
Gene Network of Susceptibility to Atypical Femoral Fractures Related to Bisphosphonate Treatment

 , , , ,
, , , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Whole-Exome Sequencing (WES)
2.3. AFF Network Construction (AFFNET)
3. Results
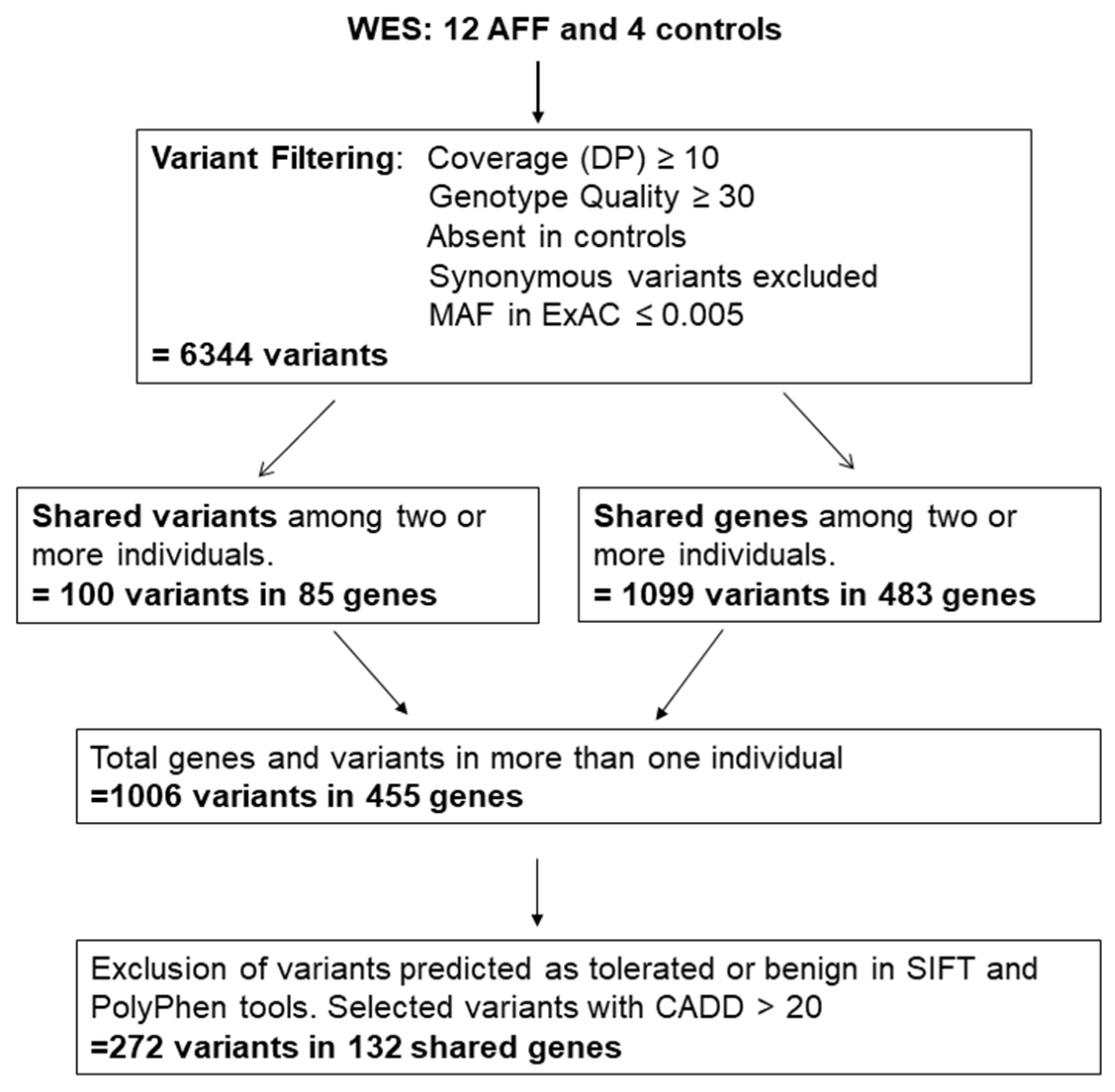
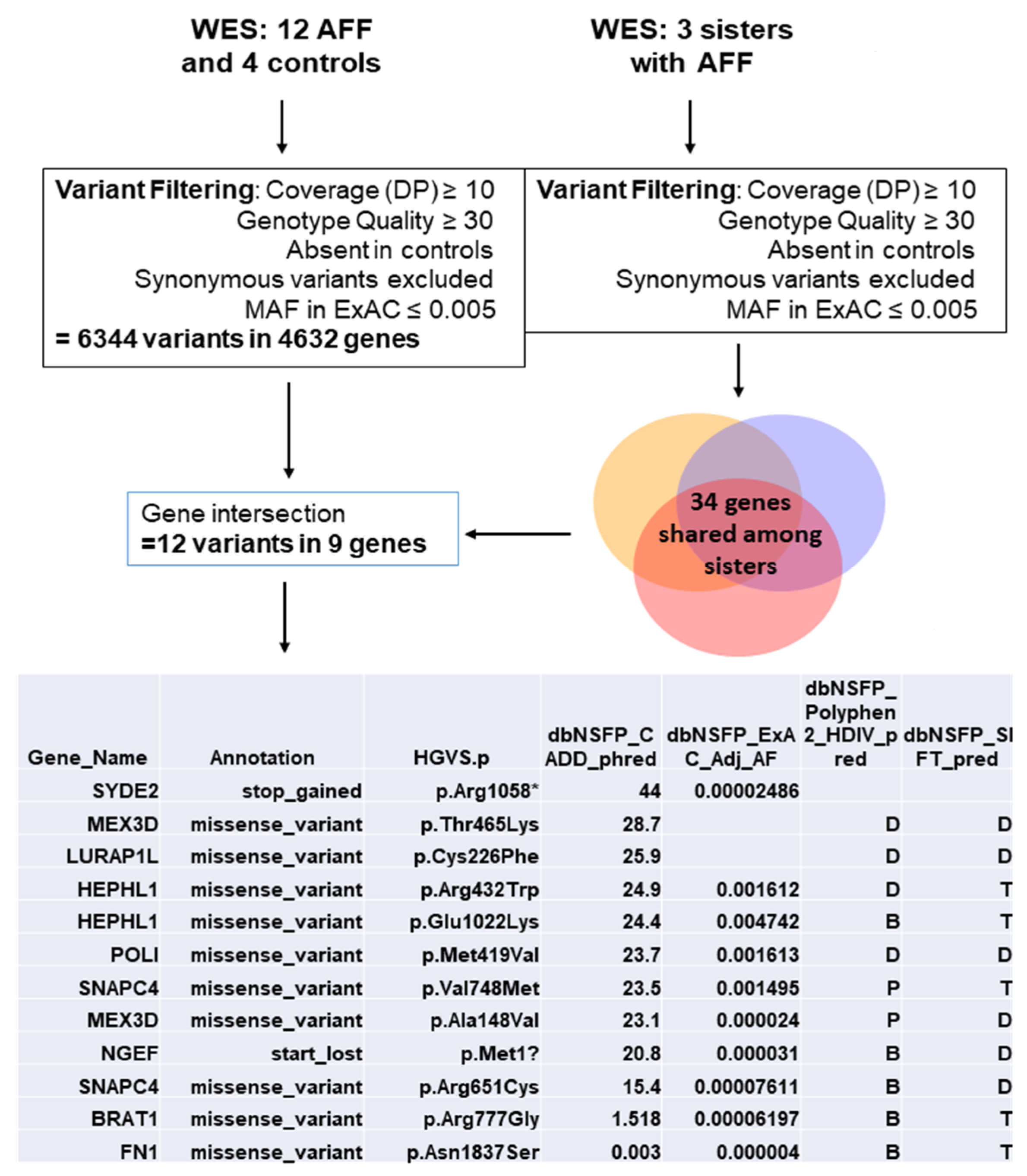
3.1. Variant Selection
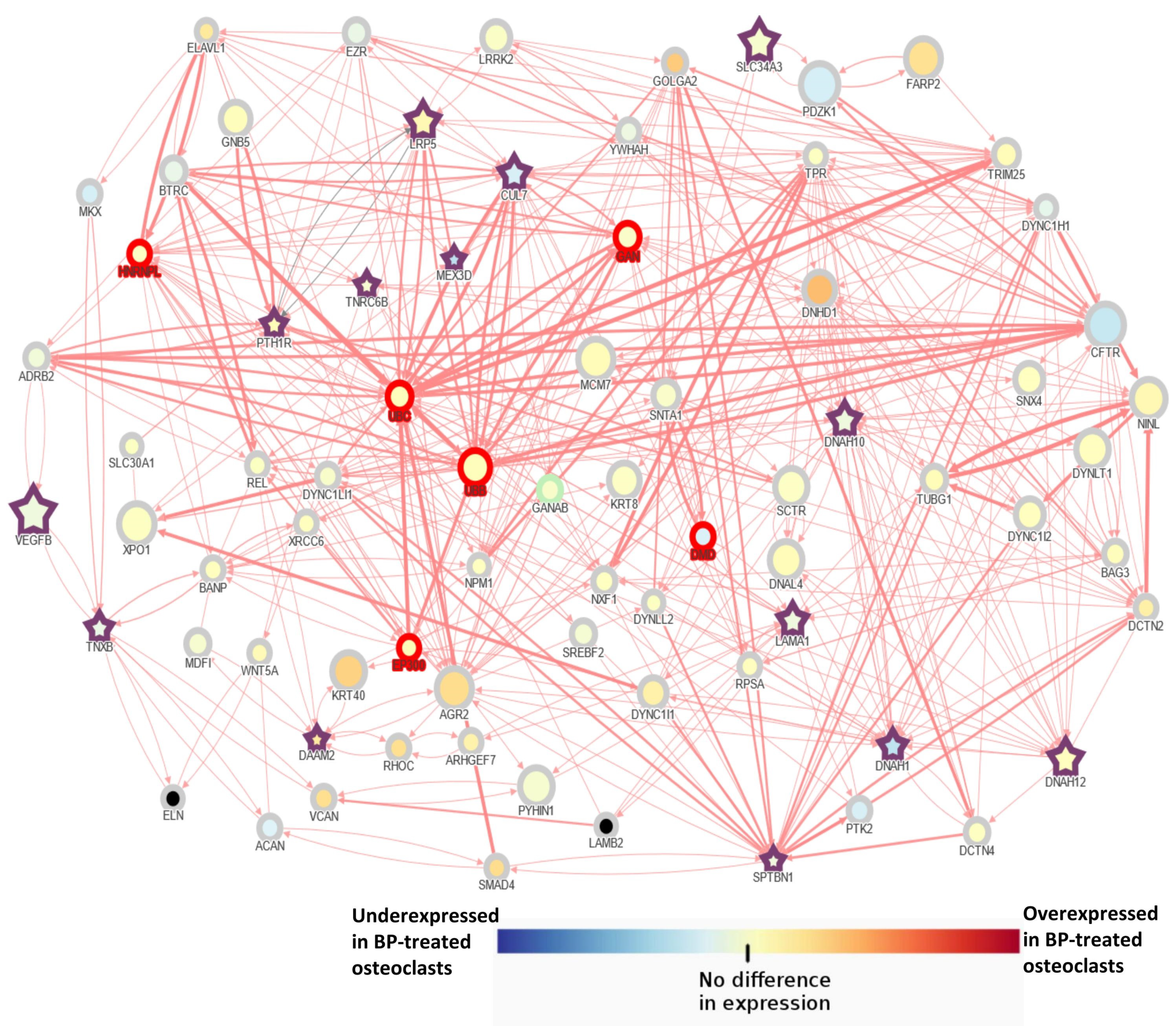
3.2. AFF Network Analysis with Candidate Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shane, E.; Burr, D.; Abrahamsen, B.; Adler, R.A.; Brown, T.D.; Cheung, A.M.; Cosman, F.; Curtis, J.R.; Dell, R.; Dempster, D.W.; et al. Atypical subtrochanteric and diaphyseal femoral fractures: Second report of a task force of the American Society for Bone and Mineral Research. J. Bone Miner. Res. 2014, 29, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Lo, J.C.; Grimsrud, C.D.; Ott, S.M.; Chandra, M.; Hui, R.L.; Ettinger, B. Atypical femur fracture incidence in women increases with duration of bisphosphonate exposure. Osteoporos. Int. 2019, 30, 2515–2520. [Google Scholar] [CrossRef]
- Starr, J.; Tay, Y.K.D.; Shane, E. Current Understanding of Epidemiology, Pathophysiology, and Management of Atypical Femur Fractures. Curr. Osteoporos Rep. 2018, 16, 519–529. [Google Scholar] [CrossRef]
- Lo, J.C.; Hui, R.L.; Grimsrud, C.D.; Chandra, M.; Neugebauer, R.S.; Gonzalez, J.R.; Budayr, A.; Lau, G.; Ettinger, B. The association of race/ethnicity and risk of atypical femur fracture among older women receiving oral bisphosphonate therapy. Bone 2016, 85, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Roca-Ayats, N.; Balcells, S.; Garcia-Giralt, N.; Falco-Mascaro, M.; Martinez-Gil, N.; Abril, J.F.; Urreizti, R.; Dopazo, J.; Quesada-Gomez, J.M.; Nogues, X.; et al. GGPS1 Mutation and Atypical Femoral Fractures with Bisphosphonates. N. Engl. J. Med. 2017, 376, 1794–1795. [Google Scholar] [CrossRef]
- Marini, F.; Falchetti, A.; Silvestri, S.; Bagger, Y.; Luzi, E.; Tanini, A.; Christiansen, C.; Brandi, M.L. Modulatory effect of farnesyl pyrophosphate synthase (FDPS) rs2297480 polymorphism on the response to long-term amino-bisphosphonate treatment in postmenopausal osteoporosis. Curr. Med. Res. Opin. 2008, 24, 2609–2615. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.J.; Choi, J.Y.; Cho, S.W.; Kang, D.; Han, K.O.; Kim, S.W.; Kim, S.Y.; Chung, Y.S.; Shin, C.S. Genetic polymorphism of geranylgeranyl diphosphate synthase (GGSP1) predicts bone density response to bisphosphonate therapy in Korean women. Yonsei Med. J. 2010, 51, 231–238. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Olmos, J.M.; Zarrabeitia, M.T.; Hernandez, J.L.; Sanudo, C.; Gonzalez-Macias, J.; Riancho, J.A. Common allelic variants of the farnesyl diphosphate synthase gene influence the response of osteoporotic women to bisphosphonates. Pharm. J. 2012, 12, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Kharazmi, M.; Michaelsson, K.; Schilcher, J.; Eriksson, N.; Melhus, H.; Wadelius, M.; Hallberg, P. A Genome-Wide Association Study of Bisphosphonate-Associated Atypical Femoral Fracture. Calcif. Tissue Int. 2019, 105, 51–67. [Google Scholar] [CrossRef] [PubMed]
- Marini, F.; Brandi, M.L. Atypical femur fractures: A distinctive tract of adult hypophosphatasia. Clin. Cases Miner. Bone Metab. 2017, 14, 324–328. [Google Scholar] [CrossRef]
- Tournis, S.; Dede, A.D. Osteogenesis imperfecta—A clinical update. Metabolism 2018, 80, 27–37. [Google Scholar] [CrossRef]
- Roca-Ayats, N.; Ng, P.Y.; Garcia-Giralt, N.; Falco-Mascaro, M.; Cozar, M.; Abril, J.F.; Quesada Gomez, J.M.; Prieto-Alhambra, D.; Nogues, X.; Dunford, J.E.; et al. Functional Characterization of a GGPPS Variant Identified in Atypical Femoral Fracture Patients and Delineation of the Role of GGPPS in Bone-Relevant Cell Types. J. Bone Miner. Res. 2018, 33, 2091–2098. [Google Scholar] [CrossRef] [PubMed]
- Perez-Nunez, I.; Perez-Castrillon, J.L.; Zarrabeitia, M.T.; Garcia-Ibarbia, C.; Martinez-Calvo, L.; Olmos, J.M.; Briongos, L.S.; Riancho, J.; Camarero, V.; Munoz Vives, J.M.; et al. Exon array analysis reveals genetic heterogeneity in atypical femoral fractures. A pilot study. Mol. Cell Biochem. 2015, 409, 45–50. [Google Scholar] [CrossRef]
- Shane, E.; Burr, D.; Ebeling, P.R.; Abrahamsen, B.; Adler, R.A.; Brown, T.D.; Cheung, A.M.; Cosman, F.; Curtis, J.R.; Dell, R.; et al. Atypical subtrochanteric and diaphyseal femoral fractures: Report of a task force of the American Society for Bone and Mineral Research. J. Bone Miner. Res. 2010, 25, 2267–2294. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Henikoff, S.; Ng, P.C. Predicting the effects of coding non-synonymous variants on protein function using the SIFT algorithm. Nat. Protoc. 2009, 4, 1073–1081. [Google Scholar] [CrossRef] [PubMed]
- Adzhubei, I.A.; Schmidt, S.; Peshkin, L.; Ramensky, V.E.; Gerasimova, A.; Bork, P.; Kondrashov, A.S.; Sunyaev, S.R. A method and server for predicting damaging missense mutations. Nat. Methods 2010, 7, 248–249. [Google Scholar] [CrossRef]
- Rentzsch, P.; Witten, D.; Cooper, G.M.; Shendure, J.; Kircher, M. CADD: Predicting the deleteriousness of variants throughout the human genome. Nucleic Acids Res. 2019, 47, D886–D894. [Google Scholar] [CrossRef]
- Stark, C.; Breitkreutz, B.J.; Reguly, T.; Boucher, L.; Breitkreutz, A.; Tyers, M. BioGRID: A general repository for interaction datasets. Nucleic Acids Res. 2006, 34, D535–D539. [Google Scholar] [CrossRef] [PubMed]
- Chatr-Aryamontri, A.; Breitkreutz, B.J.; Heinicke, S.; Boucher, L.; Winter, A.; Stark, C.; Nixon, J.; Ramage, L.; Kolas, N.; O’Donnell, L.; et al. The BioGRID interaction database: 2013 update. Nucleic Acids Res. 2013, 41, D816–D823. [Google Scholar] [CrossRef] [PubMed]
- Franceschini, A.; Szklarczyk, D.; Frankild, S.; Kuhn, M.; Simonovic, M.; Roth, A.; Lin, J.; Minguez, P.; Bork, P.; von Mering, C.; et al. STRING v9.1: Protein-protein interaction networks, with increased coverage and integration. Nucleic Acids Res. 2013, 41, D808–D815. [Google Scholar] [CrossRef]
- Lotia, S.; Montojo, J.; Dong, Y.; Bader, G.D.; Pico, A.R. Cytoscape app store. Bioinformatics 2013, 29, 1350–1351. [Google Scholar] [CrossRef]
- Yuen, T.; Stachnik, A.; Iqbal, J.; Sgobba, M.; Gupta, Y.; Lu, P.; Colaianni, G.; Ji, Y.; Zhu, L.L.; Kim, S.M.; et al. Bisphosphonates inactivate human EGFRs to exert antitumor actions. Proc. Natl. Acad. Sci. USA 2014, 111, 17989–17994. [Google Scholar] [CrossRef]
- Gentleman, R.C.; Carey, V.J.; Bates, D.M.; Bolstad, B.; Dettling, M.; Dudoit, S.; Ellis, B.; Gautier, L.; Ge, Y.; Gentry, J.; et al. Bioconductor: Open software development for computational biology and bioinformatics. Genome Biol. 2004, 5, R80. [Google Scholar] [CrossRef]
- Morris, J.A.; Kemp, J.P.; Youlten, S.E.; Laurent, L.; Logan, J.G.; Chai, R.C.; Vulpescu, N.A.; Forgetta, V.; Kleinman, A.; Mohanty, S.T.; et al. An atlas of genetic influences on osteoporosis in humans and mice. Nat. Genet. 2019, 51, 258–266. [Google Scholar] [CrossRef]
- Nguyen, H.H.; van de Laarschot, D.M.; Verkerk, A.; Milat, F.; Zillikens, M.C.; Ebeling, P.R. Genetic Risk Factors for Atypical Femoral Fractures (AFFs): A Systematic Review. JBMR Plus 2018, 2, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Koromani, F.; Trajanoska, K.; Rivadeneira, F.; Oei, L. Recent Advances in the Genetics of Fractures in Osteoporosis. Front. Endocrinol 2019, 10, 337. [Google Scholar] [CrossRef] [PubMed]
- Karasik, D.; Zillikens, M.C.; Hsu, Y.H.; Aghdassi, A.; Akesson, K.; Amin, N.; Barroso, I.; Bennett, D.A.; Bertram, L.; Bochud, M.; et al. Disentangling the genetics of lean mass. Am. J. Clin. Nutr. 2019, 109, 276–287. [Google Scholar] [CrossRef]
- Black, D.M.; Geiger, E.J.; Eastell, R.; Vittinghoff, E.; Li, B.H.; Ryan, D.S.; Dell, R.M.; Adams, A.L. Atypical Femur Fracture Risk versus Fragility Fracture Prevention with Bisphosphonates. N. Engl. J. Med. 2020, 383, 743–753. [Google Scholar] [CrossRef]
- Weinstein, R.S. Glucocorticoid-induced osteoporosis. Rev. Endocr. Metab. Disord. 2001, 2, 65–73. [Google Scholar] [CrossRef]
- Khow, K.S.; Yong, T.Y. Atypical femoral fracture in a patient treated with denosumab. J. Bone Miner. Metab. 2015, 33, 355–358. [Google Scholar] [CrossRef] [PubMed]
- Williams, B.O. LRP5: From bedside to bench to bone. Bone 2017, 102, 26–30. [Google Scholar] [CrossRef]
- Alonso, N.; Soares, D.C.; McCloskey, E.V.; Summers, G.D.; Ralston, S.H.; Gregson, C.L. Atypical femoral fracture in osteoporosis pseudoglioma syndrome associated with two novel compound heterozygous mutations in LRP5. J. Bone Miner. Res. 2015, 30, 615–620. [Google Scholar] [CrossRef] [PubMed]
- Uehara, S.; Udagawa, N.; Kobayashi, Y. Regulation of osteoclast function via Rho-Pkn3-c-Src pathways. J. Oral Biosci. 2019, 61, 135–140. [Google Scholar] [CrossRef]
- Tang, A.R.; Hinz, L.E.; Khan, A.; Kline, G.A. Phosphate matters when investigating hypercalcemia: A mutation in SLC34A3 causing HHRH. Endocrinol Diabetes Metab. Case Rep. 2019, 2019, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Kajitani, N.; Yamada, T.; Kawakami, K.; Matsumoto, K.I. TNX deficiency results in bone loss due to an increase in multinucleated osteoclasts. Biochem. Biophys. Res. Commun. 2019, 512, 659–664. [Google Scholar] [CrossRef] [PubMed]
- Chinipardaz, Z.; Liu, M.; Graves, D.T.; Yang, S. Role of Primary Cilia in Bone and Cartilage. J. Dent. Res. 2021, 220345211046606. [Google Scholar] [CrossRef]
- Blangy, A.; Bompard, G.; Guerit, D.; Marie, P.; Maurin, J.; Morel, A.; Vives, V. The osteoclast cytoskeleton—current understanding and therapeutic perspectives for osteoporosis. J. Cell Sci. 2020, 133, jcs244798. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| With AFF N = 12 | No AFF N = 4 | p-Value | |
|---|---|---|---|
| Mean age (years ± SD) | 74.5 ± 6.1 | 79 ± 7.2 | NS |
| BMI ± SD | 29.7 ± 4.6 | 25.3 ± 3.3 | NS |
| Time on BP (Years ± SD) | 9.1 ± 4.4 | 9 ± 2.7 | NS |
| Denosumab (n) | 2 | 1 | NS |
| Corticosteroid treatment (n) | 6 | 0 | <0.05 |
| AFF Bilateral (n) | 3 | - | - |
| Previous OP fractures (n) | 8 | 2 | NS |
| Genes with Rare Variants in Two AFF Cases | Genes with Rare Variants in More Than two AFF Cases | ||||
|---|---|---|---|---|---|
| Two Different Variants | One Variant | ||||
| Gene Name | Gene Name | Gene Name | Gene Name | Gene Name | Number of Variants and (Carriers) |
| AASS | DNAH10 | PSD3 | ACADL | C8orf46 | 1 (3) |
| ABCA10 | DNAH12 | PTH1R | C1orf87 | CHRNG | 3 (3) |
| ABCA4 | DNAH6 | PYHIN1 | CD1A | DAAM2 | 3 (3, one homoz) |
| ABL2 | DYSF | R3HDML | CITED4 | DNAH14 | 4 (4) |
| ADAMTS12 | EFHB | RET | GBA | DNAH2 | 3 (3) |
| ANAPC11 | EP400 | RMDN1 | IQSEC3 | DNAH9 | 3 (3) |
| ANK3 | ERCC5 | RNF157 | NSMAF | FSIP2 | 3 (3) |
| ANKRD40 | FAT4 | RNF34 | PPP2R1B | HLA-DRB1 | 2 (4) |
| ARHGEF18 | FBLN7 | RTEL1 | SERPINB2 | HRASLS | 1 (3) |
| ARID1B | FLJ00418 | SCN9A | SPTBN1 | IGFLR1 | 2 (2, one homoz) |
| ASH1L | GBP3 | 5-Sep | SYDE1 | KRT10 | 1 (5) |
| ATAD2 | GPX4 | SH3BP2 | TNFRSF25 | LAMA1 | 3 (3) |
| ATP10B | HK3 | SHROOM4 | TRAPPC2L | LRP5 | 4 (3) |
| BIN1 | HPS6 | SIRT5 | TRIM32 | MRPS12 | 1 (3) |
| C10orf54 | IGFN1 | SLC26A9 | NEB | 4 (4) | |
| C12orf42 | IGSF10 | SLC2A7 | OBSCN | 5 (5) | |
| C14orf159 | IGSF22 | SLC34A3 | TCOF1 | 3 (4) | |
| C17orf107 | KLHL33 | SLC52A2 | TNXB | 3 (3) | |
| C6 | LLGL1 | SPTBN5 | TTN | 8 (8) | |
| C9orf84 | MEX3D | SRCAP | UTRN | 3 (3) | |
| CA9 | MKS1 | TAF15 | VEGFB | 1 (3) | |
| CDC42BPG | MMP20 | TENM4 | ZC3H3 | 3 (3) | |
| CERKL | MSLNL | TJP3 | |||
| CHAMP1 | NOD2 | TMEM143 | |||
| CLCN2 | NUP153 | TNRC6B | |||
| CRYBA1 | OPLAH | TOPORS | |||
| CTSE | PACSIN2 | TSFM | |||
| CUL7 | PARD6B | TTC14 | |||
| CYYR1 | PCDHAC1 | ZNF34 | |||
| DAB2IP | PDE4DIP | ZNF646 | |||
| DAW1 | PISD | ZNF729 | |||
| DHX34 | PLA2G4D | ZSCAN32 | |||
| Gene ID | Number of Carriers | Function | Bone Association | Bibliography Source |
|---|---|---|---|---|
| CUL7 | 2 | A core component of the 3 M complex required to regulate microtubule dynamics and genome integrity | Mutations in this gene produce the 3 m syndrome, which causes skeletal abnormalities | Genecards |
| Daam2 | 3 | Involved in the canonical Wnt signaling, a pathway critical for bone formation and repair | SNPs in this gene are associated with estimated bone mineral density (eBMD). Daam2 knockout mouse showed decreased bone strength | Musculoskeletal Knowledge Portal, Morris et al., 2019 [24] |
| DNAH10 | 2 | Found in cilia and flagella; ATPase activity and microtubule motor activity | SNPs in this gene are associated with waist-hip ratio and eBMD. | Musculoskeletal Knowledge Portal |
| DNAH12 | 2 | ATPase activity and microtubule motor activity | SNPs in this gene are associated with waist-hip ratio and eBMD | Musculoskeletal Knowledge Portal |
| LAMA1 | 3 | A major component of the basal membrane which has been implicated in a wide variety of biological processes including cell adhesion, differentiation, migration, and signaling | Binding to cells via a high affinity receptor, laminin is thought to mediate the attachment, migration and organization of cells into tissues during embryonic development by interacting with other extracellular matrix components. | Genecards |
| LRP5 | 4 | A co-receptor with Frizzled protein family members for transducing signals by Wnt proteins | It plays a key role in skeletal homeostasis and many bone density related diseases are caused by mutations in this gene | Genecards |
| MEX3D | 2 | RNA binding protein, may be involved in post-transcriptional regulatory mechanisms | Found mutated in three sisters with AFF | Roca-Ayats N, et al. 2018 [12] |
| PTH1R | 2 | A receptor for parathyroid hormone (PTH) and for parathyroid hormone-like hormone (PTHLH). | Involved in the Hedgehog and PTH signaling pathways in bone and cartilage development | Genecards |
| SLC34A3 | 2 | Involved in the transporting phosphate into cells via sodium cotransport in the renal brush border membrane, and contributes to the maintenance of inorganic phosphate concentration in the kidney | Mutations in this gene are associated with hereditary hypophosphatemic rickets with hypercalciuria. | Genecards |
| SPTBN1 | 2 | Spectrin is an actin crosslinking and molecular scaffold protein that links the plasma membrane to the actin cytoskeleton, and functions in the determination of cell shape, arrangement of transmembrane proteins, and organization of organelles | SNPs in this gene are associated with eBMD and total body BMD | Musculoskeletal Knowledge Portal |
| TNRC6B | 2 | Involved in cellular senescence, innate or adaptive immune system, Wnt signaling, and calcium modulating pathways | SNPs in this gene are mainly associated with lean mass. One SNP was also associated with lower lumbar spine BMD and increased risk of fractures | Karasik D, et al. 2019 [27] |
| TNXB | 3 | A member of the tenascin family of extracellular matrix glycoproteins | Mutations in this gene are associated with the Ehlers-Danlos Syndrome | Genecards |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garcia-Giralt, N.; Roca-Ayats, N.; Abril, J.F.; Martinez-Gil, N.; Ovejero, D.; Castañeda, S.; Nogues, X.; Grinberg, D.; Balcells, S.; Rabionet, R. Gene Network of Susceptibility to Atypical Femoral Fractures Related to Bisphosphonate Treatment. Genes 2022, 13, 146. https://doi.org/10.3390/genes13010146
Garcia-Giralt N, Roca-Ayats N, Abril JF, Martinez-Gil N, Ovejero D, Castañeda S, Nogues X, Grinberg D, Balcells S, Rabionet R. Gene Network of Susceptibility to Atypical Femoral Fractures Related to Bisphosphonate Treatment. Genes. 2022; 13(1):146. https://doi.org/10.3390/genes13010146
Chicago/Turabian StyleGarcia-Giralt, Natalia, Neus Roca-Ayats, Josep F Abril, Nuria Martinez-Gil, Diana Ovejero, Santos Castañeda, Xavier Nogues, Daniel Grinberg, Susanna Balcells, and Raquel Rabionet. 2022. "Gene Network of Susceptibility to Atypical Femoral Fractures Related to Bisphosphonate Treatment" Genes 13, no. 1: 146. https://doi.org/10.3390/genes13010146
APA StyleGarcia-Giralt, N., Roca-Ayats, N., Abril, J. F., Martinez-Gil, N., Ovejero, D., Castañeda, S., Nogues, X., Grinberg, D., Balcells, S., & Rabionet, R. (2022). Gene Network of Susceptibility to Atypical Femoral Fractures Related to Bisphosphonate Treatment. Genes, 13(1), 146. https://doi.org/10.3390/genes13010146