
In Vitro Antiplatelet Activity of Mulberroside C through the Up-Regulation of Cyclic Nucleotide Signaling Pathways and Down-Regulation of Phosphoproteins



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Suspension of Human Platelets
2.3. In Vitro Platelet Aggregation Assay
2.4. Cytotoxicity Analysis
2.5. Ca2+ Mobilization Analysis
2.6. Serotonin and ATP Analysis
2.7. Flow Cytometric Determination of P-Selectin Release
2.8. Thromboxane B2 Analysis
2.9. Immunoblotting Assay
2.10. Flow Cytometric Determination of αIIb/β3 Activation
2.11. Measurement of cAMP and cGMP
2.12. Fibrin Clot Retraction
2.13. Data Analyses
3. Results
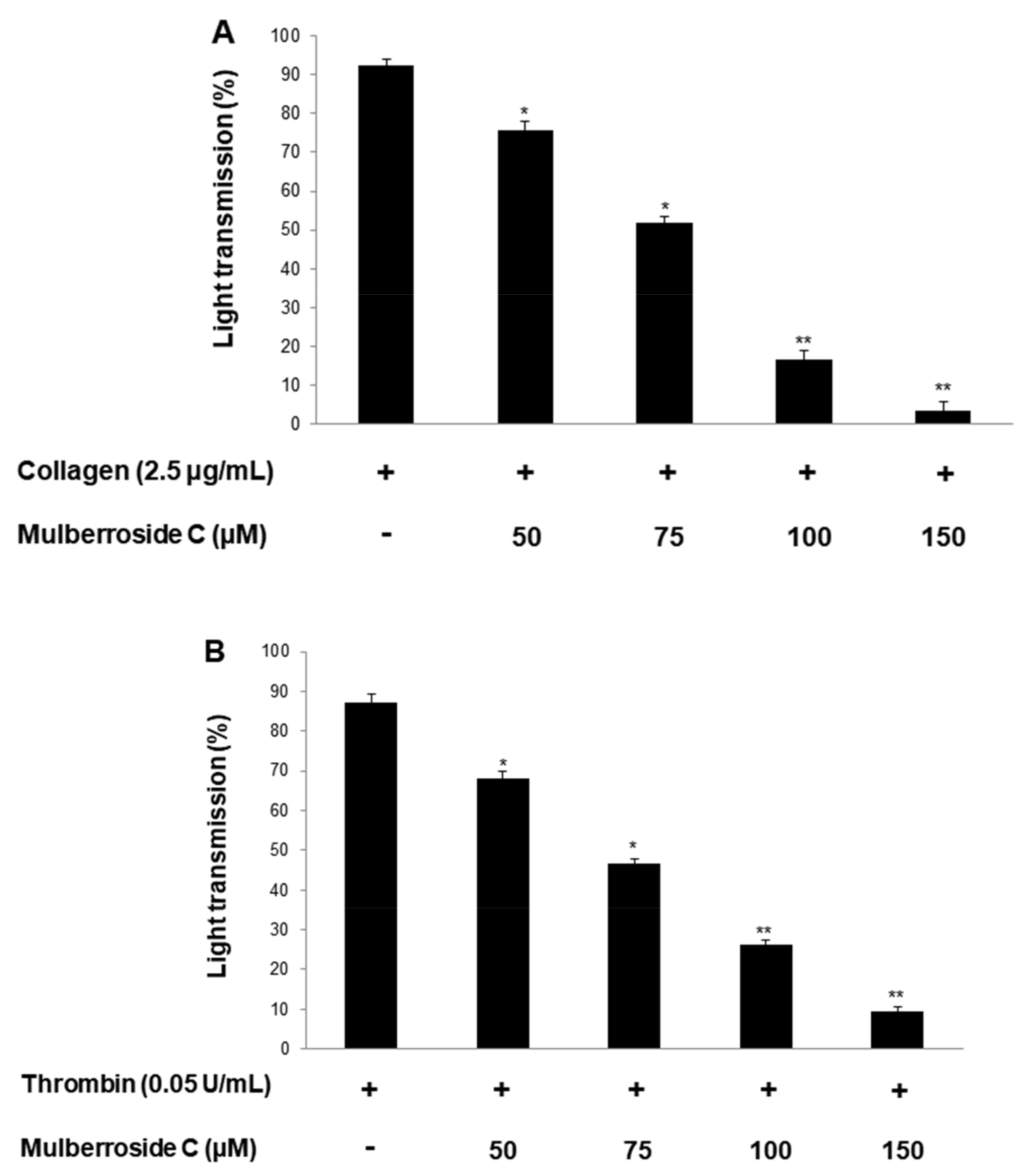
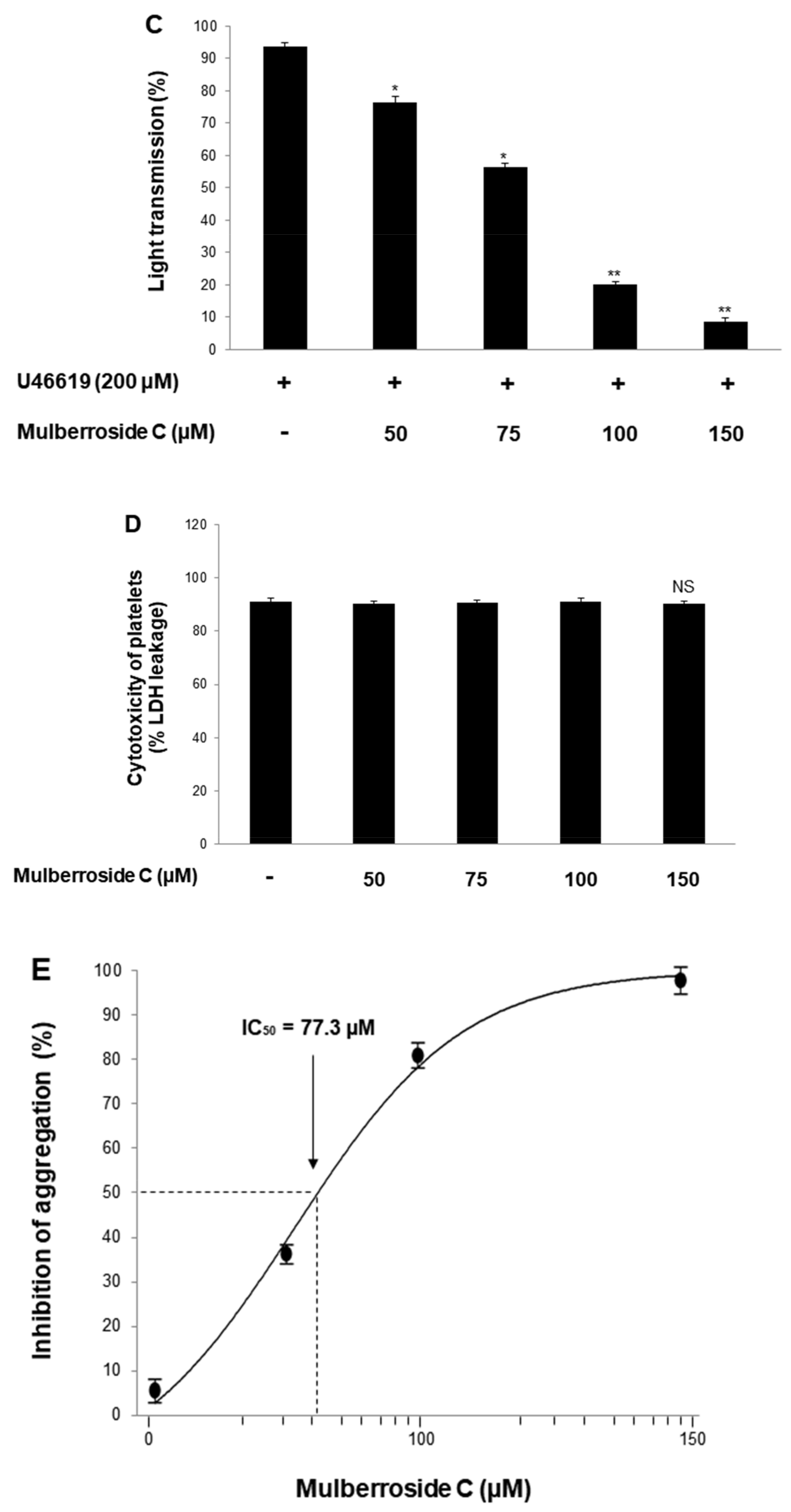
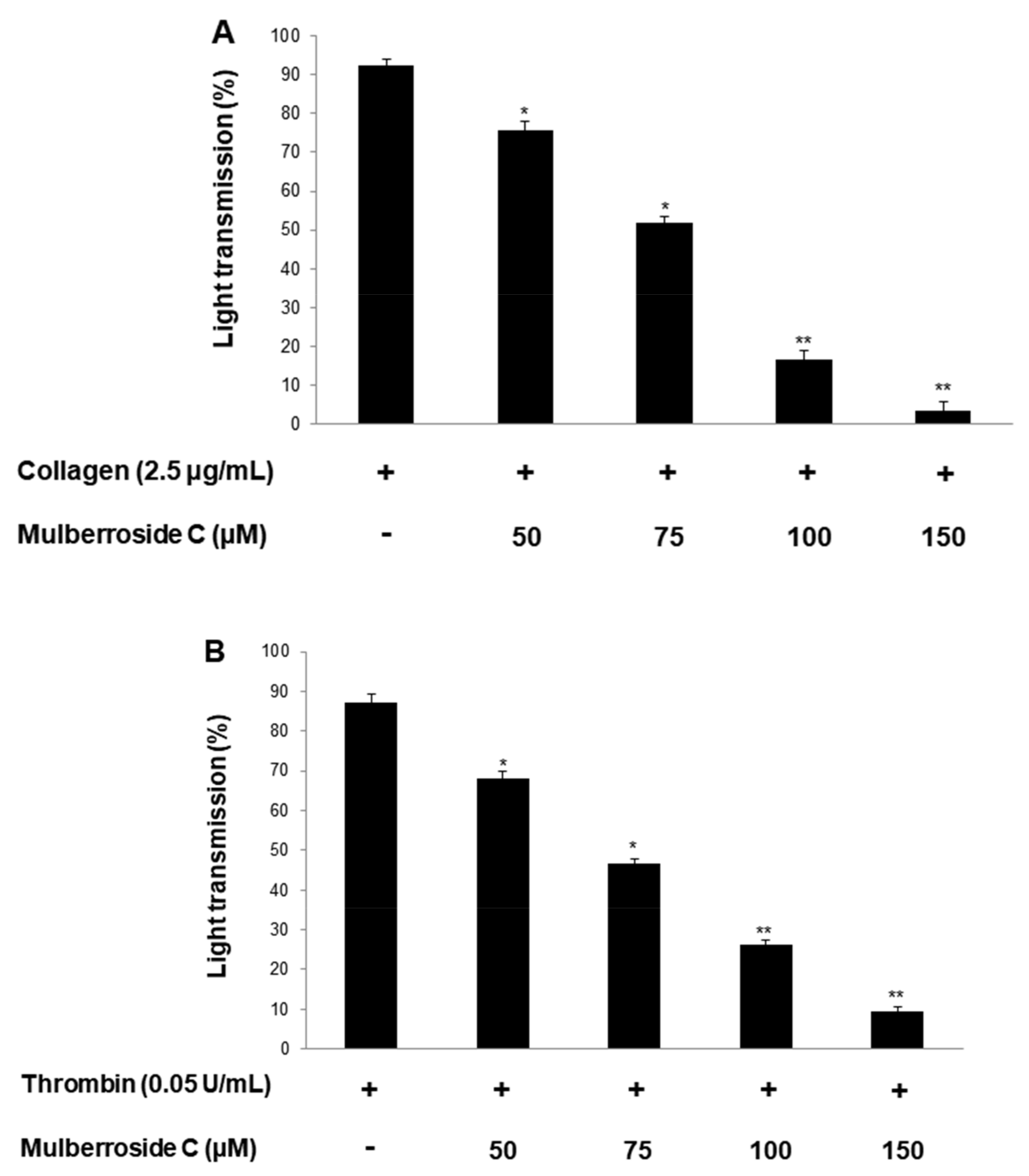
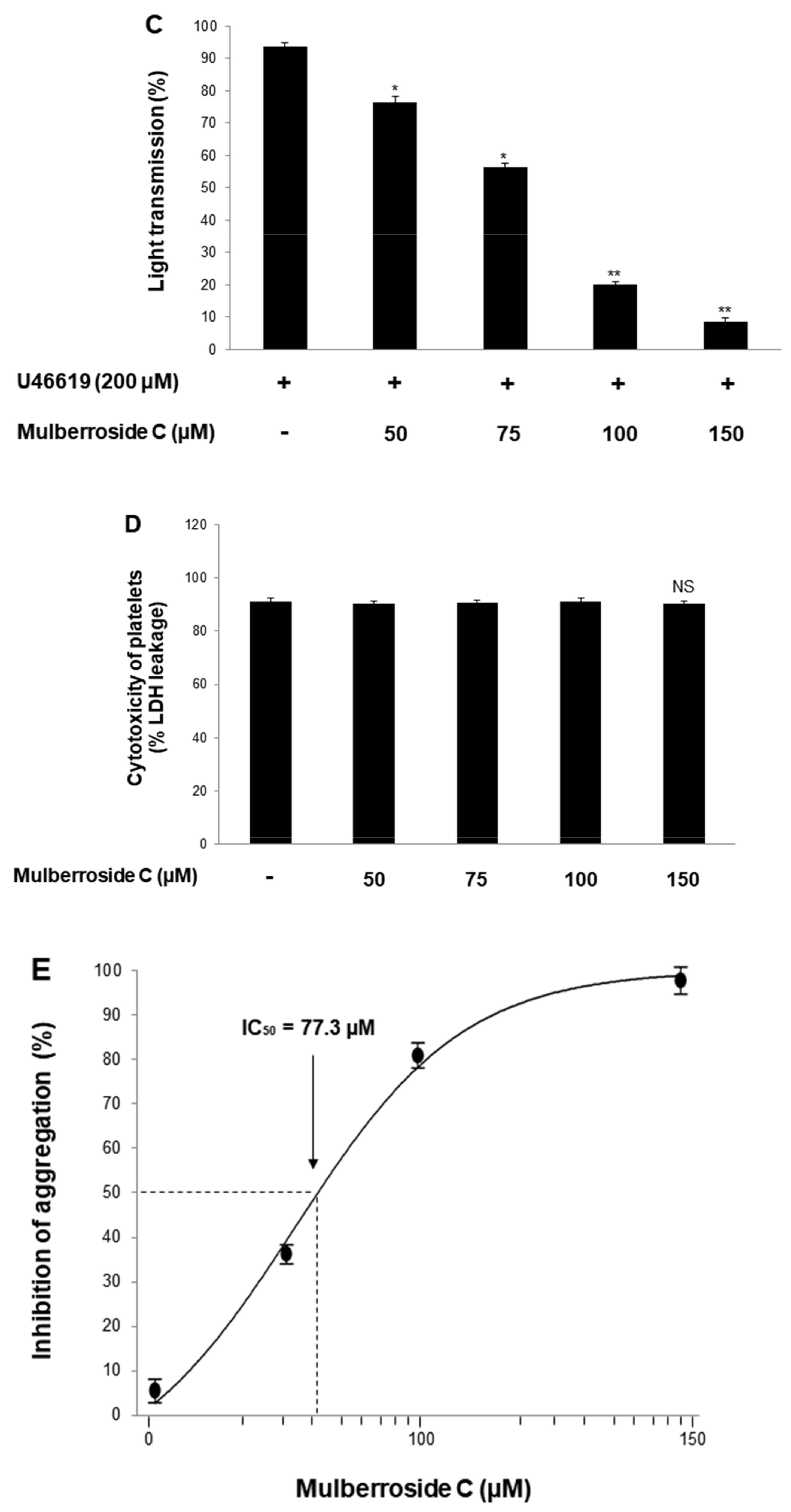
3.1. Mulberroside C Blocks Platelet Aggregaion and Cytotoxicity
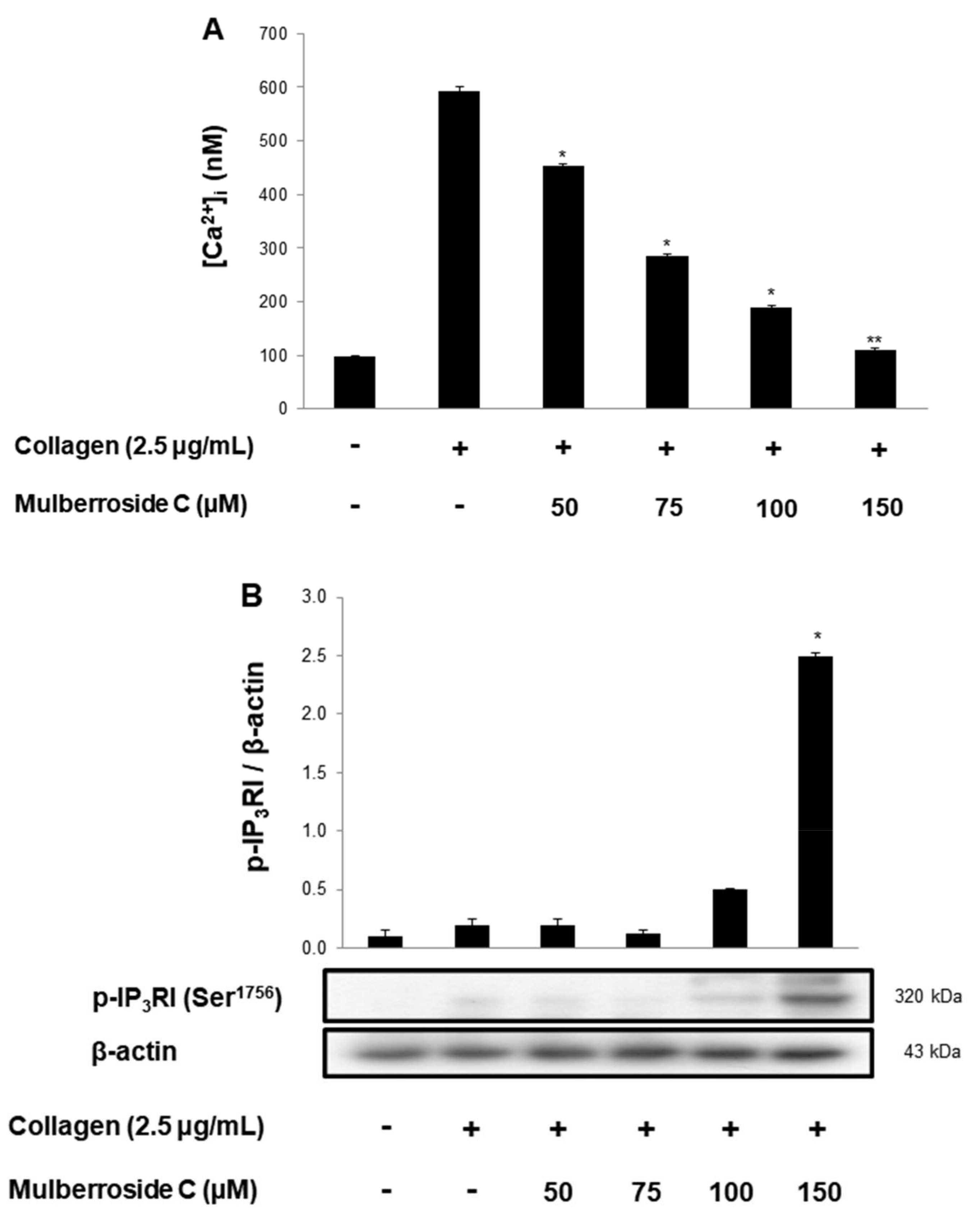
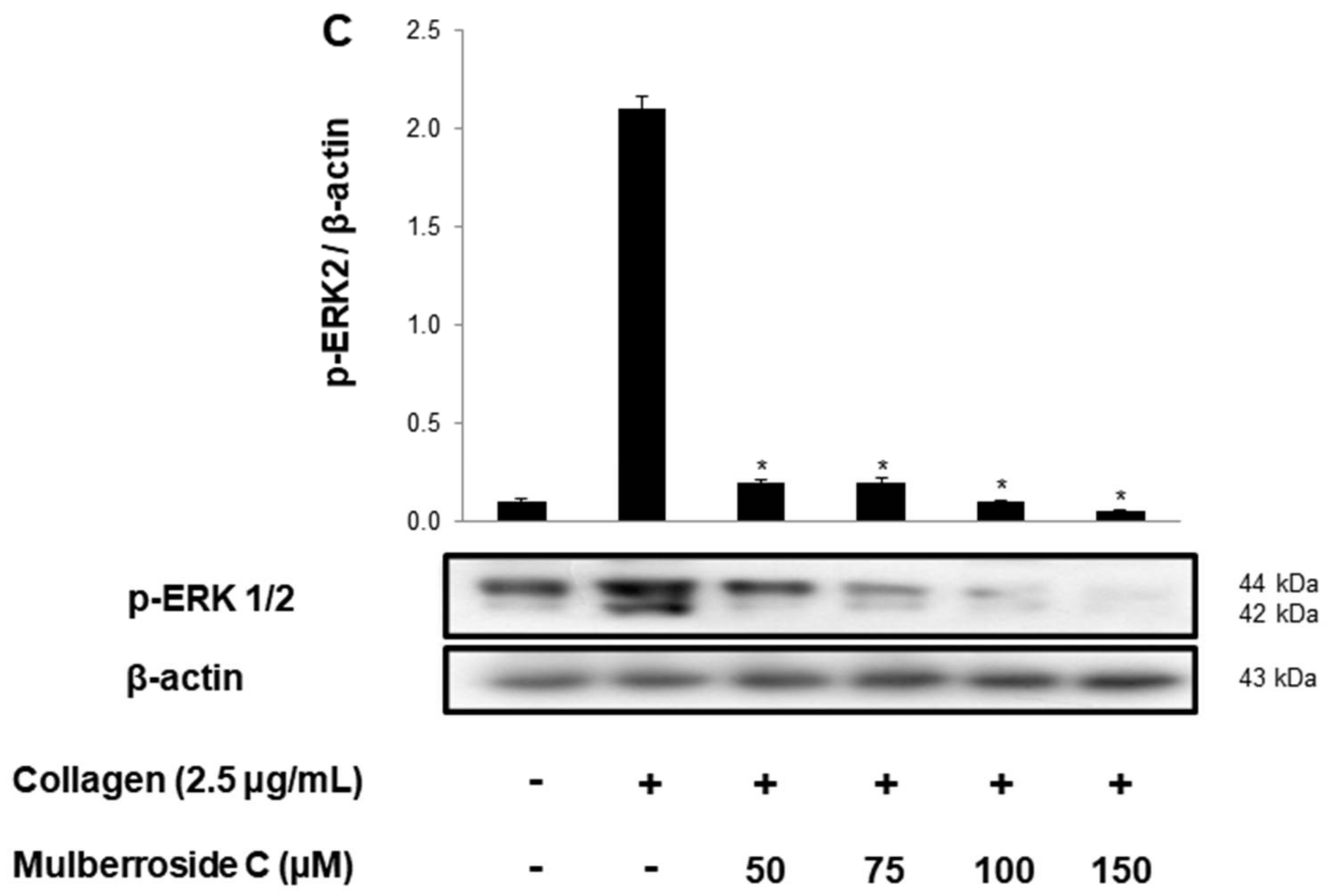
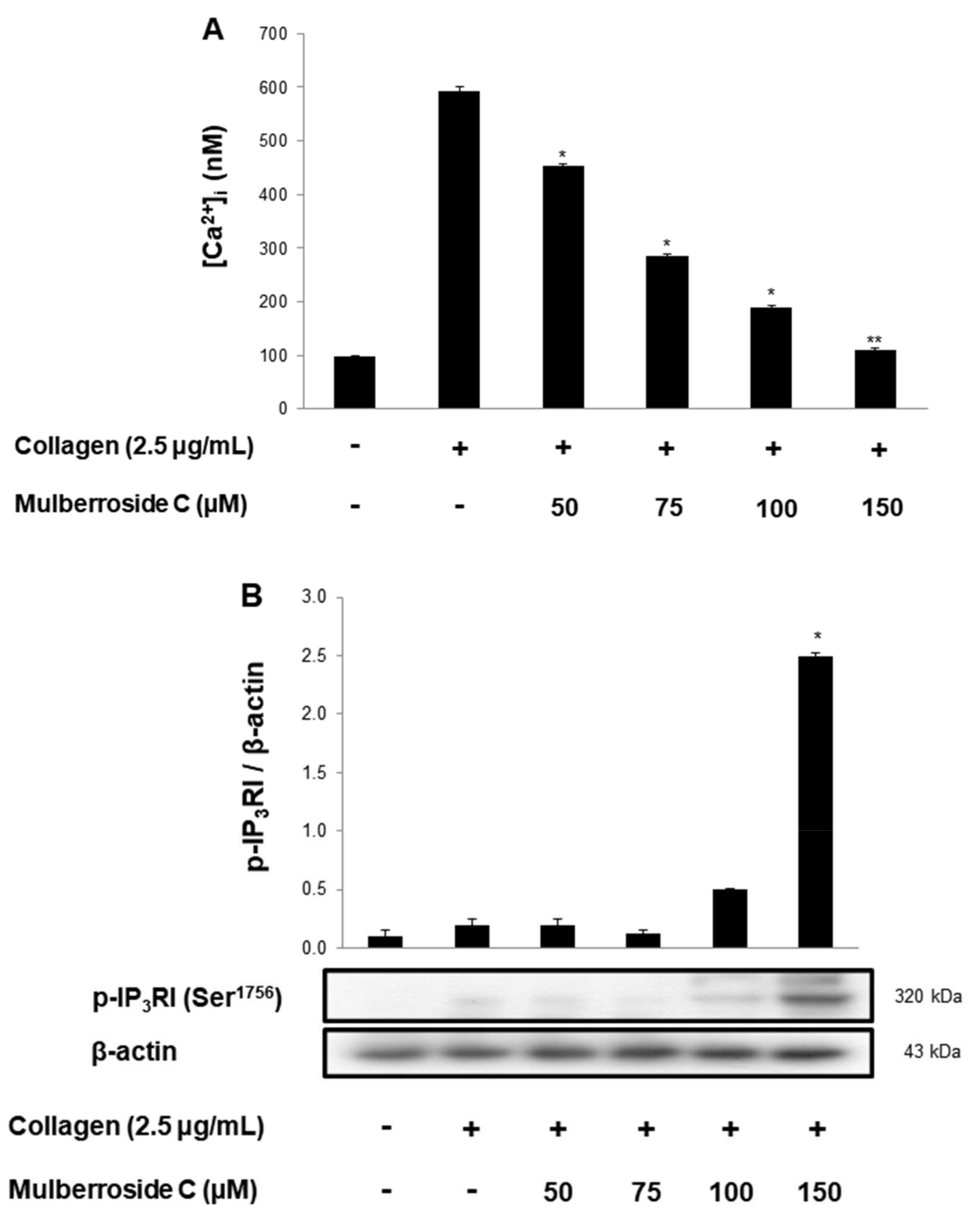
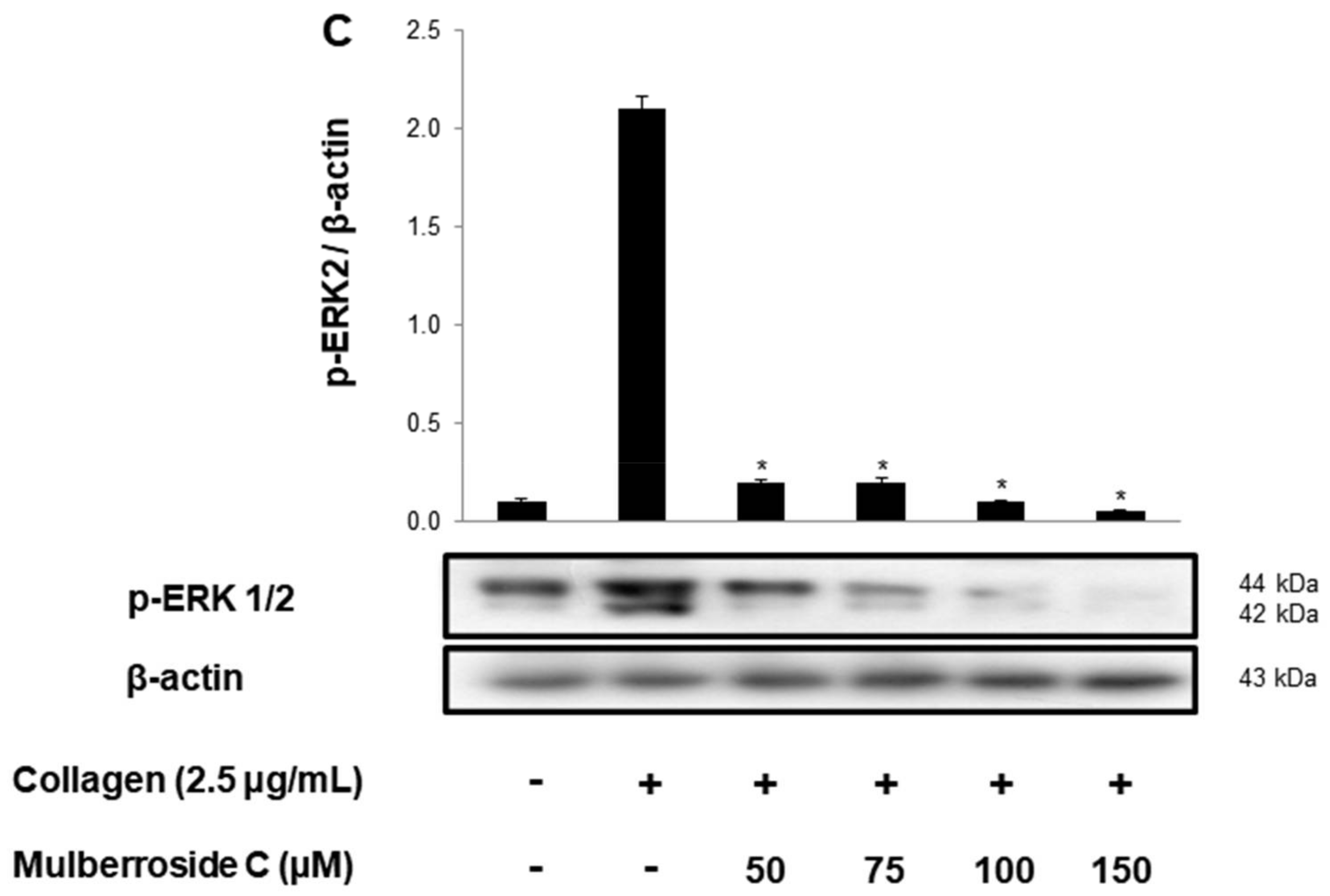
3.2. Mulberroside C Blocks Ca2+ Levels and IP3RI-, ERK-Phosphorylation
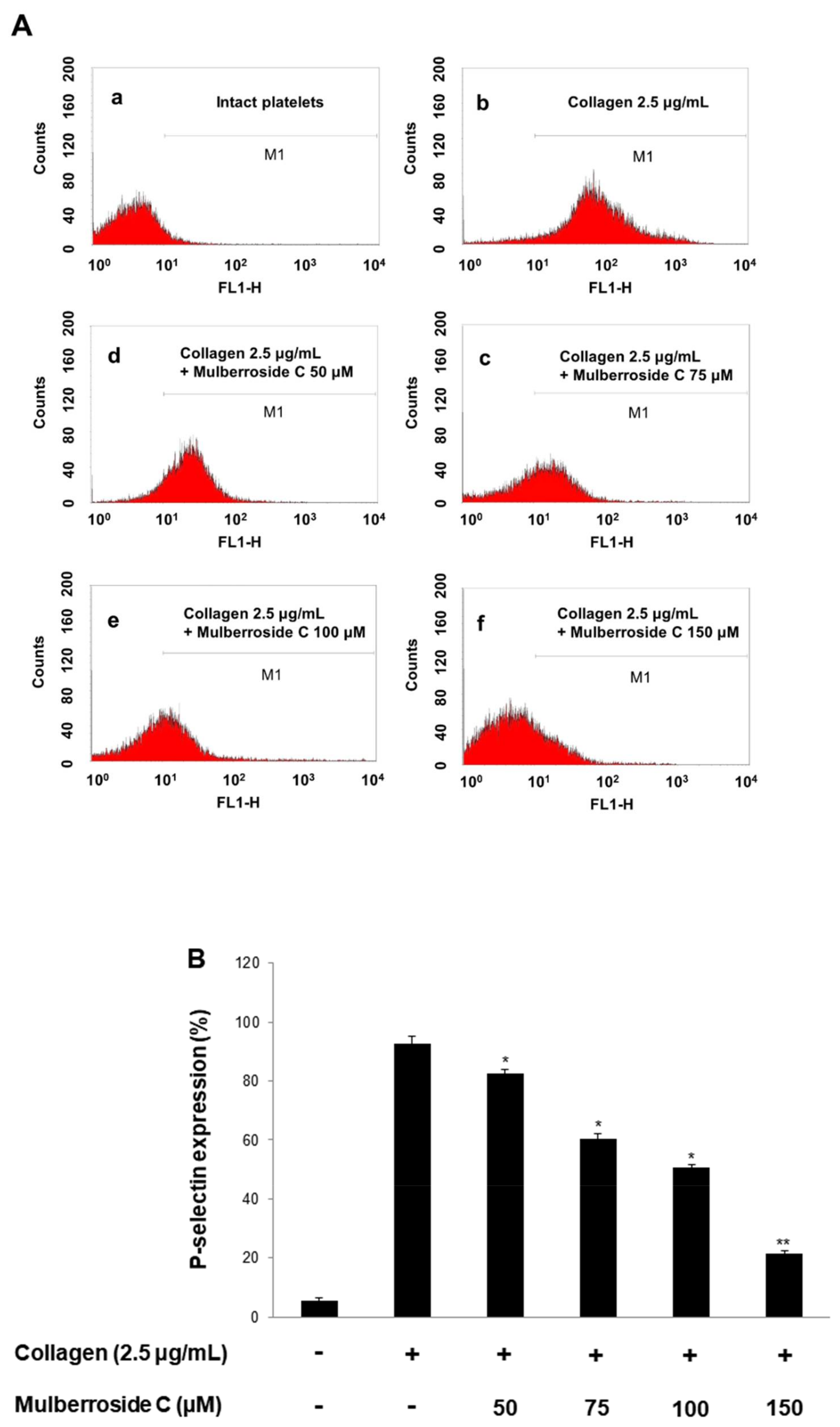
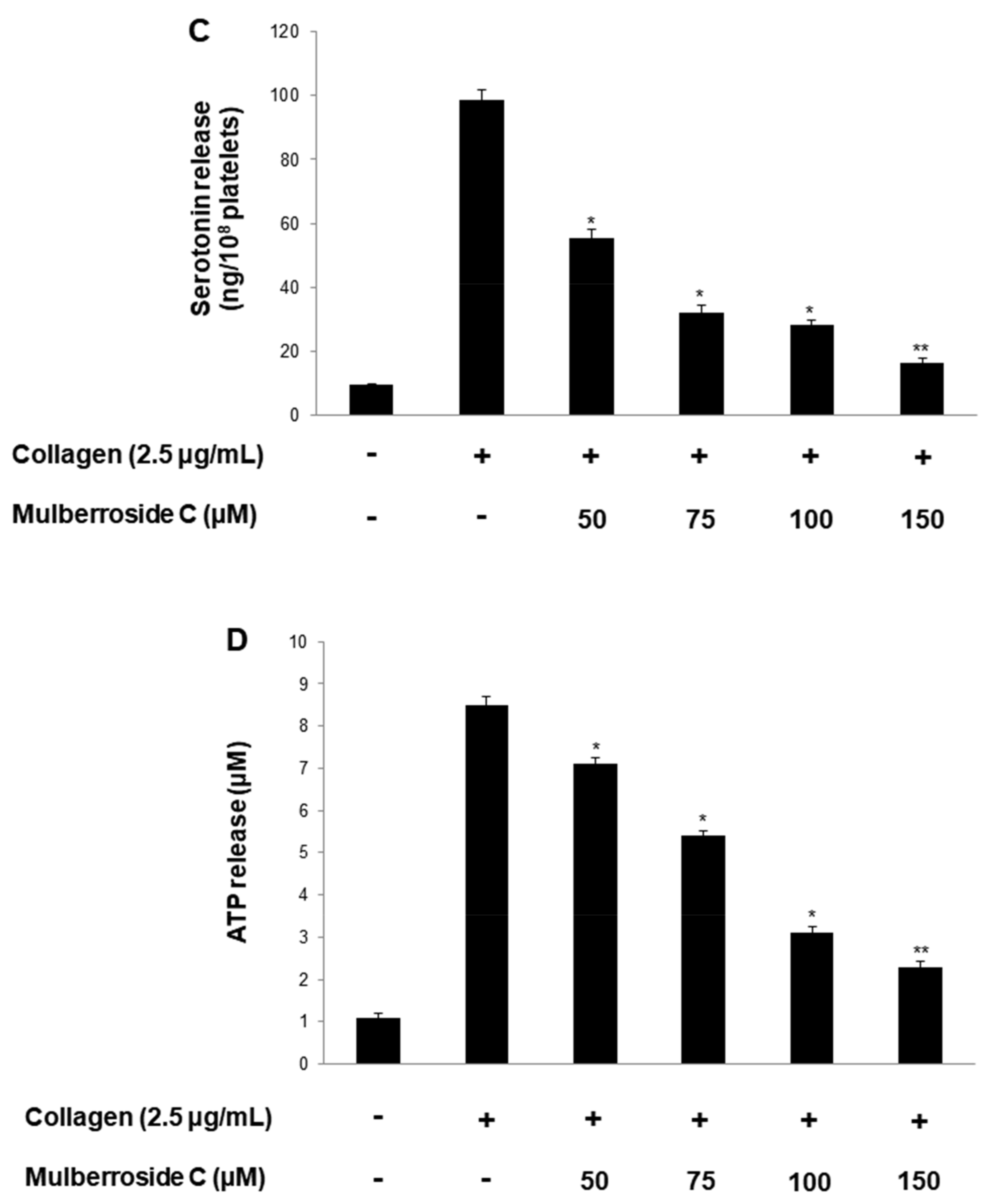
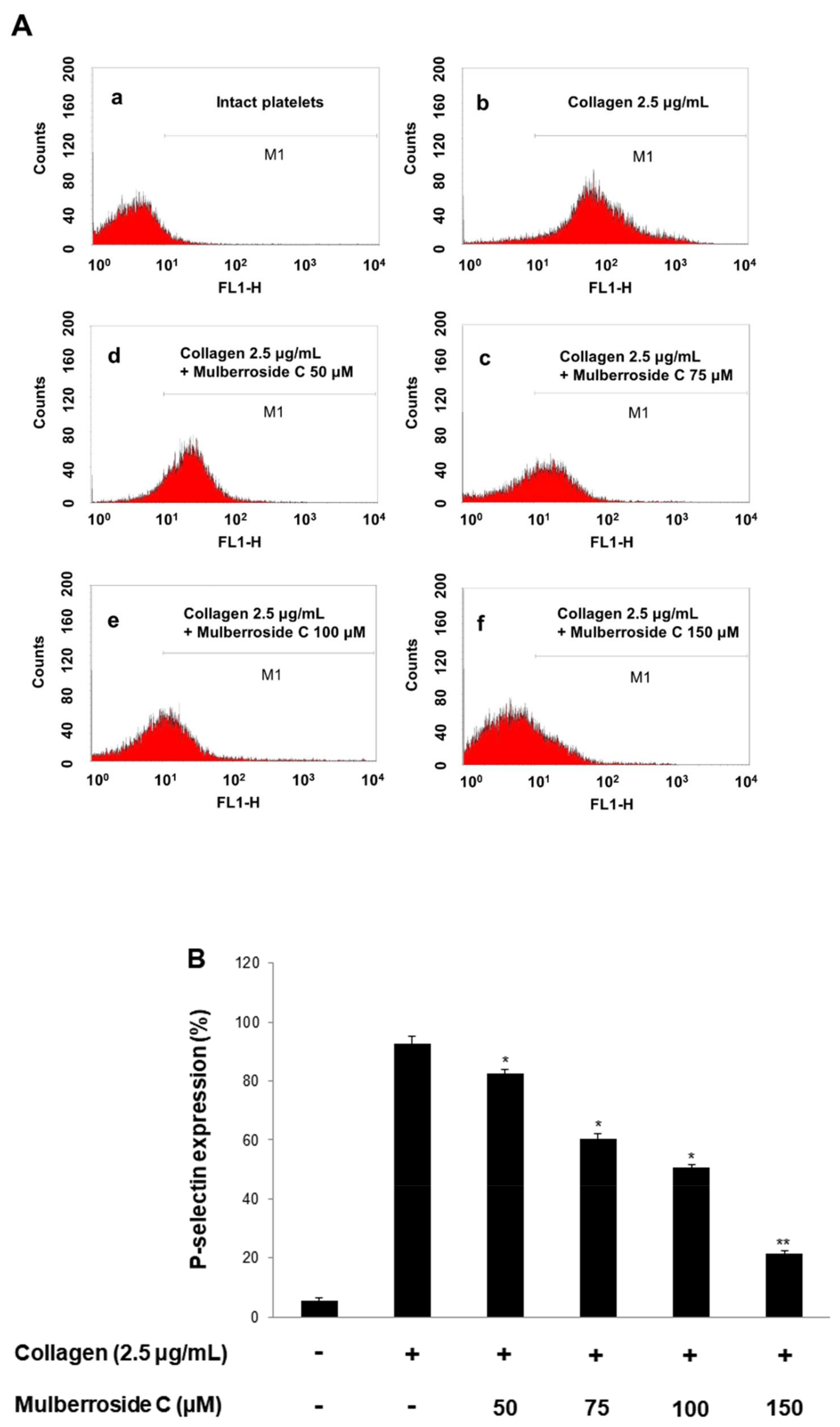
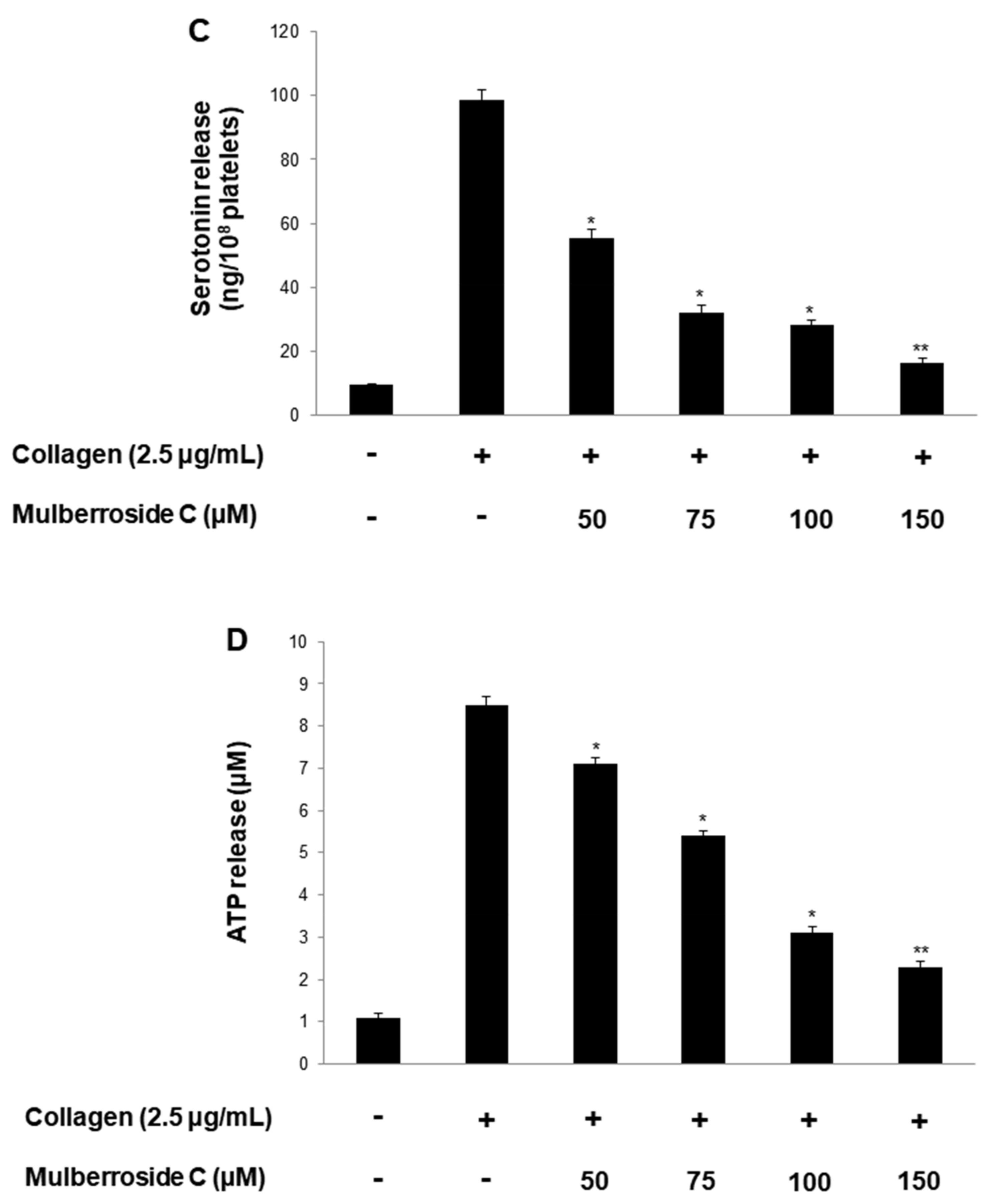
3.3. Mulberroside C Blocks P-Selectin Expression and Serotonin Release
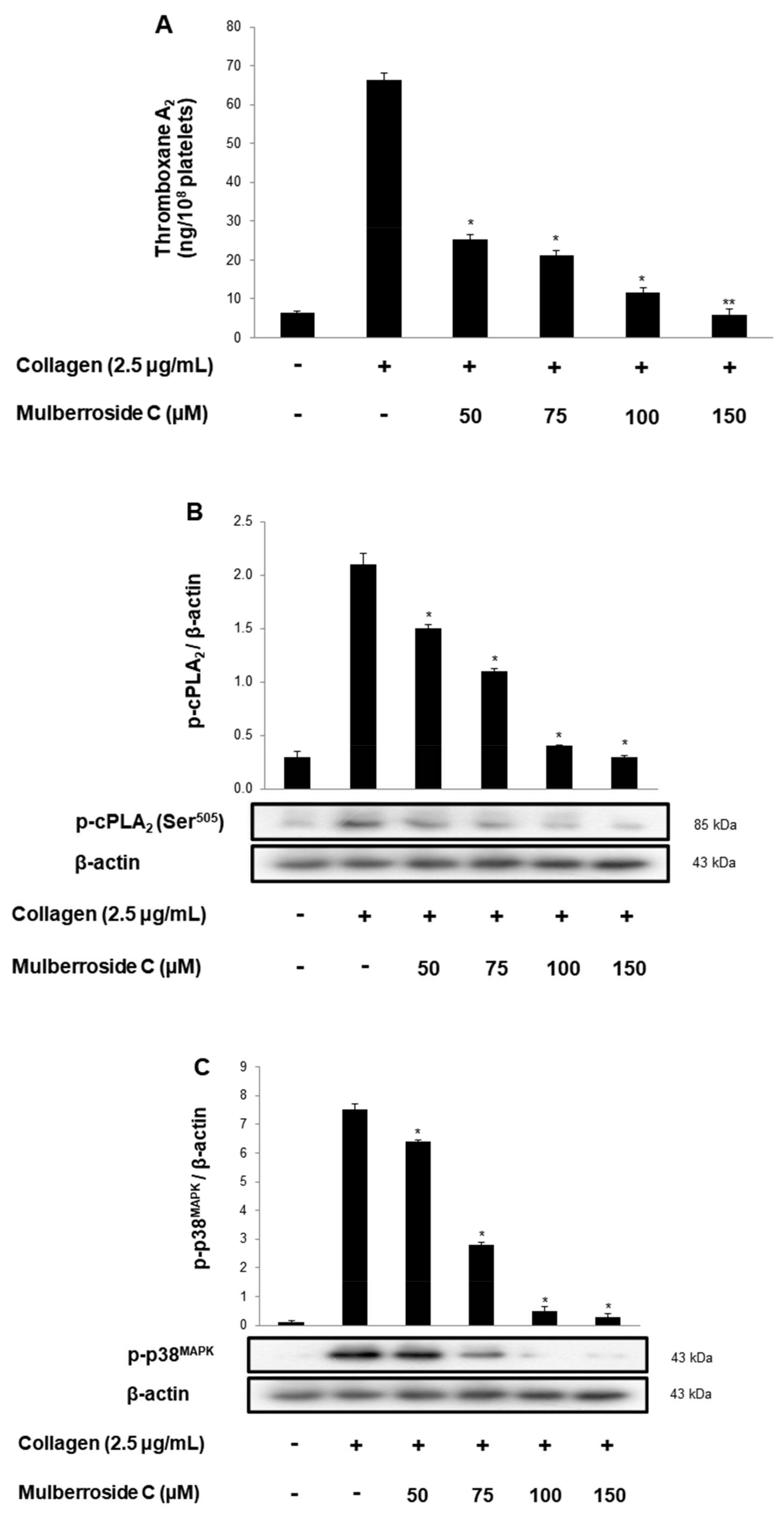
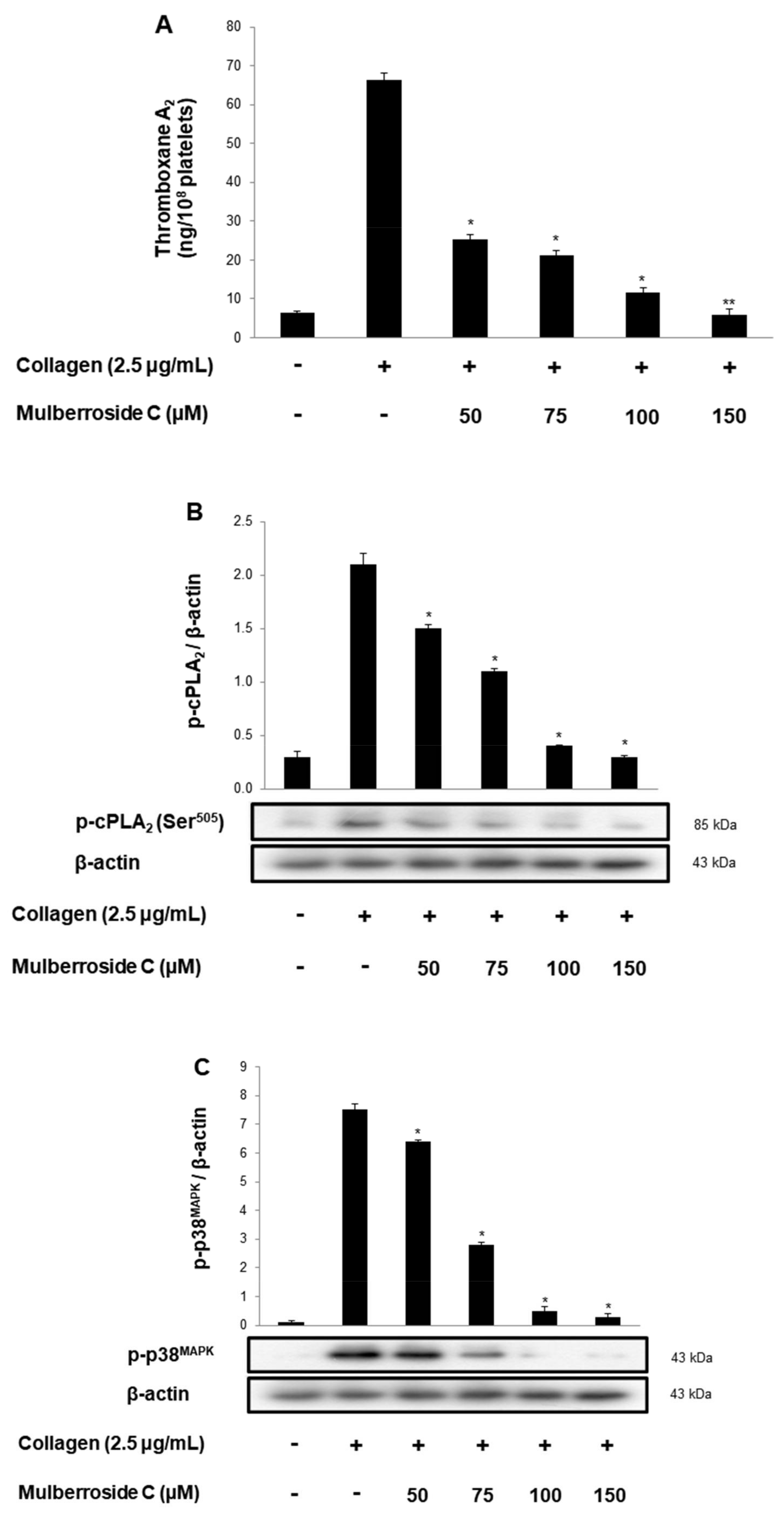
3.4. Mulberroside C Blocks Thromboxane B2 Production, and Dephosphorylation of cPLA2, p38MAPK
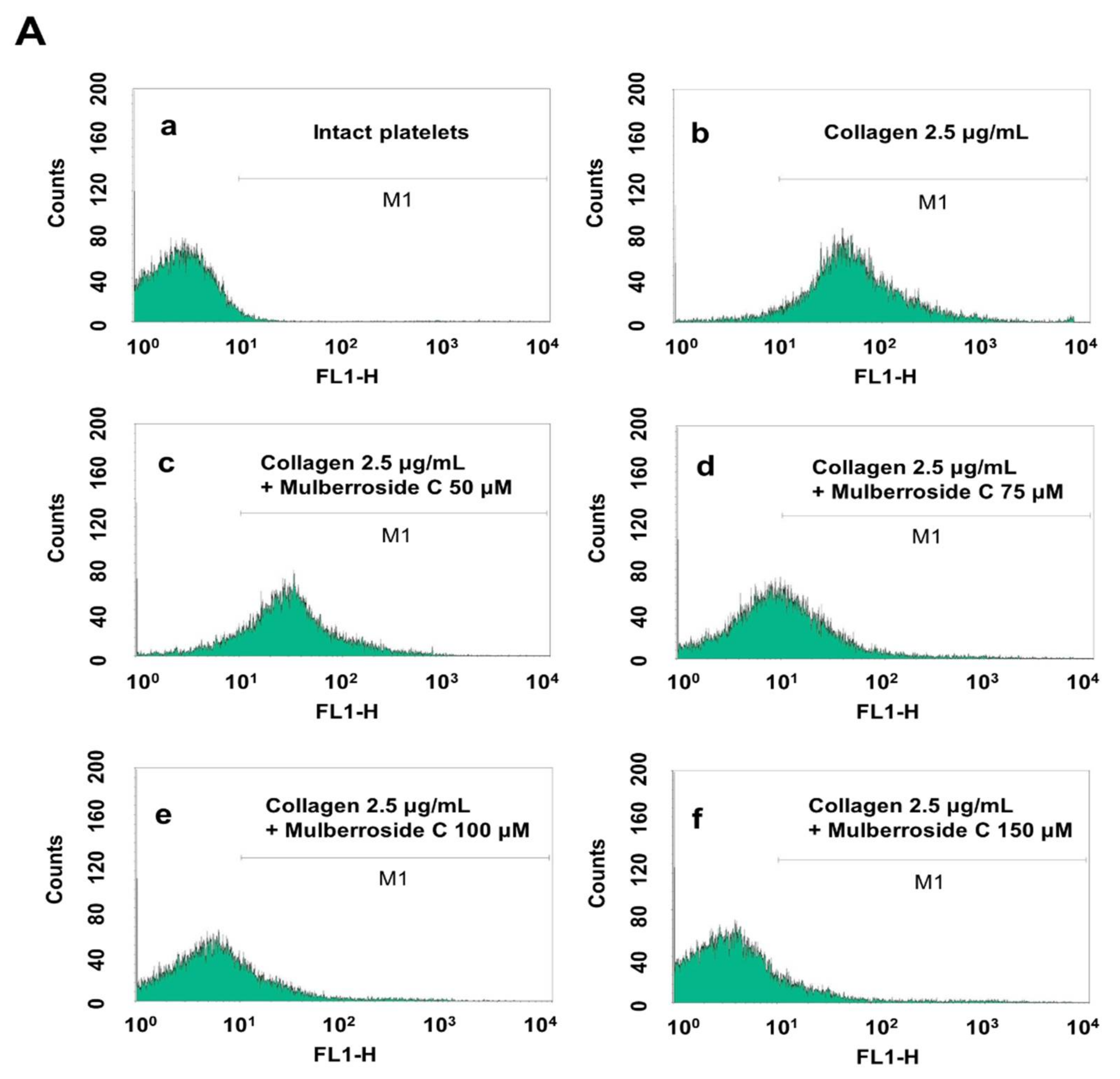
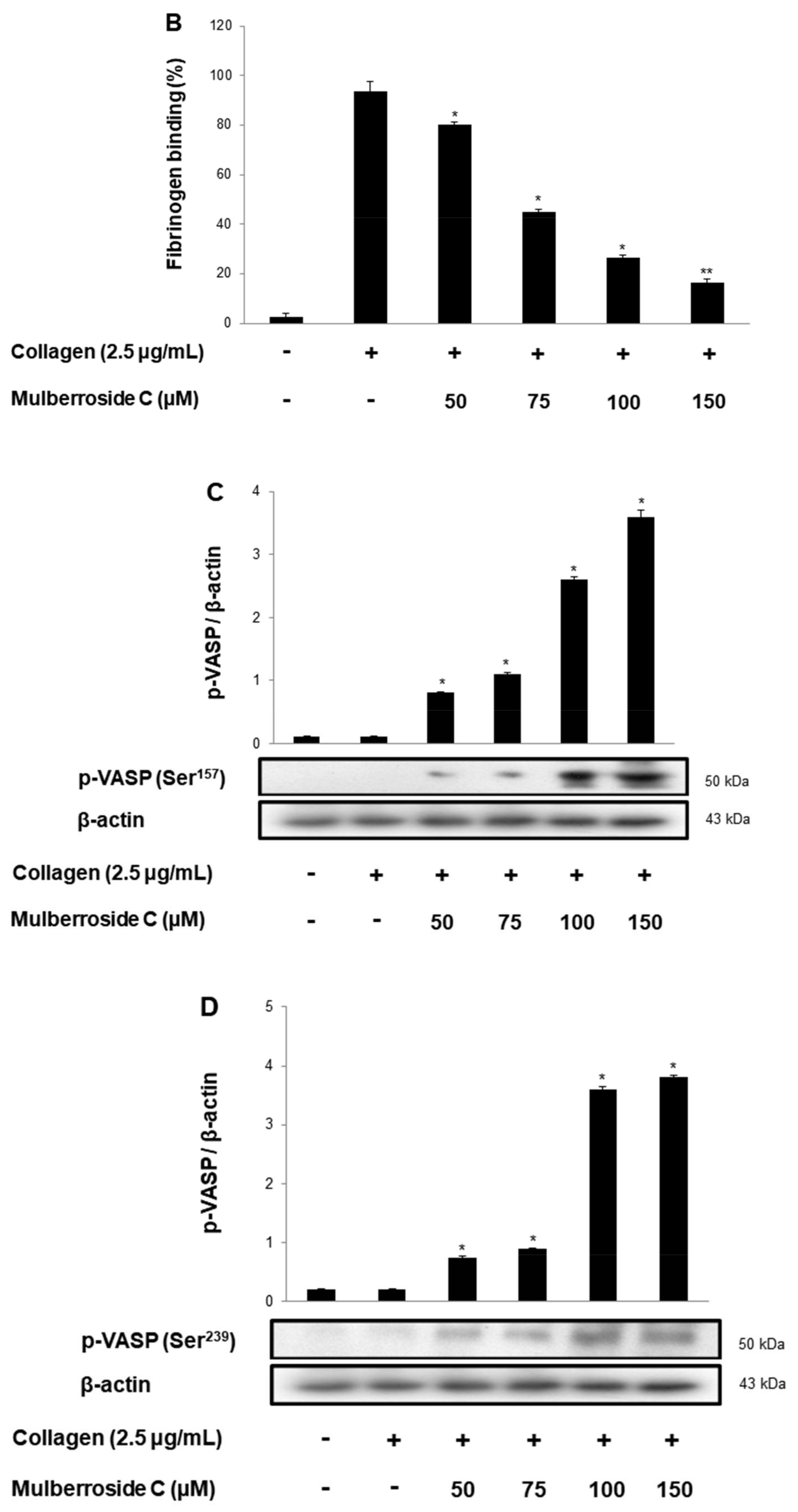
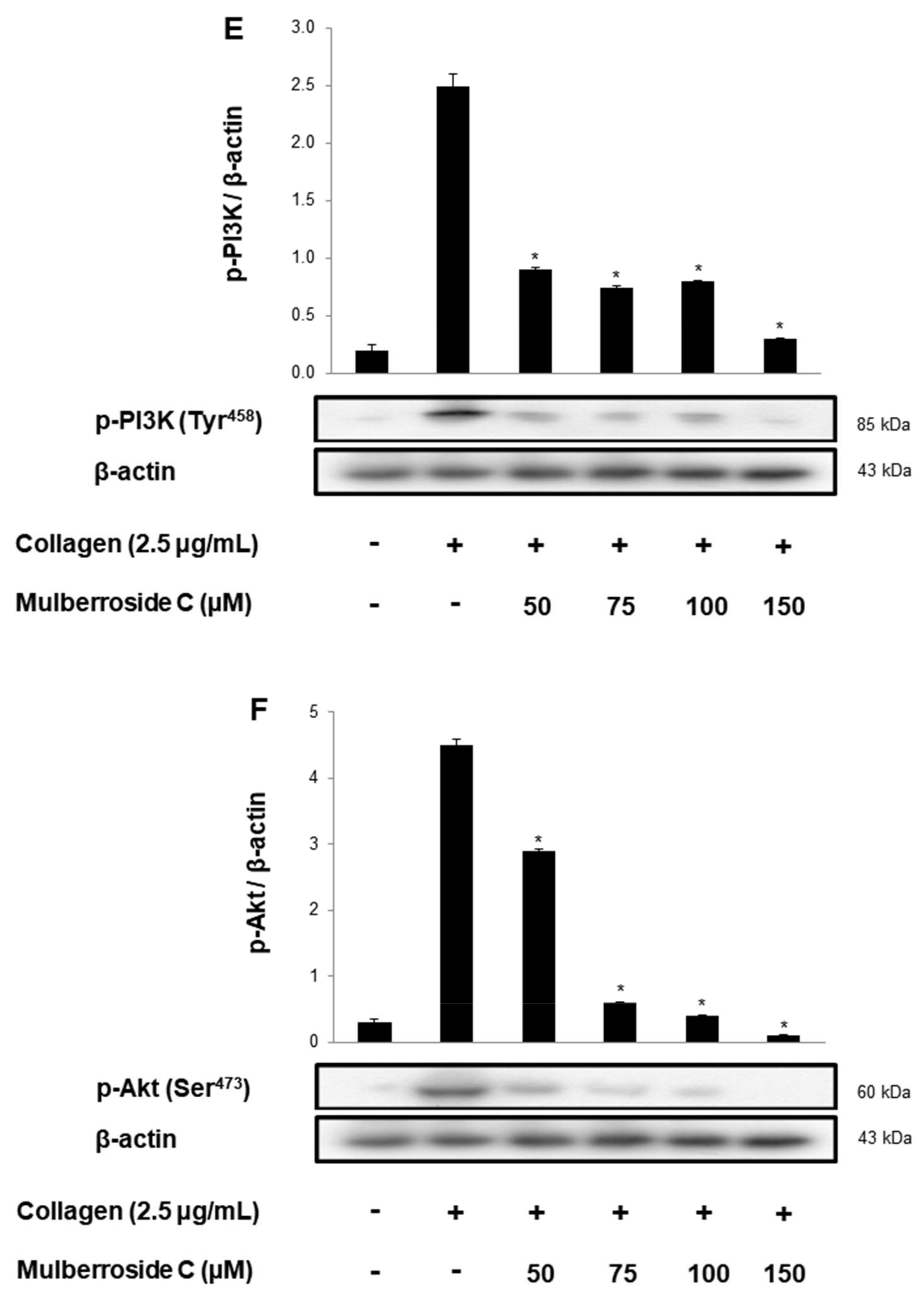
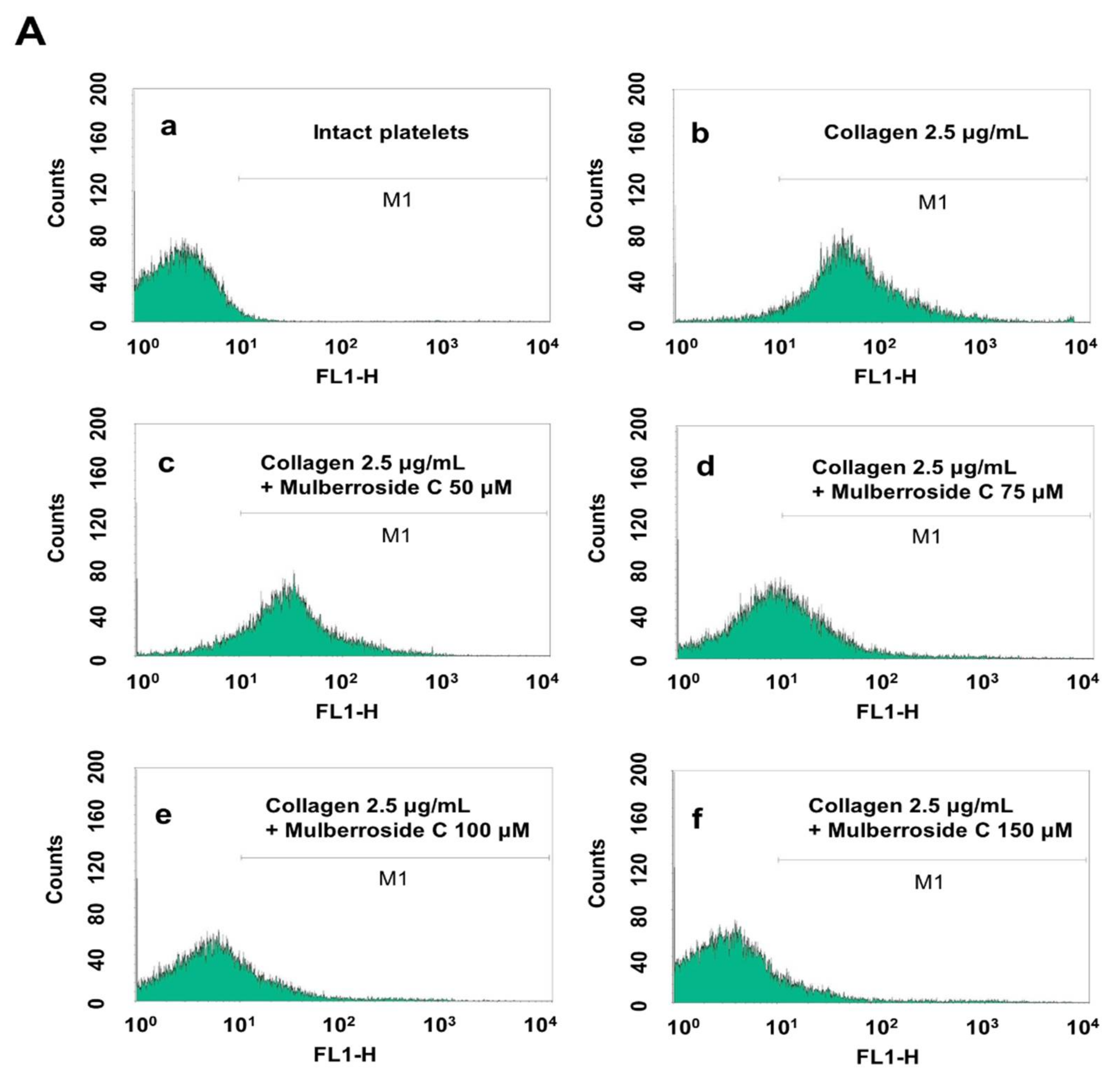
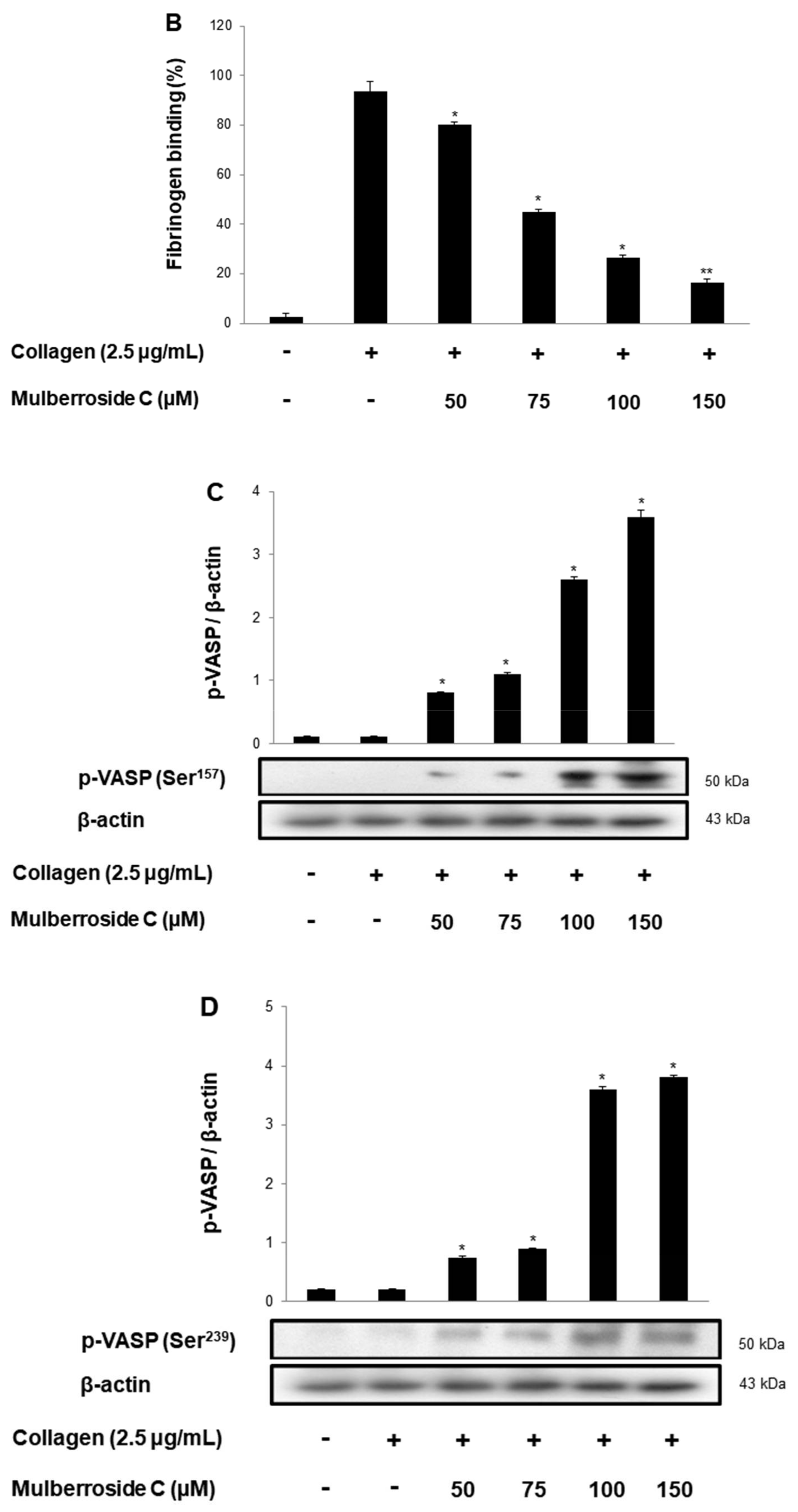
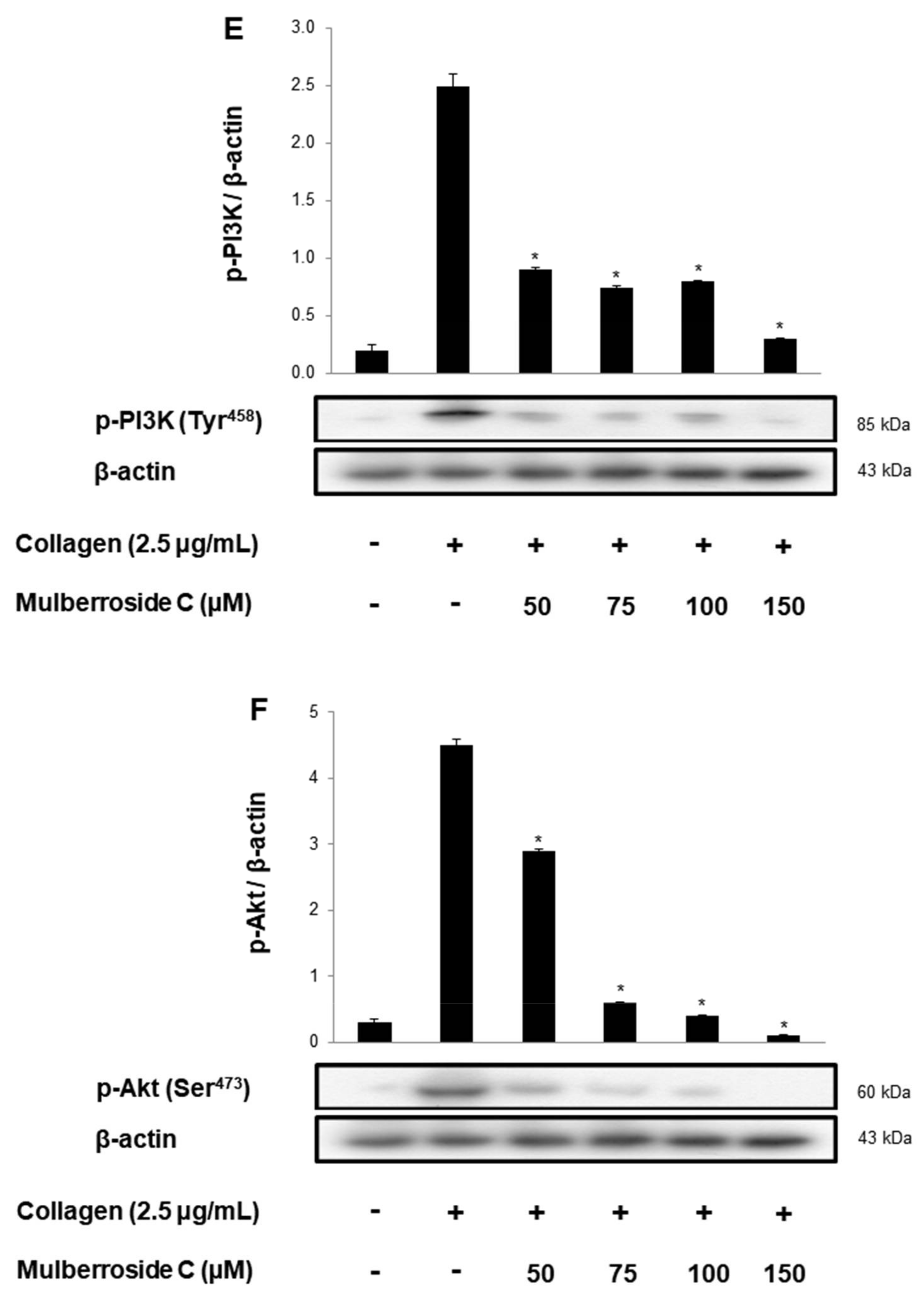
3.5. Mulberroside C Blocks Platelet Binding to Fibrinogen by Limiting αIIb/β3 Affinity and VASP, PI3K/Akt Dephosphoryation
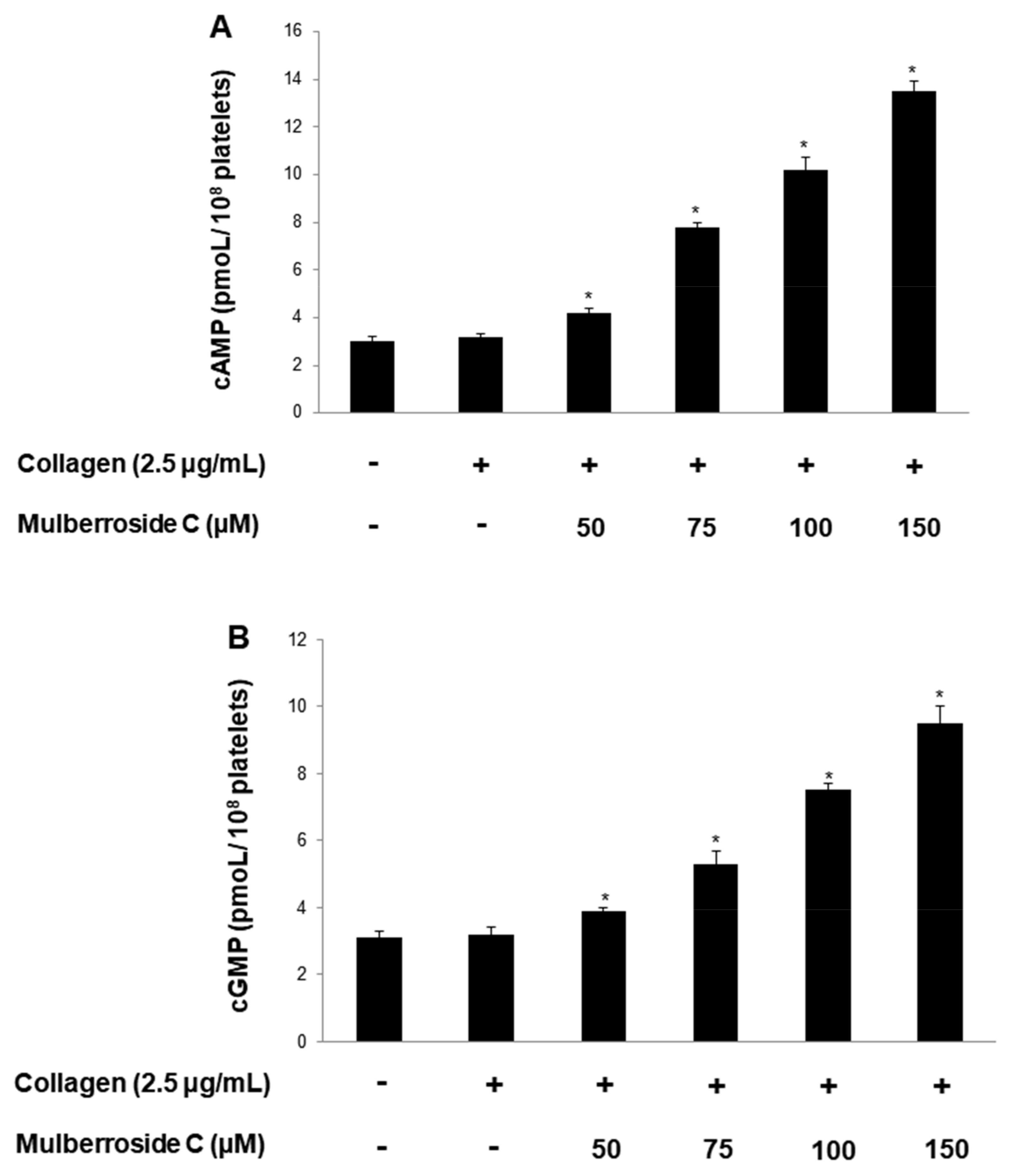
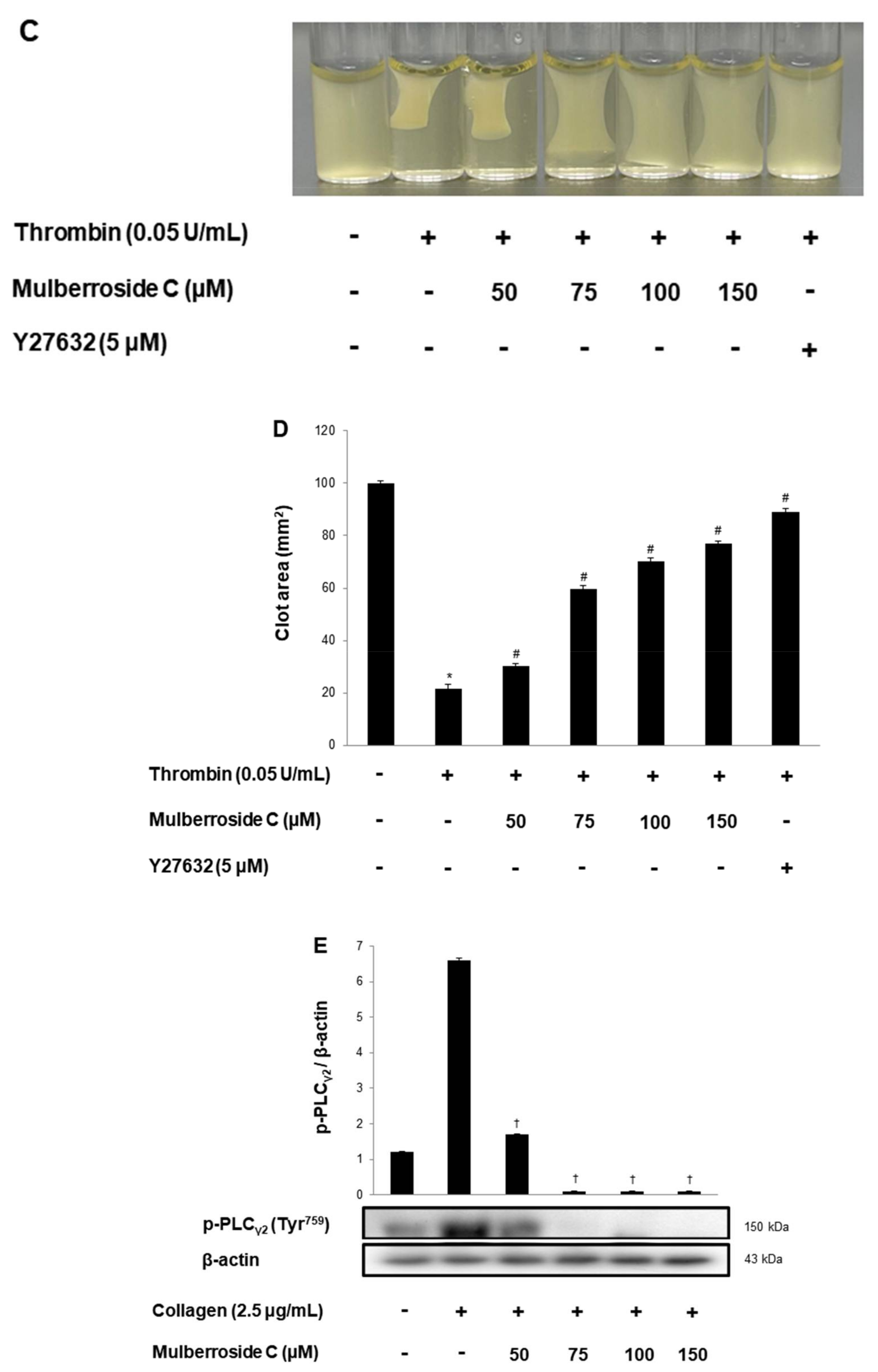
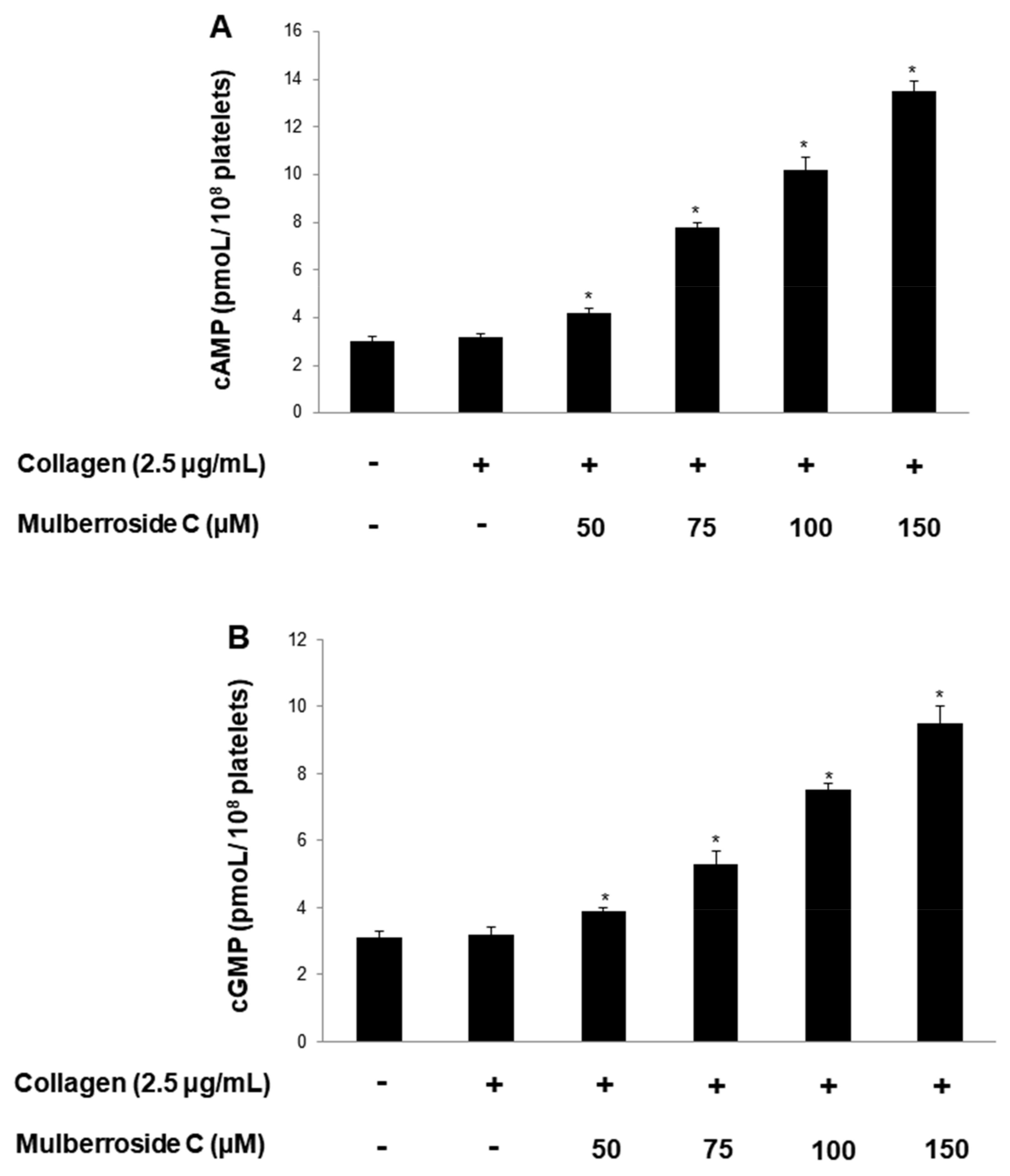
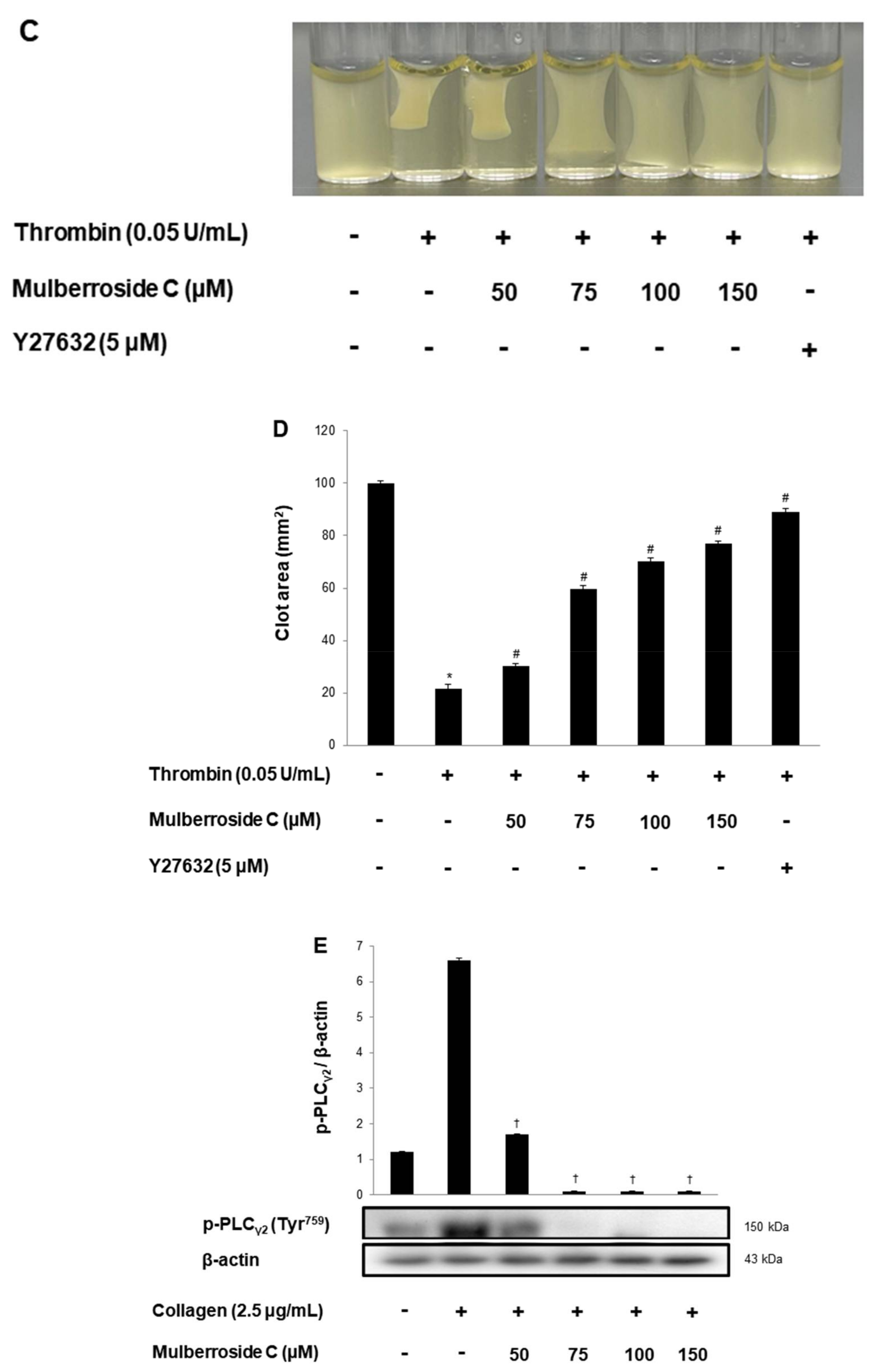
3.6. Mulberroside C Elevated Cyclic Nucleotides and Limits Thrombin-Induced Clot Retraction and Phospholipase Cγ2 Phosphorylation
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; Supuran, C.T. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 1–17. [Google Scholar] [CrossRef]
- Jackson, S.P. Arterial thrombosis-insidious, unpredictable and deadly. Nat. Med. 2011, 17, 1423–1436. [Google Scholar] [CrossRef] [PubMed]
- Chan, E.W.C.; Lye, P.Y.; Wong, S.K. Phytochemistry, pharmacology, and clinical trials of M. Alba. Chin. J. Nat. Med. 2016, 14, 17–30. [Google Scholar] [PubMed]
- Kim, D.S.; Ji, H.D.; Rhee, M.H.; Sung, Y.Y.; Yang, W.K.; Kim, S.H.; Kim, H.K. Antiplatelet activity of M. Alba leaves extract, mediated via inhibiting granule secretion and blocking the phosphorylation of extracellular-signal-regulated kinase and akt. Evid. Based. Complement. Alt. 2014. [Google Scholar] [CrossRef]
- Kim, D.S.; Irfan, M.; Sung, Y.Y.; Kim, S.H.; Park, S.H.; Choi, Y.H.; Kim, H.K. Schisandra chinensis and M. Alba synergistically inhibit in vivo thrombus formation and platelet aggregation by impairing the glycoprotein VI pathway. Evid. Based. Complement. Alt. 2017. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.J.; Yang, H.; Yoo, Y.M.; Hong, S.S.; Lee, D.; Lee, H.J.; Jeung, E.B. Morusinol extracted from M. Alba inhibits arterial thrombosis and modulates platelet activation for the treatment of cardiovascular disease. J. Atheroscler. Thromb. 2012, 6, 1204020479. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; He, Z.D.; Jiang, R.W.; Ye, W.C.; Xu, H.X.; But, P.P.H. Antiviral flavonoids from the root bark of M. Alba L. Phytochemistry 2003, 62, 1235–1238. [Google Scholar] [CrossRef]
- Farndale, R.W. Collagen-induced platelet activation. Blood Cell Mol. Dis. 2006, 36, 162–165. [Google Scholar] [CrossRef]
- Zarbock, A.; Polanowska-Grabowska, R.K.; Ley, K. Platelet-neutrophil-interactions: Linking hemostasis and inflammation. Blood Rev. 2007, 21, 99–111. [Google Scholar] [CrossRef]
- Rosado, J.A.; Sage, S.O. The ERK cascade, a new pathway involved in the activation of store-mediated calcium entry in human platelets. Trends Cardiovasc. Med. 2002, 12, 229–234. [Google Scholar] [CrossRef]
- Kramer, R.M.; Roberts, E.F.; Um, S.L.; Börsch-Haubold, A.G.; Watson, S.P.; Fisher, M.J.; Jakubowski, J.A. p38 mitogen-activated protein kinase phosphorylates cytosolic phospholipase A2 (cPLA2) in thrombin-stimulated platelets Evidence that proline-directed phosphorylation is not required for mobilization of arachidonic acid by cPLA2. J. Biol. Chem. 1996, 271, 27723–27729. [Google Scholar] [CrossRef] [Green Version]
- Needleman, P.; Moncada, S.; Bunting, S.; Vane, J.R.; Hamberg, M.; Samuelsson, B. Identification of an enzyme in platelet microsomes which generates thromboxaneA2 from prostaglandin endoperoxides. Nature 1976, 261, 558–560. [Google Scholar] [CrossRef]
- Smyth, E.M. Thromboxane and the thromboxane receptor in cardiovascular disease. J. Clin. Lipidol. 2010, 5, 209–219. [Google Scholar] [CrossRef] [Green Version]
- Smolenski, A. Novel roles of cAMP/cGMP-dependent signaling in platelets. J. Thromb. Haemost. 2012, 10, 167–176. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, U.R.; Walter, U.; Eigenthaler, M. Taming platelets with cyclic nucleotides. Biochem. Pharmacol. 2001, 62, 1153–1161. [Google Scholar] [CrossRef]
- Shin, J.H.; Kwon, H.W.; Lee, D.H. Ginsenoside F4 inhibits platelet aggregation and thrombus formation by dephosphorylation of IP3RI and VASP. J. Appl. Biol. Chem. 2019, 62, 93–100. [Google Scholar] [CrossRef] [Green Version]
- Grynkiewicz, G.; Poenie, M.; Tsien, R.Y. A new generation of Ca2+ indicators with greatly improved fluorescence properties. J. Biol. Chem. 1985, 260, 3440–3450. [Google Scholar] [CrossRef]
- Laurent, V.; Loisel, T.P.; Harbeck, B.; Wehman, A.; Gröbe, L.; Jockusch, B.M.; Wehland, J.; Gertler, F.B.; Carlier, M.F. Role of proteins of the Ena/VASP family in actin-based motility of Listeria monocytogenes. J. Cell Biol. 1999, 144, 1245–1258. [Google Scholar] [CrossRef] [Green Version]
- Sudo, T.; Ito, H.; Kimura, Y. Phosphorylation of the vasodilator-stimulated phosphoprotein (VASP) by the anti-platelet drug, cilostazol, in platelets. Platelets 2003, 4, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; De, S.; Damron, D.S.; Chen, W.S.; Hay, N.; Byzova, T.V. Impaired platelet responses to thrombin and collagen in AKT-1–deficient mice. Blood 2004, 104, 1703–1710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valet, C.; Severin, S.; Chicanne, G.; Laurent, P.A.; Gaits-Iacovoni, F.; Gratacap, M.P.; Payrastre, B. The role of class I, II and III PI 3-kinases in platelet production and activation and their implication in thrombosis. Adv. Biol Regul. 2016, 61, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Suzuki-Inoue, K.; Hughes, C.E.; Inoue, O.; Kaneko, M.; Cuyun-Lir, O.; Takafuta, T.; Watson, S.P.; Ozaki, Y. Involvement of Src kinases and PLCγ2 in clot retraction. Thromb. Res. 2007, 120, 251–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madhumitha, S.; Indhuleka, A. Cardioprotective effect of M. Alba L. leaves in isoprenaline induced rats. Int. J. Pharm. Sci. Res. 2012, 3, 1475. [Google Scholar]
- Panth, N.; Paudel, K.R.; Gong, D.S.; Oak, M.H. Vascular protection by ethanol extract of M. Alba root bark: Endothelium-dependent relaxation of rat aorta and decrease of smooth muscle cell migration and proliferation. Evid. Based. Complement. Alt. 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Payrastre, B.; Missy, K.; Trumel, C.; Bodin, S.; Plantavid, M.; Chap, H. The integrin αIIb/β3 in human platelet signal transduction. Biochem. Pharmacol. 2000, 60, 1069–1074. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwon, H.-W.; Lee, D.-H.; Rhee, M.H.; Shin, J.-H. In Vitro Antiplatelet Activity of Mulberroside C through the Up-Regulation of Cyclic Nucleotide Signaling Pathways and Down-Regulation of Phosphoproteins. Genes 2021, 12, 1024. https://doi.org/10.3390/genes12071024
Kwon H-W, Lee D-H, Rhee MH, Shin J-H. In Vitro Antiplatelet Activity of Mulberroside C through the Up-Regulation of Cyclic Nucleotide Signaling Pathways and Down-Regulation of Phosphoproteins. Genes. 2021; 12(7):1024. https://doi.org/10.3390/genes12071024
Chicago/Turabian StyleKwon, Hyuk-Woo, Dong-Ha Lee, Man Hee Rhee, and Jung-Hae Shin. 2021. "In Vitro Antiplatelet Activity of Mulberroside C through the Up-Regulation of Cyclic Nucleotide Signaling Pathways and Down-Regulation of Phosphoproteins" Genes 12, no. 7: 1024. https://doi.org/10.3390/genes12071024
APA StyleKwon, H.-W., Lee, D.-H., Rhee, M. H., & Shin, J.-H. (2021). In Vitro Antiplatelet Activity of Mulberroside C through the Up-Regulation of Cyclic Nucleotide Signaling Pathways and Down-Regulation of Phosphoproteins. Genes, 12(7), 1024. https://doi.org/10.3390/genes12071024