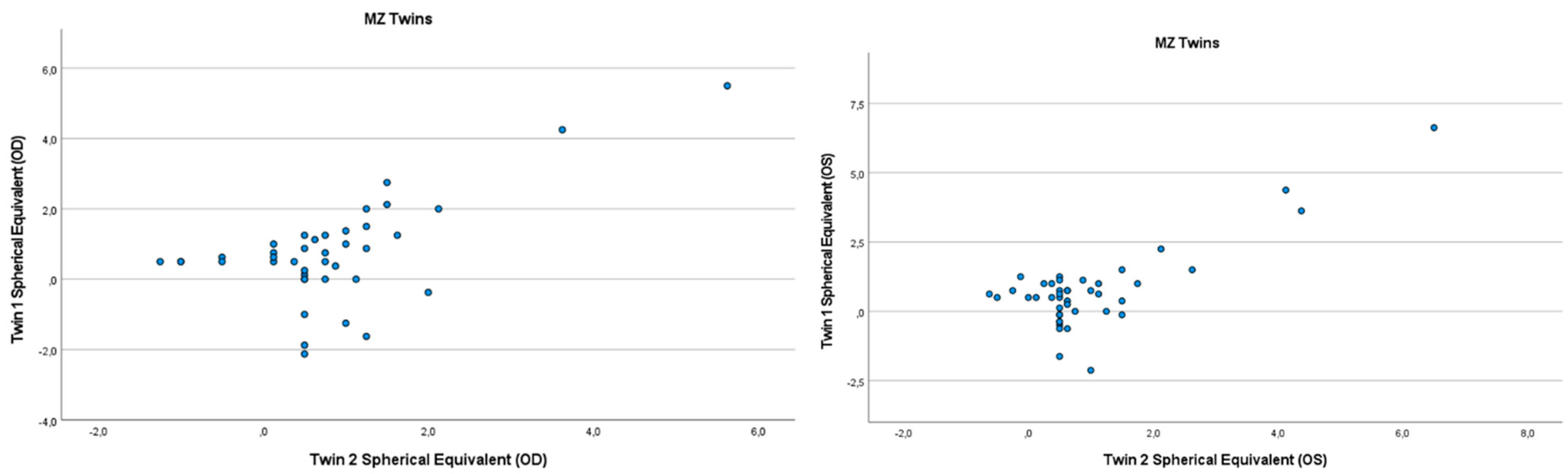
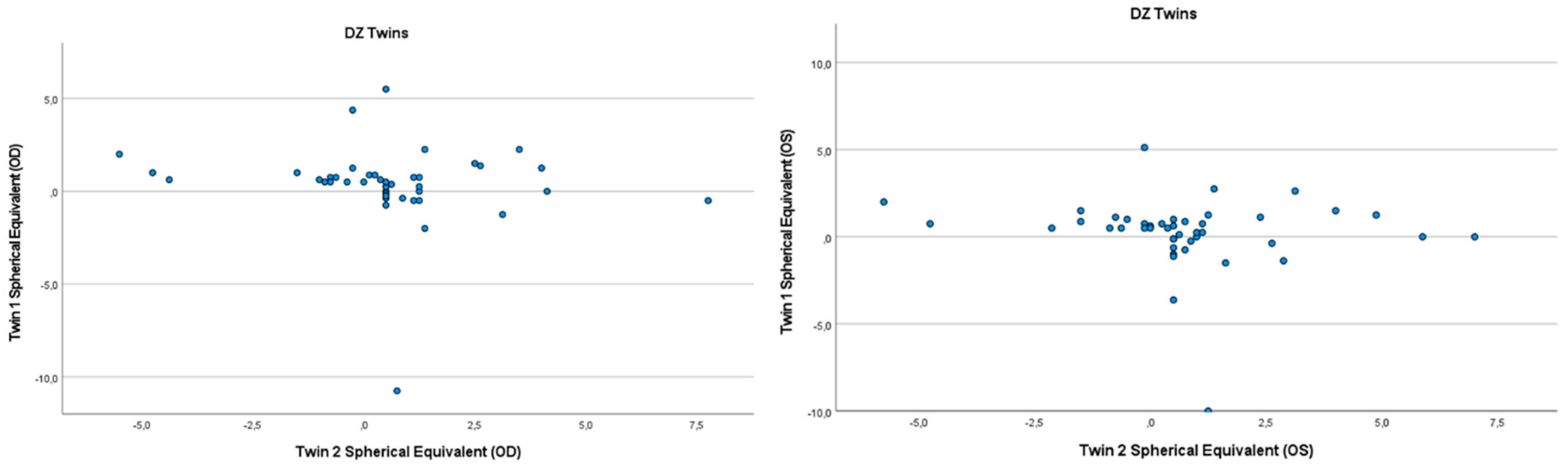

{kind=link}
{kind=link}
| Characteristics | MZ Twins | DZ Twins | p Value |
|---|---|---|---|
| Sex, pairs | |||
| Male | 8 | 5 | 0.923 * |
| Female | 14 | 10 | |
| Male/Female | 6 | ||
| Age, years | |||
| Mean ± SE | 22.18 ± 0.51 | 23.03 ± 0.63 | 0.694 ** |
| Median | 20.11 | 22.31 | |
| Min, Max | 18, 39 | 20, 40 | |
| Spherical equivalent, D | |||
| OD Mean ± SE | 3.155 ± 0.20 | 2.995 ± 0.19 | 0.425 ** |
| Median | 3.055 | 2.850 | |
| Min, Max | 0.750, 6.500 | 0.825, 5.125 | |
| OS Mean ± SE | 3.145 ± 0.85 | 2.850 ± 0.63 | 0.071 ** |
| Median | 2.122 | 1.965 | |
| Min, Max | 0.725, 5.65 | 0.815, 3.52 | |
| Myopia (n = 48) | Hyperopia (n = 14) | Myopia with Astigmatism (n = 239) | Hyperopia with Astigmatism (n = 72) | Control (n = 104) | |
|---|---|---|---|---|---|
| Gender | |||||
| Male, n (%) | 17 (35.42) | 6 (42.86) | 83 (34.73) | 32 (44.44) | 46 (44.23) |
| Female, n (%) | 31 (64.58) | 8 (57.14) | 156 (65.27) | 40 (55.56) | 58 (55.77) |
| p-value * | <0.001 | <0.001 | <0.001 | <0.001 | – |
| Age | |||||
| Median (Min; Max) | 20 (18; 40) | 20 (19; 26) | 20 (18; 40) | 22.5 (18; 40) | 20 (18; 40) |
| p-value * | 0.493 | 0.926 | 0.602 | 0.086 | – |
| Spherical equivalent, D | |||||
| OD | |||||
| Median | −2.75 | 2.35 | −11.25 | 2.5 | 0.15 |
| (Min; Max) | (−5.00; −1.5) | (1; 4.5) | (−17.35; −1.25) | (1.35; 4.37) | (−0.35; 0.35) |
| p-value * | <0.001 | <0.001 | <0.001 | <0.001 | – |
| OS | |||||
| Median | −2.5 | 2.25 | −7.125 | 2.75 | −0.125 |
| (Min; Max) | (-3.5; −1.75) | (1.25; 3.75) | (−16.75; −1.35) | (1.875; 5.125) | (0.125; −0.375) |
| p-value * | <0.001 | <0.001 | <0.001 | <0.001 | – |
| Gene | SNP | Genotype | Hyperopia with Astigmatism (n = 72) | Control (n = 104) | OR | 95% C.I. | p Value | |
|---|---|---|---|---|---|---|---|---|
| Lower | Upper | |||||||
| GJD2 | rs634990 | CC | 17 | 23 | 1.060 | 0.517 | 2.174 | 0.874 |
| CT | 30 | 59 | 0.508 | 0.273 | 0.946 | 0.033 * | ||
| TT | 23 | 17 | 2.360 | 1.146 | 4.860 | 0.020 * | ||
| rs524952 | AA | 17 | 19 | 1.298 | 0.616 | 2.735 | 0.492 | |
| AT | 31 | 57 | 0.529 | 0.281 | 0.997 | 0.049 * | ||
| TT | 20 | 17 | 1.863 | 0.888 | 3.907 | 0.100 | ||
| RASGRF1 | rs8027411 | TT | 16 | 20 | 1.186 | 0.566 | 2.484 | 0.652 |
| TG | 45 | 63 | 1.058 | 0.569 | 1.968 | 0.858 | ||
| GG | 11 | 20 | 0.748 | 0.334 | 1.676 | 0.481 | ||
| rs4778879 | AA | 23 | 41 | 0.671 | 0.352 | 1.276 | 0.224 | |
| AG | 38 | 39 | 1.792 | 0.959 | 3.348 | 0.068 | ||
| GG | 8 | 16 | 0.656 | 0.263 | 1.632 | 0.364 | ||
| rs28412916 | CC | 22 | 38 | 0.705 | 0.366 | 1.356 | 0.295 | |
| CA | 38 | 39 | 1.786 | 0.951 | 3.356 | 0.071 | ||
| AA | 8 | 17 | 0.604 | 0.244 | 1.494 | 0.275 | ||
| GJD2 (rs634990) | GJD2 (rs524952) | Frequency, n (%) | OR | 95% C.I. | p Value | ||
|---|---|---|---|---|---|---|---|
| Control | Hyperopia | Lower | Upper | ||||
| T | T | 68 (76.4) | 9 (69.2) | 0.838 | 0.416 | 1.688 | 0.621 |
| C | T | 56 (62.9) | 6 (46.2) | 0.410 | 0.220 | 0.765 | 0.005 * |
| T | A | 53 (59.6) | 7 (53.8) | 0.613 | 0.332 | 1.132 | 0.118 |
| C | A | 72 (80.9) | 10 (76.9) | 0.501 | 0.246 | 1.019 | 0.056 |
| Haplotype | Frequency, n (%) | OR | 95% C.I. | p Value | |||
|---|---|---|---|---|---|---|---|
| GJD2 (rs634990) | GJD2 (rs524952) | Control | Hyperopia with Astigmatism | Lower | Upper | ||
| T | T | 68 (76.4) | 49 (74.2) | 0.890 | 0.426 | 1.861 | 0.757 |
| C | T | 56 (62.9) | 26 (39.4) | 0.383 | 0.199 | 0.737 | 0.004 * |
| T | A | 53 (59.6) | 30 (45.5) | 0.566 | 0.297 | 1.077 | 0.083 |
| C | A | 72 (80.9) | 43 (65.2) | 0.441 | 0.212 | 0.918 | 0.029 * |
| Gene (SNP) | Genotype | Degree of Refraction, D | OD | OS | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| p Value | OR | 95% C.I. | p Value | OR | 95% C.I. | |||||
| Lower | Upper | Lower | Upper | |||||||
| Hyperopia and Hyperopia with Astigmatism (n = 287) | ||||||||||
| GJD2 (rs524952) | AA | Low (<+2.25) | 0.350 | 2.00 | 0.467 | 8.557 | 0.812 | 0.833 | 0.185 | 3.750 |
| Moderate (+2.25 to +5.00) | – | – | – | – | – | – | – | |||
| AT | Low (<+2.25) | 0.045 * | 0.255 | 0.067 | 0.971 | 0.768 | 0.848 | 0.284 | 2.533 | |
| Moderate (+2.25 to +5.00) | – | – | – | – | 0.850 | 1.273 | 0.105 | 15.385 | ||
| TT | Low (<+2.25) | 0.151 | 2.625 | 0.702 | 9.809 | 0.592 | 1.400 | 0.409 | 4.791 | |
| Moderate (+2.25 to +5.00) | – | – | – | – | 0.664 | 1.750 | 0.140 | 21.876 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kunceviciene, E.; Muskieta, T.; Sriubiene, M.; Liutkeviciene, R.; Smalinskiene, A.; Grabauskyte, I.; Insodaite, R.; Juoceviciute, D.; Kucinskas, L. Association of CX36 Protein Encoding Gene GJD2 with Refractive Errors. Genes 2022, 13, 1166. https://doi.org/10.3390/genes13071166
Kunceviciene E, Muskieta T, Sriubiene M, Liutkeviciene R, Smalinskiene A, Grabauskyte I, Insodaite R, Juoceviciute D, Kucinskas L. Association of CX36 Protein Encoding Gene GJD2 with Refractive Errors. Genes. 2022; 13(7):1166. https://doi.org/10.3390/genes13071166
Chicago/Turabian StyleKunceviciene, Edita, Tomas Muskieta, Margarita Sriubiene, Rasa Liutkeviciene, Alina Smalinskiene, Ingrida Grabauskyte, Ruta Insodaite, Dovile Juoceviciute, and Laimutis Kucinskas. 2022. "Association of CX36 Protein Encoding Gene GJD2 with Refractive Errors" Genes 13, no. 7: 1166. https://doi.org/10.3390/genes13071166
APA StyleKunceviciene, E., Muskieta, T., Sriubiene, M., Liutkeviciene, R., Smalinskiene, A., Grabauskyte, I., Insodaite, R., Juoceviciute, D., & Kucinskas, L. (2022). Association of CX36 Protein Encoding Gene GJD2 with Refractive Errors. Genes, 13(7), 1166. https://doi.org/10.3390/genes13071166