
Environmental Enrichment Rescues Social Behavioral Deficits and Synaptic Abnormalities in Pten Haploinsufficient Mice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Environmental Enrichment
2.3. Behavioral Assays
2.3.1. General Procedures
2.3.2. Three-Chamber Social Approach
2.3.3. Social Recognition
2.3.4. Marble Burying
2.3.5. Open Field Test
2.4. Post-Mortem Analyses
2.4.1. Brain and Body Mass
2.4.2. Synaptosomal Preparation and Western Blot Analysis
2.5. Statistical Analysis
3. Results
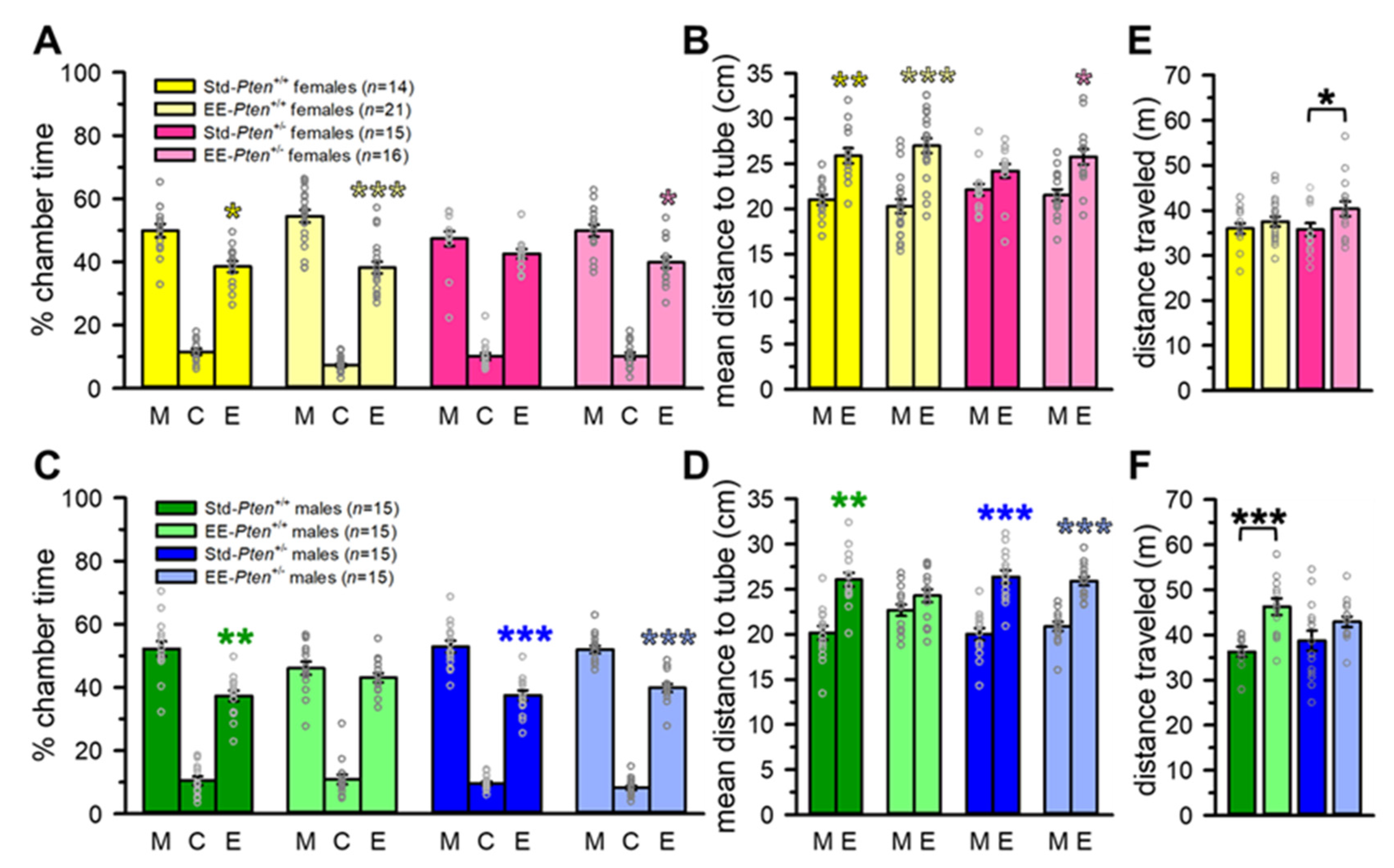
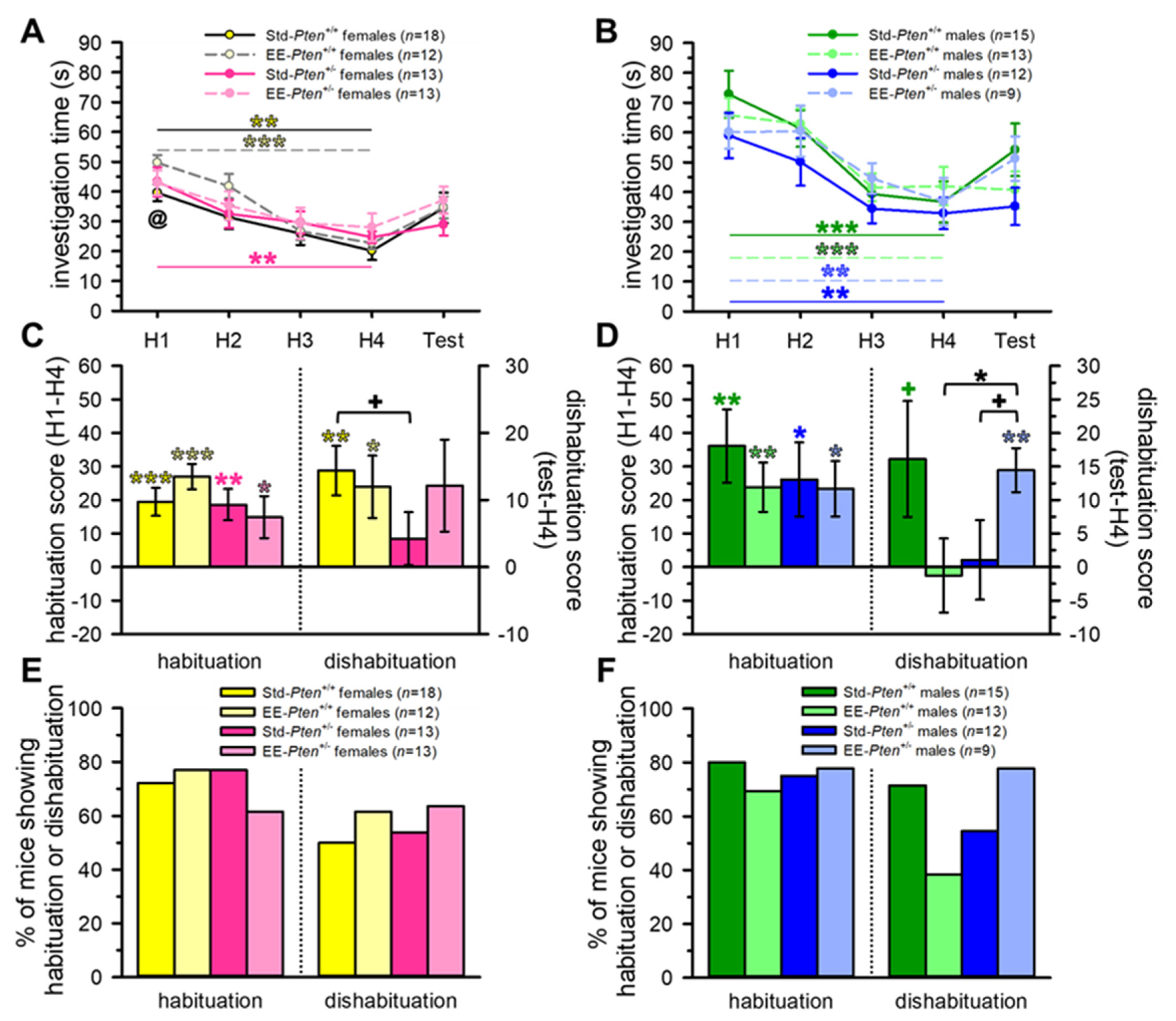
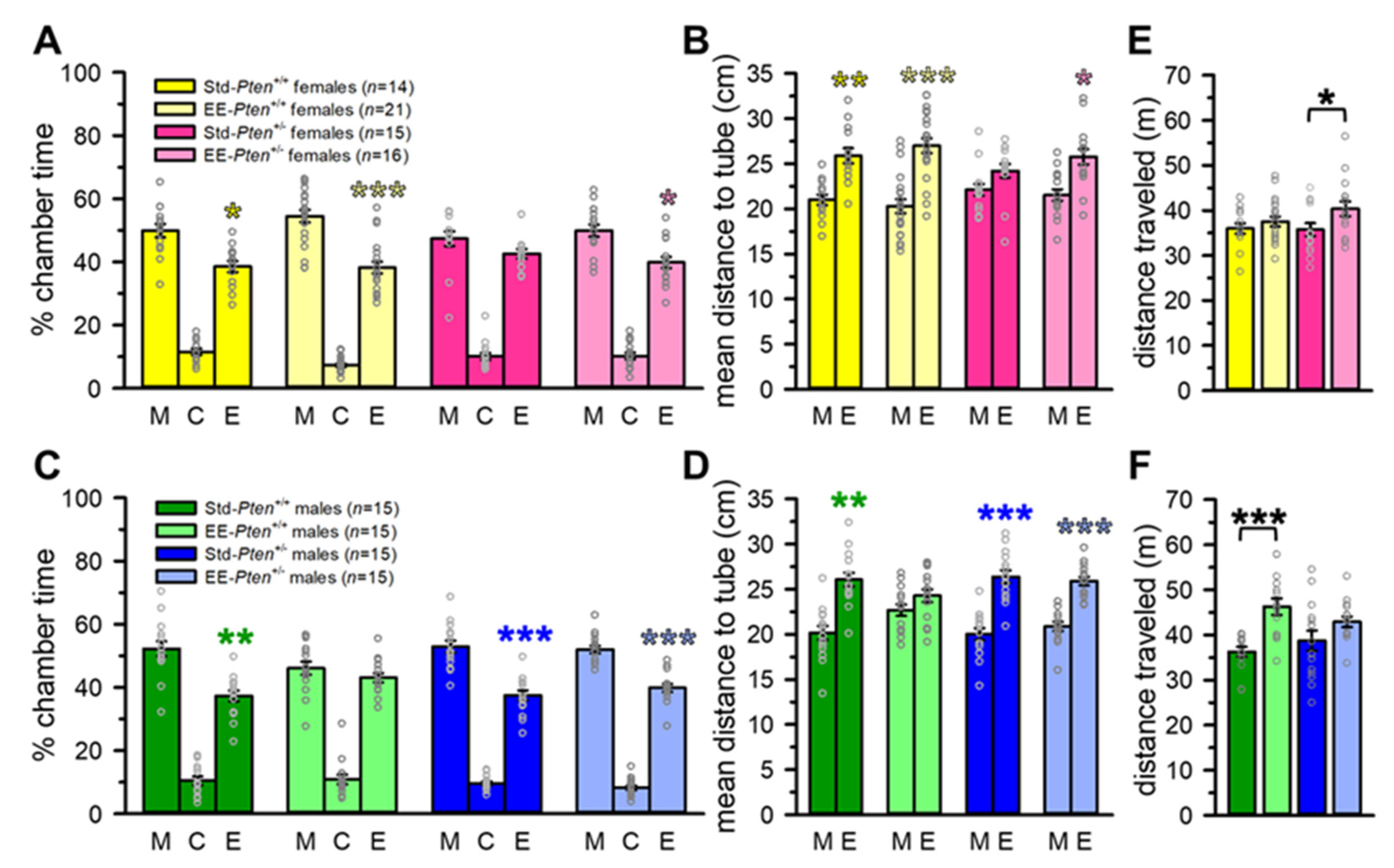
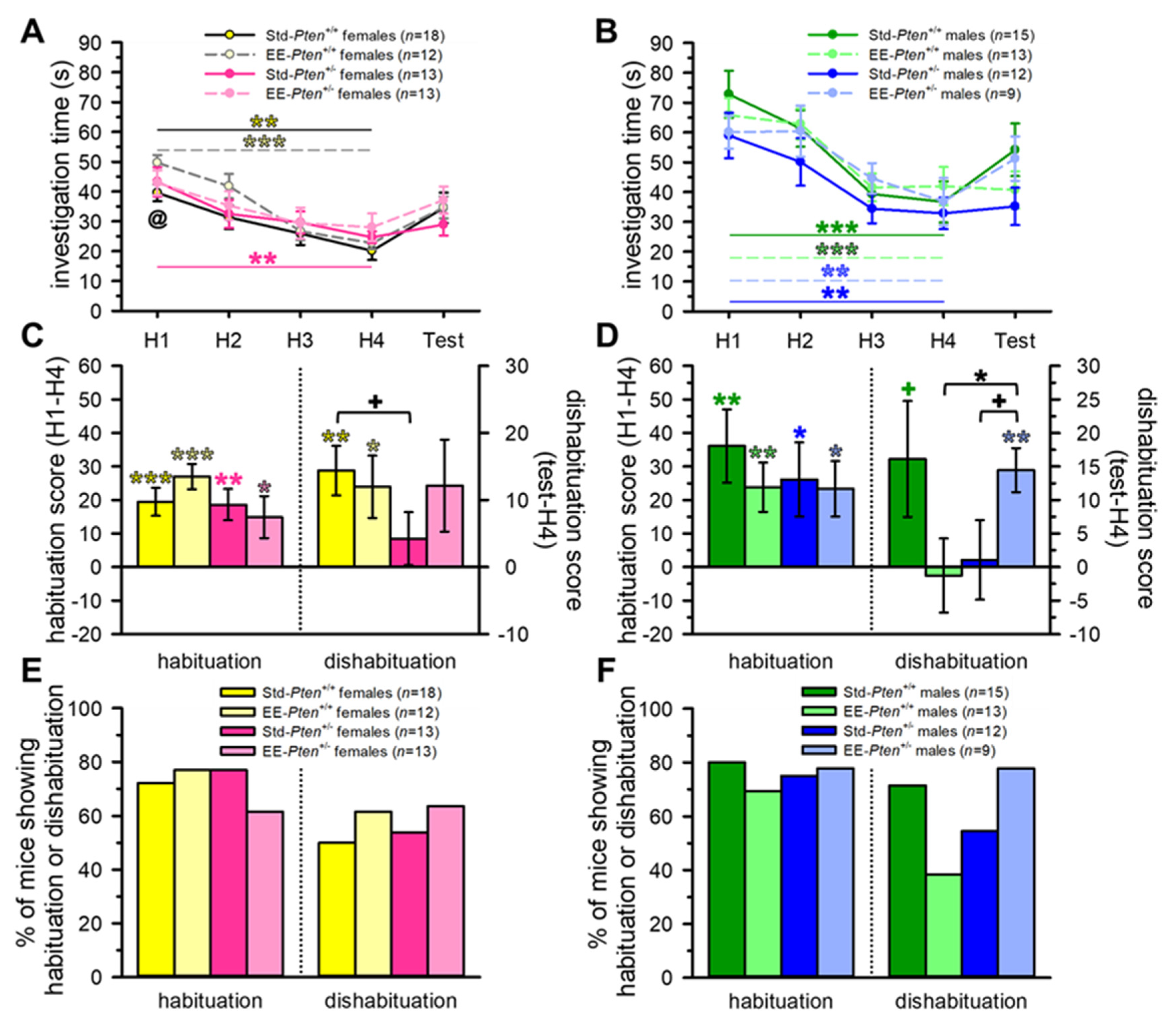
3.1. Environmental Enrichment Rescues Social Behavioral Deficits in Pten+/− Mice
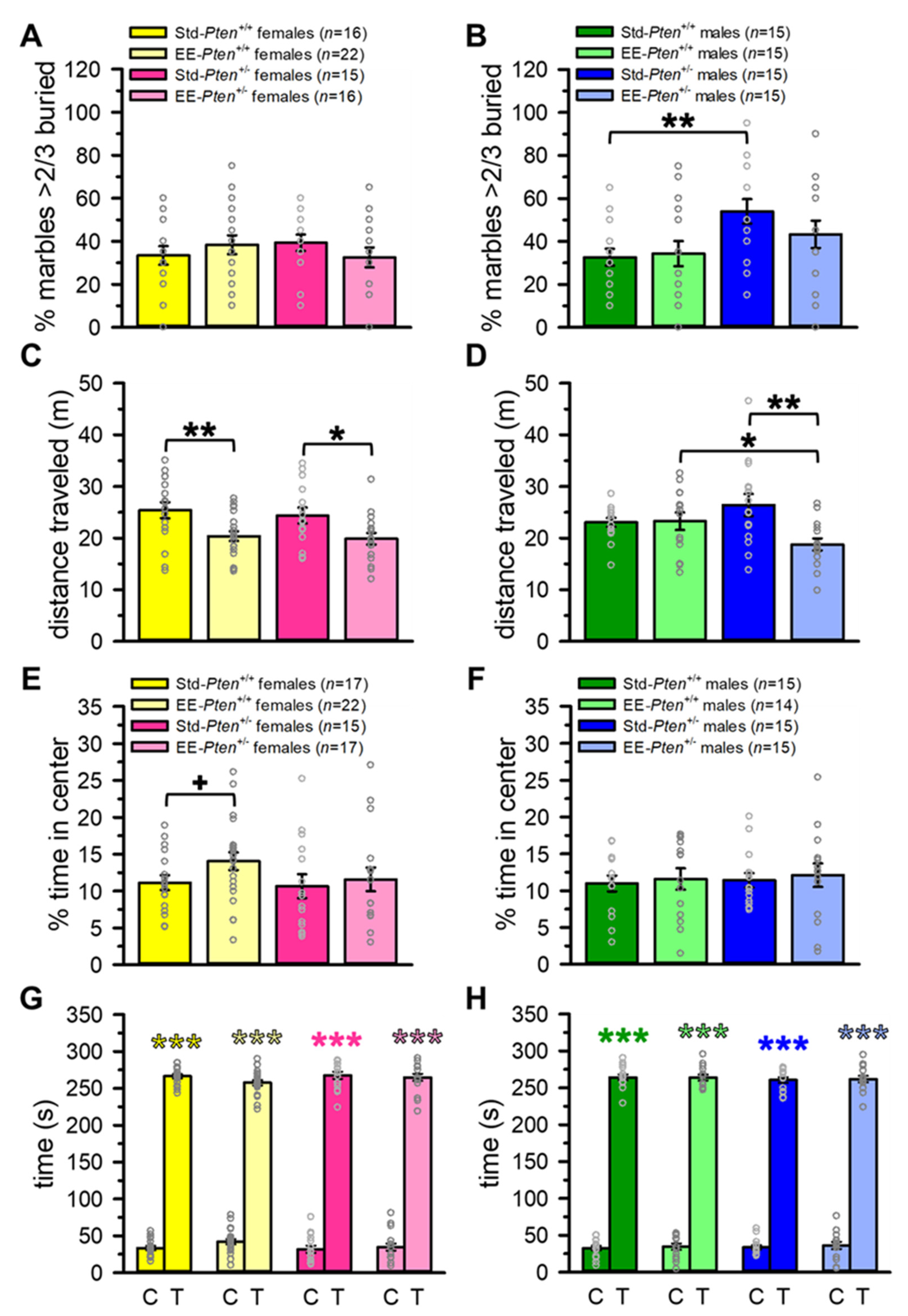
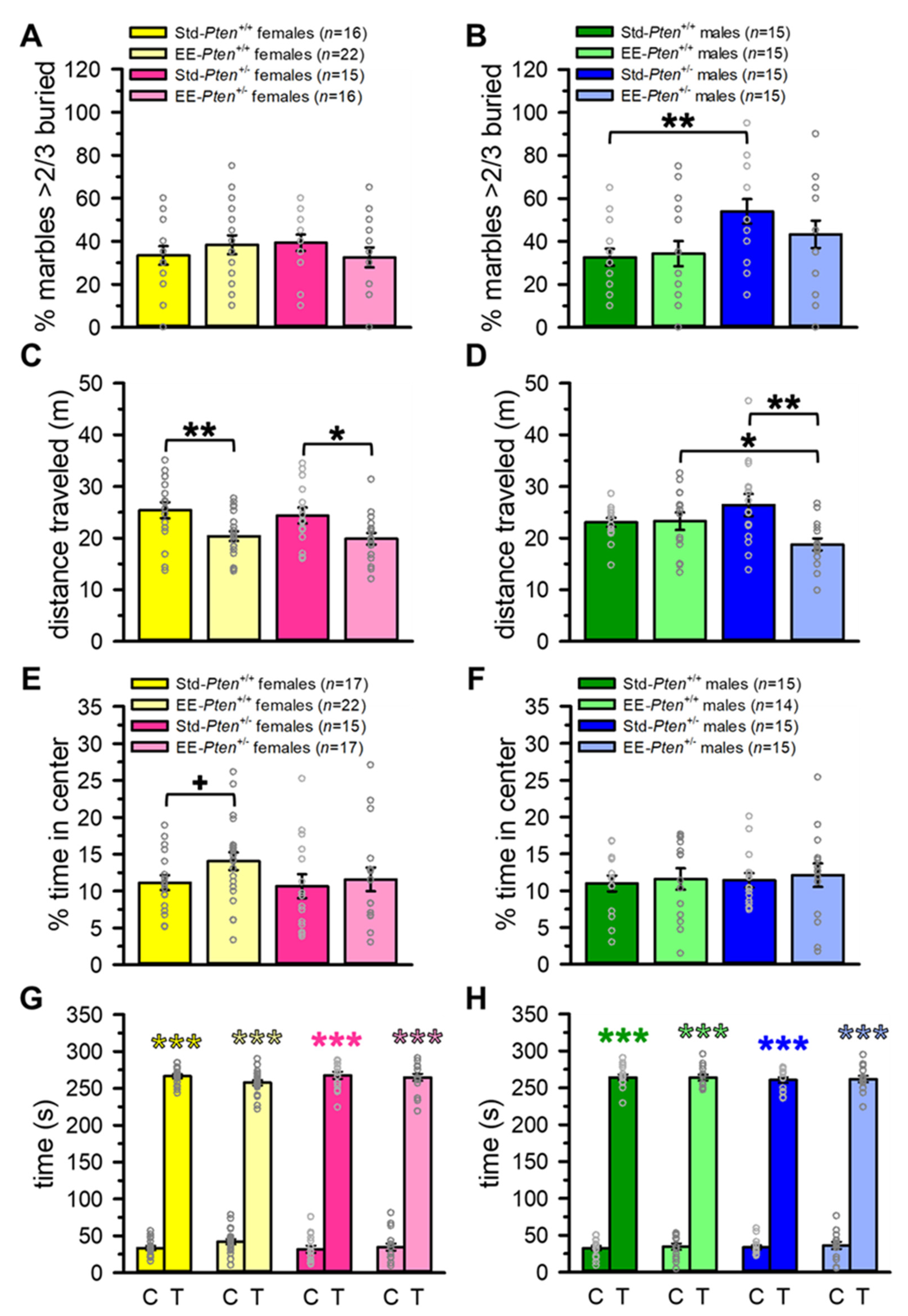
3.2. Environmental Enrichment Partially Rescues ASD-Relevant Repetitive Behavior in Pten+/− Male Mice
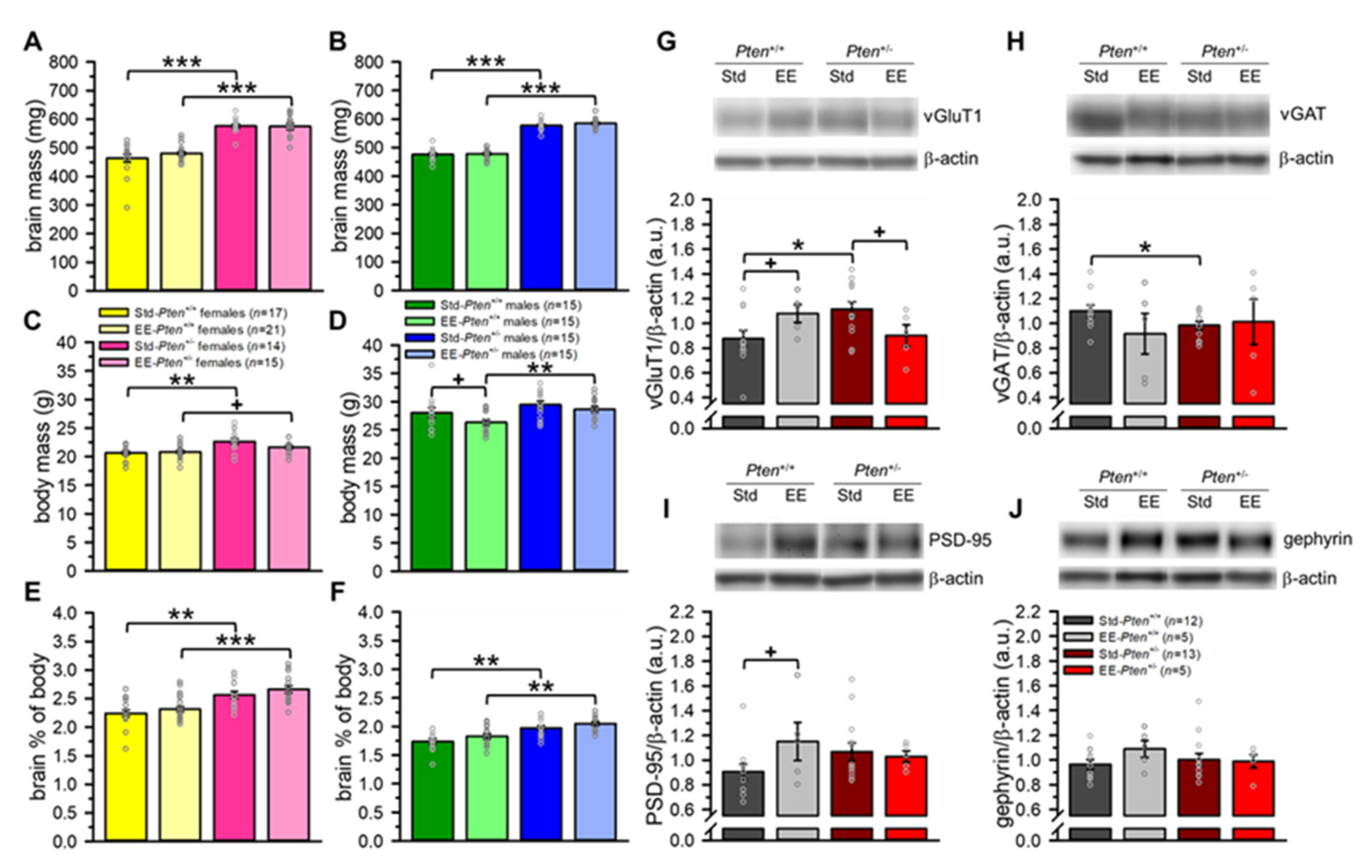
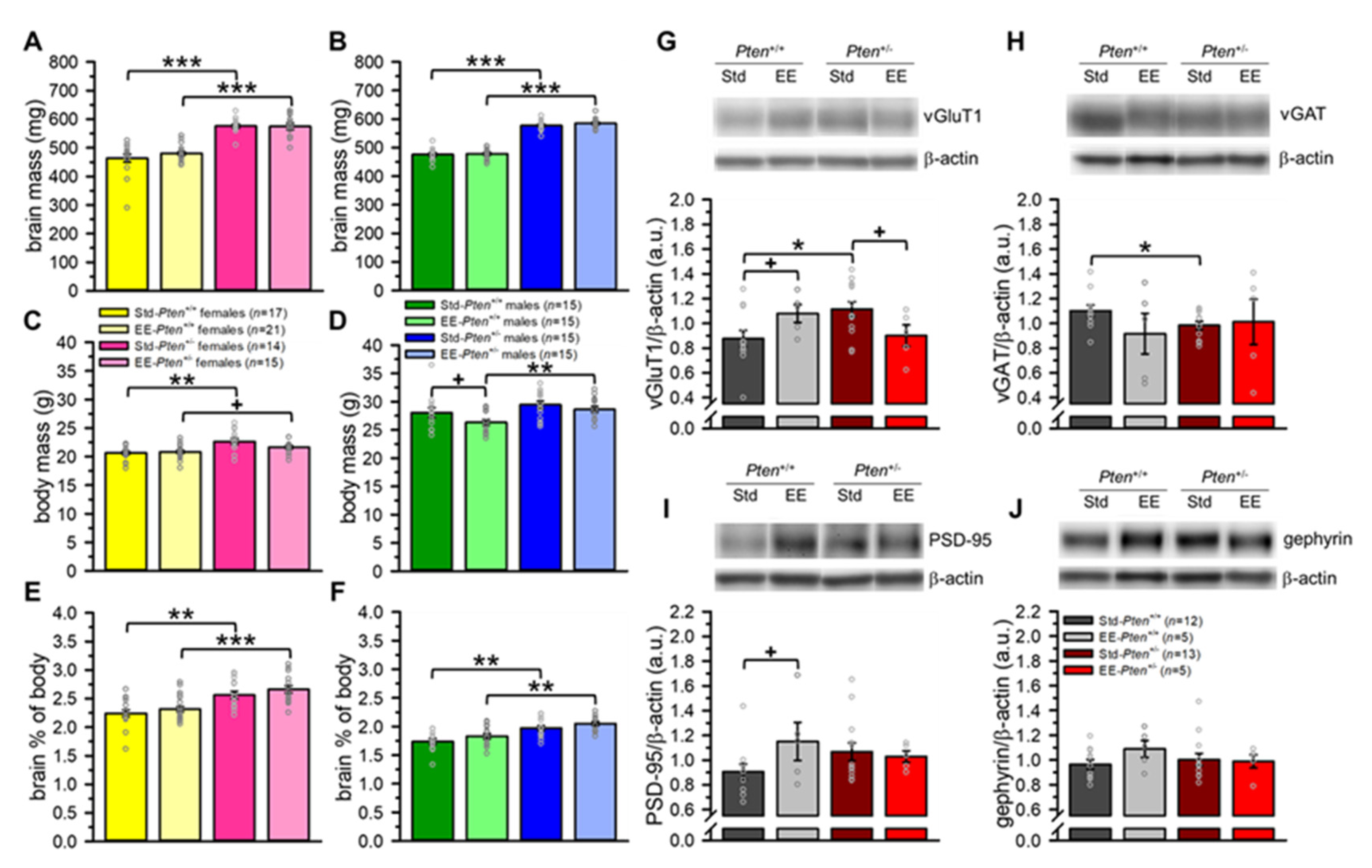
3.3. Environmental Enrichment Rescues Pre-Synaptic Protein Levels, but Does Not Modify Brain Overgrowth in Pten+/− Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Butler, M.; Dasouki, M.; Zhou, X.-P.; Talebizadeh, Z.; Brown, M.; Takahashi, T.; Miles, J.; Wang, C.; Stratton, R.; Pilarski, R.; et al. Subset of individuals with autism spectrum disorders and extreme macrocephaly associated with germline PTEN tumour suppressor gene mutations. J. Med. Genet. 2005, 42, 318–321. [Google Scholar] [CrossRef] [Green Version]
- Buxbaum, J.D.; Cai, G.; Chaste, P.; Nygren, G.; Goldsmith, J.; Reichert, J.; Anckarsäter, H.; Maria, R.; Smith, C.J.; Silverman, J.M.; et al. Mutation screening of the PTEN gene in patients with autism spectrum disorders and macrocephaly. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2007, 144B, 484–491. [Google Scholar] [CrossRef] [Green Version]
- Hobert, J.A.; Embacher, R.; Mester, J.L.; Frazier, T.W., 2nd; Eng, C. Biochemical screening and PTEN mutation analysis in individuals with autism spectrum disorders and macrocephaly. Eur. J. Hum. Genet. 2014, 22, 273–276. [Google Scholar] [CrossRef] [Green Version]
- Klein, S.; Sharifi-Hannauer, P.; Martinez-Agosto, J.A. Macrocephaly as a clinical indicator of genetic subtypes in autism. Autism Res. 2013, 6, 51–56. [Google Scholar] [CrossRef] [PubMed]
- McBride, K.L.; Varga, E.A.; Pastore, M.T.; Prior, T.W.; Manickam, K.; Atkin, J.F.; Herman, G.E. Confirmation study of PTEN mutations among individuals with autism or developmental delays/mental retardation and macrocephaly. Autism Res. 2010, 3, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Varga, E.A.; Pastore, M.; Prior, T.; Herman, G.E.; McBride, K.L. The prevalence of PTEN mutations in a clinical pediatric cohort with autism spectrum disorders, developmental delay, and macrocephaly. Genet. Med. 2009, 11, 111–117. [Google Scholar] [CrossRef] [Green Version]
- Yeung, K.S.; Tso, W.W.Y.; Ip, J.J.K.; Mak, C.C.Y.; Leung, G.K.C.; Tsang, M.H.Y.; Ying, D.; Pei, S.L.C.; Lee, S.L.; Yang, W.; et al. Identification of mutations in the PI3K-AKT-mTOR signalling pathway in patients with macrocephaly and developmental delay and/or autism. Mol. Autism 2017, 8, 66. [Google Scholar] [CrossRef] [PubMed]
- Frazier, T.W.; Embacher, R.; Tilot, A.K.; Koenig, K.; Mester, J.; Eng, C. Molecular and phenotypic abnormalities in individuals with germline heterozygous PTEN mutations and autism. Molec. Psychiat. 2015, 20, 1132–1138. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Escudero, I.; Oliver, M.D.; Andres-Pons, A.; Molina, M.; Cid, V.J.; Pulido, R. A comprehensive functional analysis of PTEN mutations: Implications in tumor- and autism-related syndromes. Hum. Mol. Genet. 2011, 20, 4132–4142. [Google Scholar] [CrossRef] [Green Version]
- Spinelli, L.; Black, F.M.; Berg, J.N.; Eickholt, B.J.; Leslie, N.R. Functionally distinct groups of inherited PTEN mutations in autism and tumour syndromes. J. Med. Genet. 2015, 52, 128–134. [Google Scholar] [CrossRef] [Green Version]
- Boccone, L.; Dessì, V.; Zappu, A.; Piga, S.; Piludu, M.B.; Rais, M.; Massidda, C.; De Virgiliis, S.; Cao, A.; Loudianos, G. Bannayan–Riley–Ruvalcaba syndrome with reactive nodular lymphoid hyperplasia and autism and a PTEN mutation. Am. J. Med. Genet. A 2006, 140A, 1965–1969. [Google Scholar] [CrossRef]
- Conti, S.; Condo, M.; Posar, A.; Mari, F.; Resta, N.; Renieri, A.; Neri, I.; Patrizi, A.; Parmeggiani, A. Phosphatase and tensin homolog (PTEN) gene mutations and autism: Literature review and a case report of a patient with Cowden syndrome, autistic disorder, and epilepsy. J. Child. Neurol. 2012, 27, 392–397. [Google Scholar] [CrossRef]
- Goffin, A.; Hoefsloot, L.H.; Bosgoed, E.; Swillen, A.; Fryns, J.-P. PTEN mutation in a family with Cowden syndrome and autism. Am. J. Med. Genet. 2001, 105, 521–524. [Google Scholar] [CrossRef] [PubMed]
- Clipperton-Allen, A.E.; Page, D.T. Pten haploinsufficient mice show broad brain overgrowth but selective impairments in autism-relevant behavioral tests. Hum. Mol. Genet. 2014, 23, 3490–3505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clipperton-Allen, A.E.; Page, D.T. Decreased aggression and increased repetitive behavior in Pten haploinsufficient mice. Genes Brain Behav. 2015, 14, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Clipperton-Allen, A.E.; Page, D.T. Connecting genotype with behavioral phenotype in mouse models of autism associated with PTEN mutations. Cold Spring Harb Perspect Med. 2020, 10, a037010. [Google Scholar] [CrossRef]
- Page, D.T.; Kuti, O.J.; Prestia, C.; Sur, M. Haploinsufficiency for Pten and Serotonin transporter cooperatively influences brain size and social behavior. Proc. Natl. Acad. Sci. USA 2009, 106, 1989–1994. [Google Scholar] [CrossRef] [Green Version]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders: DSM-5; American Psychiatric Association: Arlington, VA, USA, 2013. [Google Scholar]
- Chen, Y.; Huang, W.C.; Sejourne, J.; Clipperton-Allen, A.E.; Page, D.T. Pten mutations alter brain growth trajectory and allocation of cell types through elevated beta-catenin signaling. J. Neurosci. 2015, 35, 10252–10267. [Google Scholar] [CrossRef]
- Clipperton-Allen, A.E.; Cohen, O.S.; Aceti, M.; Zucca, A.; Levy, J.; Ellegood, J.; Lerch, J.P.; Page, D.T. Pten haploinsufficiency disrupts scaling across brain areas during development in mice. Transl. Psychiatry 2019, 9, 329. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.C.; Chen, Y.; Page, D.T. Hyperconnectivity of prefrontal cortex to amygdala projections in a mouse model of macrocephaly/autism syndrome. Nat. Commun. 2016, 7, 13421. [Google Scholar] [CrossRef]
- Huang, W.C.; Chen, Y.; Page, D.T. Genetic suppression of mTOR rescues synaptic and social behavioral abnormalities in a mouse model of Pten haploinsufficiency. Autism Res. 2019, 12, 1463–1471. [Google Scholar] [CrossRef]
- Loomes, R.; Hull, L.; Mandy, W.P.L. What is the male-to-female ratio in autism spectrum disorder? A systematic review and meta-analysis. J. Am. Acad Child. Adolesc. Psychiatry 2017, 56, 466–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giarelli, E.; Wiggins, L.D.; Rice, C.E.; Levy, S.E.; Kirby, R.S.; Pinto-Martin, J.; Mandell, D. Sex differences in the evaluation and diagnosis of autism spectrum disorders among children. Disabil Health J. 2010, 3, 107–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russell, G.; Steer, C.; Golding, J. Social and demographic factors that influence the diagnosis of autistic spectrum disorders. Soc. Psychiatry Psychiatr. Epidemiol. 2011, 46, 1283–1293. [Google Scholar] [CrossRef]
- Baron-Cohen, S. The extreme male brain theory of autism. Trends Cogn. Sci. 2002, 6, 248–254. [Google Scholar] [CrossRef]
- Jacquemont, S.; Coe, B.P.; Hersch, M.; Duyzend, M.H.; Krumm, N.; Bergmann, S.; Beckmann, J.S.; Rosenfeld, J.A.; Eichler, E.E. A higher mutational burden in females supports a "female protective model" in neurodevelopmental disorders. Am. J. Hum. Genet. 2014, 94, 415–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skuse, D.H. Imprinting, the X-chromosome, and the male brain: Explaining sex differences in the liability to autism. Pediatr. Res. 2000, 47, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Autism Science Foundation. What is Autism? Treatment Options. Available online: https://autismsciencefoundation.org/what-is-autism/treatment-options/ (accessed on 13 April 2021).
- Schreibman, L.; Dawson, G.; Stahmer, A.C.; Landa, R.; Rogers, S.J.; McGee, G.G.; Kasari, C.; Ingersoll, B.; Kaiser, A.P.; Bruinsma, Y.; et al. Naturalistic developmental behavioral interventions: Empirically validated treatments for autism spectrum disorder. J. Autism Dev. Disord. 2015, 45, 2411–2428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lonetti, G.; Angelucci, A.; Morando, L.; Boggio, E.M.; Giustetto, M.; Pizzorusso, T. Early environmental enrichment moderates the behavioral and synaptic phenotype of MeCP2 null mice. Biol. Psychiatry 2010, 67, 657–665. [Google Scholar] [CrossRef] [PubMed]
- Oddi, D.; Subashi, E.; Middei, S.; Bellocchio, L.; Lemaire-Mayo, V.; Guzman, M.; Crusio, W.E.; D’Amato, F.R.; Pietropaolo, S. Early social enrichment rescues adult behavioral and brain abnormalities in a mouse model of fragile X syndrome. Neuropsychopharmacology 2015, 40, 1113–1122. [Google Scholar] [CrossRef] [Green Version]
- More, L.; Lauterborn, J.C.; Papaleo, F.; Brambilla, R. Enhancing cognition through pharmacological and environmental interventions: Examples from preclinical models of neurodevelopmental disorders. Neurosci. Biobehav. Rev. 2020, 110, 28–45. [Google Scholar] [CrossRef]
- Nithianantharajah, J.; Hannan, A.J. Enriched environments, experience-dependent plasticity and disorders of the nervous system. Nat. Rev. Neurosci. 2006, 7, 697–709. [Google Scholar] [CrossRef]
- Huttenrauch, M.; Salinas, G.; Wirths, O. Effects of long-term environmental enrichment on anxiety, memory, hippocampal plasticity and overall brain gene expression in C57BL6 mice. Front. Mol. Neurosci. 2016, 9, 62. [Google Scholar] [CrossRef] [Green Version]
- van Praag, H.; Kempermann, G.; Gage, F.H. Neural consequences of environmental enrichment. Nat. Rev. Neurosci. 2000, 1, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.G.; Toyoda, H.; Wang, Y.K.; Zhuo, M. Enhanced synaptic long-term potentiation in the anterior cingulate cortex of adult wild mice as compared with that in laboratory mice. Mol. Brain 2009, 2, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Podsypanina, K.; Ellenson, L.H.; Nemes, A.; Gu, J.; Tamura, M.; Yamada, K.M.; Cordon-Cardo, C.; Catoretti, G.; Fisher, P.E.; Parsons, R. Mutation of Pten/Mmac1 in mice causes neoplasia in multiple organ systems. Proc. Natl. Acad. Sci. USA 1999, 96, 1563–1568. [Google Scholar] [CrossRef] [Green Version]
- Clipperton-Allen, A.E.; Chen, Y.; Page, D.T. Autism-relevant behaviors are minimally impacted by conditional deletion of Pten in oxytocinergic neurons. Autism Res. 2016, 9, 1248–1262. [Google Scholar] [CrossRef] [PubMed]
- Page, D.T.; Kuti, O.J.; Sur, M. Computerized assessment of social approach behavior in mouse. Front. Behav. Neurosci. 2009, 3, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Takayanagi, Y.; Fujita, E.; Yu, Z.; Yamagata, T.; Momoi, M.Y.; Momoi, T.; Onaka, T. Impairment of social and emotional behaviors in Cadm1-knockout mice. Biochem. Biophys. Res. Commun. 2010, 396, 703–708. [Google Scholar] [CrossRef] [PubMed]
- Thomas, A.; Burant, A.; Bui, N.; Graham, D.; Yuva-Paylor, L.A.; Paylor, R. Marble burying reflects a repetitive and perseverative behavior more than novelty-induced anxiety. Psychopharmacology 2009, 204, 361–373. [Google Scholar] [CrossRef] [Green Version]
- Sejourne, J.; Llaneza, D.; Kuti, O.J.; Page, D.T. Social behavioral deficits coincide with the onset of seizure susceptibility in mice lacking serotonin receptor 2c. PLoS ONE 2015, 10, e0136494. [Google Scholar] [CrossRef] [Green Version]
- Burrows, E.L.; Koyama, L.; May, C.; Hill-Yardin, E.L.; Hannan, A.J. Environmental enrichment modulates affiliative and aggressive social behaviour in the neuroligin-3 R451C mouse model of autism spectrum disorder. Pharmacol. Biochem. Behav. 2020, 195, 172955. [Google Scholar] [CrossRef] [PubMed]
- Cao, M.; Pu, T.; Wang, L.; Marshall, C.; He, H.; Hu, G.; Xiao, M. Early enriched physical environment reverses impairments of the hippocampus, but not medial prefrontal cortex, of socially-isolated mice. Brain Behav. Immun. 2017, 64, 232–243. [Google Scholar] [CrossRef]
- Haemisch, A.; Voss, T.; Gartner, K. Effects of environmental enrichment on aggressive behavior, dominance hierarchies, and endocrine states in male DBA/2J mice. Physiol. Behav. 1994, 56, 1041–1048. [Google Scholar] [CrossRef]
- Mesa-Gresa, P.; Perez-Martinez, A.; Redolat, R. Environmental enrichment improves novel object recognition and enhances agonistic behavior in male mice. Aggress Behav. 2013, 39, 269–279. [Google Scholar] [CrossRef]
- Van Loo, P.L.; Mol, J.A.; Koolhaas, J.M.; Van Zutphen, B.F.; Baumans, V. Modulation of aggression in male mice: Influence of group size and cage size. Physiol. Behav. 2001, 72, 675–683. [Google Scholar] [CrossRef] [Green Version]
- Bouet, V.; Freret, T.; Dutar, P.; Billard, J.M.; Boulouard, M. Continuous enriched environment improves learning and memory in adult NMRI mice through theta burst-related-LTP independent mechanisms but is not efficient in advanced aged animals. Mech. Ageing Dev. 2011, 132, 240–248. [Google Scholar] [CrossRef]
- Burrows, E.L.; McOmish, C.E.; Buret, L.S.; Van den Buuse, M.; Hannan, A.J. Environmental enrichment ameliorates behavioral impairments modeling schizophrenia in mice lacking metabotropic glutamate receptor 5. Neuropsychopharmacology 2015, 40, 1947–1956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinla, I.; Leidmaa, E.; Visnapuu, T.; Philips, M.A.; Vasar, E. Enrichment and individual housing reinforce the differences in aggressiveness and amphetamine response in 129S6/SvEv and C57BL/6 strains. Behav. Brain Res. 2014, 267, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.J.; Zou, R.; Huang, S.; Song, T.J.; Yu, X. Enriched environment rearing from birth reduced anxiety, improved learning and memory, and promoted social interactions in adult male mice. Neuroscience 2020, 442, 138–150. [Google Scholar] [CrossRef] [PubMed]
- Flurkey, K.; Currer, J.M.; Harrison, D.E. Mouse models in aging research. In The Mouse in Biomedical Research, 2nd ed.; Fox, J.G., Davisson, M.T., Quimby, F.W., Barthold, S.W., Newcomer, C.E., Smith, A.L., Eds.; Academic Press: Burlington, MA, USA, 2007; Volume 3, pp. 637–672. [Google Scholar]
- Semple, B.D.; Blomgren, K.; Gimlin, K.; Ferriero, D.M.; Noble-Haeusslein, L.J. Brain development in rodents and humans: Identifying benchmarks of maturation and vulnerability to injury across species. Prog. Neurobiol. 2013, 106–107, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, E.; Lee, J.; Kim, E. Excitation/inhibition imbalance in animal models of autism spectrum disorders. Biol. Psychiatry 2017, 81, 838–847. [Google Scholar] [CrossRef] [PubMed] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Clipperton-Allen, A.E.; Zhang, A.; Cohen, O.S.; Page, D.T. Environmental Enrichment Rescues Social Behavioral Deficits and Synaptic Abnormalities in Pten Haploinsufficient Mice. Genes 2021, 12, 1366. https://doi.org/10.3390/genes12091366
Clipperton-Allen AE, Zhang A, Cohen OS, Page DT. Environmental Enrichment Rescues Social Behavioral Deficits and Synaptic Abnormalities in Pten Haploinsufficient Mice. Genes. 2021; 12(9):1366. https://doi.org/10.3390/genes12091366
Chicago/Turabian StyleClipperton-Allen, Amy E., Angela Zhang, Ori S. Cohen, and Damon Theron Page. 2021. "Environmental Enrichment Rescues Social Behavioral Deficits and Synaptic Abnormalities in Pten Haploinsufficient Mice" Genes 12, no. 9: 1366. https://doi.org/10.3390/genes12091366
APA StyleClipperton-Allen, A. E., Zhang, A., Cohen, O. S., & Page, D. T. (2021). Environmental Enrichment Rescues Social Behavioral Deficits and Synaptic Abnormalities in Pten Haploinsufficient Mice. Genes, 12(9), 1366. https://doi.org/10.3390/genes12091366