Characterization of QTLs and Candidate Genes for Days to Heading in Rice Recombinant Inbred Lines

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Phenotype Evaluation
2.2. Sequencing Library Construction and Genotyping
2.3. Linkage Mapping and Statistical Analysis
2.4. Sequencing and Marker Analysis for Hd16, Hd1, and Ghd7
3. Results
3.1. Days to Heading and Grain Filling Rate of Baegilmi in Comparison with Koshihikari
3.2. Mapping of qDH3, qDH6, and qDH7 for Days to Heading
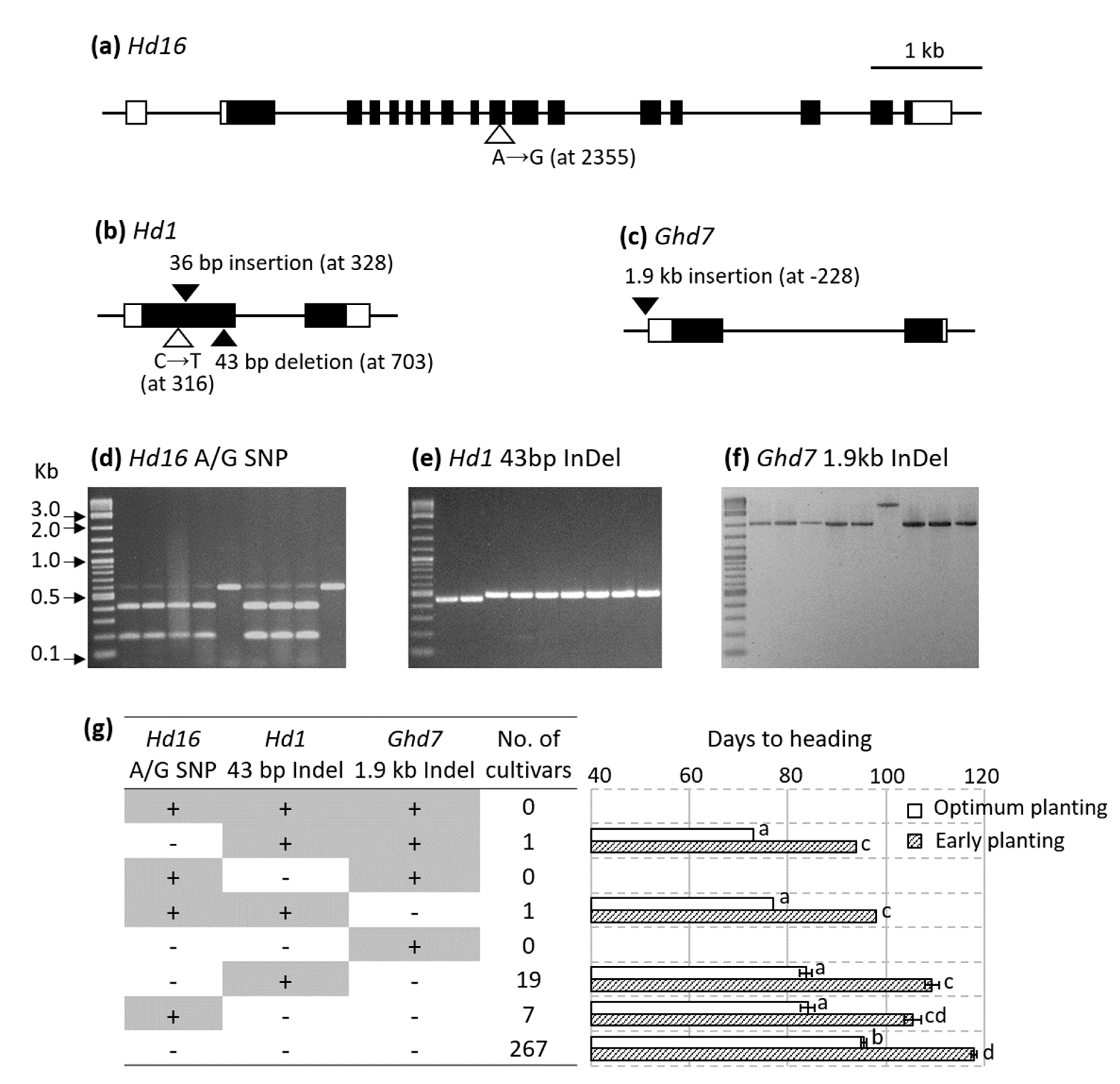
3.3. Hd16, Hd1, and Ghd7 Underlying the Days to Heading QTLs
3.4. Allelic Composition of Hd16, Hd1, and Ghd7 among Commercial Rice Cultivars
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shrestha, R.; Gómez-Ariza, J.; Brambilla, V.; Fornara, F. Molecular control of seasonal flowering in rice, arabidopsis and temperate cereals. Ann. Bot. 2014, 114, 1445–1458. [Google Scholar] [CrossRef] [PubMed]
- Hori, K.; Matsubara, K.; Yano, M. Genetic control of flowering time in rice: Integration of Mendelian genetics and genomics. Theor. Appl. Genet. 2016, 129, 2241–2252. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, K.; Yano, M. Genetic and molecular dissection of flowering time control in rice. In Rice Genomics, Genetics and Breeding; Sasaki, T., Ashikari, M., Eds.; Springer: Singapore, 2018; pp. 177–190. [Google Scholar]
- Blümel, M.; Dally, N.; Jung, C. Flowering time regulation in crops—What did we learn from Arabidopsis? Curr. Opin. Biotech. 2015, 32, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Komiya, R.; Yokoi, S.; Shimamoto, K. A gene network for long-day flowering activates RFT1 encoding a mobile flowering signal in rice. Development 2009, 136, 3443–3450. [Google Scholar] [CrossRef] [PubMed]
- Tamaki, S.; Matsuo, S.; Hann, L.W.; Yokoi, S.; Shimamoto, K. Hd3a protein is a mobile flowering signal in rice. Science 2007, 316, 1033–1036. [Google Scholar] [CrossRef] [PubMed]
- Yano, M.; Katayose, Y.; Ashikari, M.; Yamanouchi, U.; Monna, L.; Fuse, T.; Baba, T.; Yamamoto, K.; Umehara, Y.; Nagamura, Y.; et al. Hd1, a major photoperiod sensitivity quantitative trait locus in rice, is closely related to the Arabidopsis flowering time gene CONSTANS. Plant Cell 2000, 12, 2473–2483. [Google Scholar] [CrossRef] [PubMed]
- Hayama, R.; Yokoi, S.; Tamaki, S.; Yano, M.; Shimamoto, K. Adaptation of photoperiodic control pathways produces short-day flowering in rice. Nature 2003, 422, 719–722. [Google Scholar] [CrossRef]
- Doi, K.; Izawa, T.; Fuse, T.; Yamanouchi, U.; Kubo, T.; Shimatani, Z.; Yano, M.; Yoshimura, A. Ehd1, a B-type response regulator in rice, confers short-day promotion of flowering and controls FT-like gene expression independently of Hd1. Genes Dev. 2004, 18, 926–936. [Google Scholar] [CrossRef]
- Xue, W.; Xing, Y.; Weng, X.; Zhao, Y.; Tang, W.; Wang, L.; Zhou, H.; Yu, S.; Xu, C.; Li, X.; et al. Natural variation in Ghd7 is an important regulator of heading date and yield potential in rice. Nat. Genet. 2008, 40, 761–767. [Google Scholar] [CrossRef]
- Kim, S.Y.; Seo, J.H.; Bae, H.K.; Hwang, C.D.; Ko, J.M. Rice cultivars adaptable for rice based cropping systems in a paddy field in the Yeongnam plain area of Korea. Korean J. Agri. Sci. 2018, 45, 355–363. [Google Scholar] [CrossRef]
- Kim, Y.D.; Kang, S.G.; Ku, B.I.; Choi, M.K.; Park, H.K.; Park, T.S.; Back, N.H.; Kim, S.J.; Ko, J.K. Selection of suitable rice cultivars for silage barley-rice double cropping in Honam plain area. Korean J. Int. Agric. 2010, 22, 337–340. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Jeung, J.; Cho, Y.; Kim, B. Development of early maturing rice lines with genes conferring resistance to bacterial blight and rice stripe virus for enhancing the adaptability in plain area. Korean J. Breed. Sci 2015, 47, 118–127. [Google Scholar] [CrossRef]
- Mo, Y.; Jeong, J.-M.; Kim, W.-J.; Kim, B.-K.; Jeung, J.-U. ‘Baegilmi’, an extremely early maturing blast resistant rice with good grain appearance. Korean J. Breed. Sci. 2019, 51, 151–159. [Google Scholar] [CrossRef][Green Version]
- Kobayashi, A.; Hori, K.; Yamamoto, T.; Yano, M. Koshihikari: A premium short-grain rice cultivar—Its expansion and breeding in Japan. Rice 2018, 11, 15. [Google Scholar] [CrossRef] [PubMed]
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4326. [Google Scholar] [CrossRef]
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A robust, simple genotyping-by-sequencing (GBS) approach for high diversity species. PLoS ONE 2011, 6, e19379. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate long-read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef]
- Glaubitz, J.C.; Casstevens, T.M.; Lu, F.; Harriman, J.; Elshire, R.J.; Sun, Q.; Buckler, E.S. TASSEL-GBS: A high capacity genotyping by sequencing analysis pipeline. PLoS ONE 2014, 9, e90346. [Google Scholar] [CrossRef]
- Jang, Y.J.; Seo, M.; Hersh, C.P.; Rhee, S.J.; Kim, Y.; Lee, G.P. An evolutionarily conserved non-synonymous SNP in a leucine-rich repeat domain determines anthracnose resistance in watermelon. Theor. Appl. Genet. 2019, 132, 473–488. [Google Scholar] [CrossRef]
- Meng, L.; Li, H.; Zhang, L.; Wang, J. QTL IciMapping: Integrated software for genetic linkage map construction and quantitative trait locus mapping in biparental populations. Crop. J. 2015, 3, 269–283. [Google Scholar] [CrossRef]
- Kim, S.R.; Torollo, G.; Yoon, M.R.; Kwak, J.; Lee, C.K.; Prahalada, G.D.; Choi, I.R.; Yeo, U.S.; Jeong, O.Y.; Jena, K.K.; et al. Loss-of-function alleles of Heading date 1 (Hd1) are associated with adaptation of temperate japonica rice plants to the tropical region. Front. Plant Sci. 2018, 9, 1827. [Google Scholar] [CrossRef]
- Hori, K.; Ogiso-Tanaka, E.; Matsubara, K.; Yamanouchi, U.; Ebana, K.; Yano, M. Hd16, a gene for casein kinase I, is involved in the control of rice flowering time by modulating the day-length response. Plant J. 2013, 76, 36–46. [Google Scholar] [CrossRef]
- Fujino, K.; Yamanouchi, U. Genetic effect of a new allele for the flowering time locus Ghd7 in rice. Breed. Sci. 2020, 70, 342–346. [Google Scholar] [CrossRef]
- Kojima, S.; Takahashi, Y.; Kobayashi, Y.; Monna, L.; Sasaki, T.; Araki, T.; Yano, M. Hd3a, a rice ortholog of the Arabidopsis FT gene, promotes transition to flowering downstream of Hd1 under short-day conditions. Plant Cell Physiol. 2002, 43, 1096–1105. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y.; Teshima, K.M.; Yokoi, S.; Innan, H.; Shimamoto, K. Variations in Hd1 proteins, Hd3a promoters, and Ehd1 expression levels contribute to diversity of flowering time in cultivated rice. Proc. Natl. Acad. Sci. USA 2009, 106, 4555–4560. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Ariza, J.; Galbiati, F.; Goretti, D.; Brambilla, V.; Shrestha, R.; Pappolla, A.; Courtois, B.; Fornara, F. Loss of floral repressor function adapts rice to higher latitudes in Europe. J. Exp. Bot. 2015, 66, 2027–2039. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhou, X.; Yan, W.; Zhang, Z.; Lu, L.; Han, Z.; Zhao, H.; Liu, H.; Song, P.; Hu, Y.; et al. Combinations of the Ghd7, Ghd8 and Hd1 genes largely define the ecogeographical adaptation and yield potential of cultivated rice. New Phytol. 2015, 208, 1056–1066. [Google Scholar] [CrossRef] [PubMed]
- Saito, H.; Okumoto, Y.; Tsukiyama, T.; Xu, C.; Teraishi, M.; Tanisaka, T. Allelic differentiation at the E1/Ghd7 locus has allowed expansion of rice cultivation area. Plants 2019, 8, 550. [Google Scholar] [CrossRef]
- Wei, F.J.; Tsai, Y.C.; Wu, H.P.; Huang, L.T.; Chen, Y.C.; Chen, Y.F.; Wu, C.C.; Tseng, Y.T.; Hsing, Y.C. Both Hd1 and Ehd1 are important for artificial selection of flowering time in cultivated rice. Plant Sci. 2015, 242, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.M.; Park, H.S.; Baek, M.K.; Suh, J.P.; Kim, C.S.; Lee, K.M.; Park, S.G.; Cho, Y.C. Characterization of grain-related traits of 300 Korean rice varieties. In Proceedings of the 2019 Korean Society of Breeding Science (KSBS) & Society for the Advancement of Breeding Research in Asia and Oceania (SABRAO), Gwangju, Korea, 2–5 July 2019. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| QTL | Chr a | Flanking Markers b | Year | Peak (cM) | LOD c | PVE d (%) | Add e | |
|---|---|---|---|---|---|---|---|---|
| Left | Right | |||||||
| qDH3 | 3 | S3_28142709 | S3_34851991 | 2016 | 129 | 7.5 | 10.3 | −3.4 |
| 2017 | 129 | 5.1 | 8.4 | −3.1 | ||||
| qDH6 | 6 | S6_8634012 | S6_10449013 | 2016 | 39 | 15.1 | 24.7 | 5.2 |
| 2017 | 40 | 12.6 | 26.2 | 5.4 | ||||
| qDH7 | 7 | S7_5256691 | S7_10453336 | 2016 | 45 | 13.7 | 22.8 | 5.0 |
| 2017 | 46 | 12.7 | 25.2 | 5.3 | ||||
| Year | Three-Way ANOVA | Mean DH c | |||||
|---|---|---|---|---|---|---|---|
| Effect a | F Value | p Value | PVE b | K | B | ||
| 2016 | Main | qDH3 | 24.5 | 2.5 × 10−6 | 7.6 | 76.4 | 81.6 |
| qDH6 | 63.7 | 1.2 × 10−12 | 19.8 | 83.4 | 73.9 | ||
| qDH7 | 82.3 | 3.8 × 10−15 | 25.5 | 84.3 | 73.6 | ||
| Interaction | qDH3 × qDH6 | 4.1 | 0.0453 | 1.3 | |||
| qDH6 × qDH7 | 0.5 | ns | - | ||||
| qDH3 × qDH7 | 4.8 | 0.0301 | 1.5 | ||||
| qDH3 × qDH6 × qDH7 | 5.5 | 0.0206 | 1.7 | ||||
| 2017 | Main | qDH3 | 19.4 | 2.6 × 10−5 | 6.6 | 79.6 | 84.7 |
| qDH6 | 54.9 | 3.3 × 10−11 | 18.8 | 86.9 | 76.9 | ||
| qDH7 | 73.4 | 9.7 × 10−14 | 25.2 | 87.7 | 76.3 | ||
| Interaction | qDH3 × qDH6 | 2.0 | ns | - | |||
| qDH6 × qDH7 | 2.8 | ns | - | ||||
| qDH3 × qDH7 | 4.2 | 0.0433 | 1.4 | ||||
| qDH3 × qDH6 × qDH7 | 3.1 | ns | - | ||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mo, Y.; Jeong, J.-M.; Ha, S.-K.; Kim, J.; Lee, C.; Lee, G.P.; Jeung, J.-U. Characterization of QTLs and Candidate Genes for Days to Heading in Rice Recombinant Inbred Lines. Genes 2020, 11, 957. https://doi.org/10.3390/genes11090957
Mo Y, Jeong J-M, Ha S-K, Kim J, Lee C, Lee GP, Jeung J-U. Characterization of QTLs and Candidate Genes for Days to Heading in Rice Recombinant Inbred Lines. Genes. 2020; 11(9):957. https://doi.org/10.3390/genes11090957
Chicago/Turabian StyleMo, Youngjun, Jong-Min Jeong, Su-Kyung Ha, Jinhee Kim, Changmin Lee, Gung Pyo Lee, and Ji-Ung Jeung. 2020. "Characterization of QTLs and Candidate Genes for Days to Heading in Rice Recombinant Inbred Lines" Genes 11, no. 9: 957. https://doi.org/10.3390/genes11090957
APA StyleMo, Y., Jeong, J.-M., Ha, S.-K., Kim, J., Lee, C., Lee, G. P., & Jeung, J.-U. (2020). Characterization of QTLs and Candidate Genes for Days to Heading in Rice Recombinant Inbred Lines. Genes, 11(9), 957. https://doi.org/10.3390/genes11090957