A SNP-Based Genome-Wide Association Study to Mine Genetic Loci Associated to Salinity Tolerance in Mungbean (Vigna radiata L.)


Abstract
1. Introduction
- (1)
- Tolerance to osmotic stress—under osmotic stress (caused through salt or drought), changes in the turgor of leaves cause stomata closure, reducing transpiration and ensuring water retention. Loss in turgidity triggers changes in hormones and gene expression through upregulation of abscisic acid (ABA), leading to increased osmotic adjustment to minimize salinity effects by maintaining foliar turgidity [6]. Osmotic adjustment may help cells regain water and turgidity [13].
- (2)
- Ion exclusion—ion (most often Na+) exclusion by plant roots prevents Na+ accumulation in leaves and avoids the toxic effect of higher ion concentrations, resulting in premature senescence and death of mature leaves and stunted growth of young leaves. Excluding excessive ions entails the up- and downregulation of ion channels and transporter genes to allow a tighter control of Na+ transport throughout the plant [3]. Ion exclusion occurs in some glycophytes and is very effective in halophytes. It happens right at the roots, preventing salts reaching the plant’s aerial parts. Development of saline vesicles at the leaf epidermis facilitates excretion of excessive salts to prevent accumulation in different plant organs. Part of this mechanism is due to salt deposition on mature leaves (leaf senescence), while avoiding accumulation in young developing leaves [6]. This mechanism avoids or postpones problems caused by Na+, but requires compensation through K+ uptake. Otherwise, there will be a greater need for organic solutes for osmotic adjustment. Again, synthesizing solutes jeopardizes the plant’s energy balance. Thus, the plant must be able to cope with both strategies of ion toxicity and turgor loss at the same time [3].
- (3)
- Tissue tolerance to ions—tissue tolerance involves a tolerance mechanism of Na+, or other cations, and in some species it involves Cl−. It is achieved at cellular and intracellular levels where salt ions are compartmentalized. This mechanism is to avoid toxic concentration in cytoplasm and more specifically in the mesophyll cells of the leaves [3]. Tissue tolerance to ions allows for increased survival and retention of mature leaves. It is achieved through synthesis and accumulation of compatible solutes (CS) [14]. The CS are vital for plant osmotolerance and provide adaptive roles in osmotic adjustment during salt accumulation [15]. However, the functions of CS are not limited to osmotic adjustment. They (CS) can also act as low molecular chaperons. Their hydrophilic property gives them the ability to replace water on protein or membrane surfaces [14,16] and herewith protect enzymes from being denatured and stabilize membranes. They also mitigate the effect of reactive oxygen species (ROS) on cell functions [16]. Compatible solutes, being small water-soluble molecules, when present in high concentrations, remain neutral with respect to cellular functions [17]. They consist of either nitrogen-rich compounds like amino acids, betaines and amines or sugars, polyols and organic acids [14,18].
2. Materials and Methods
2.1. Plant Materials
2.2. Phenotyping
2.3. Test Statistics
2.4. Genotyping
2.5. GWAS Models
3. Results
3.1. Variation of Salt Stress Tolerance During Germination
3.2. Salinity Tolerance among Geographical Origins
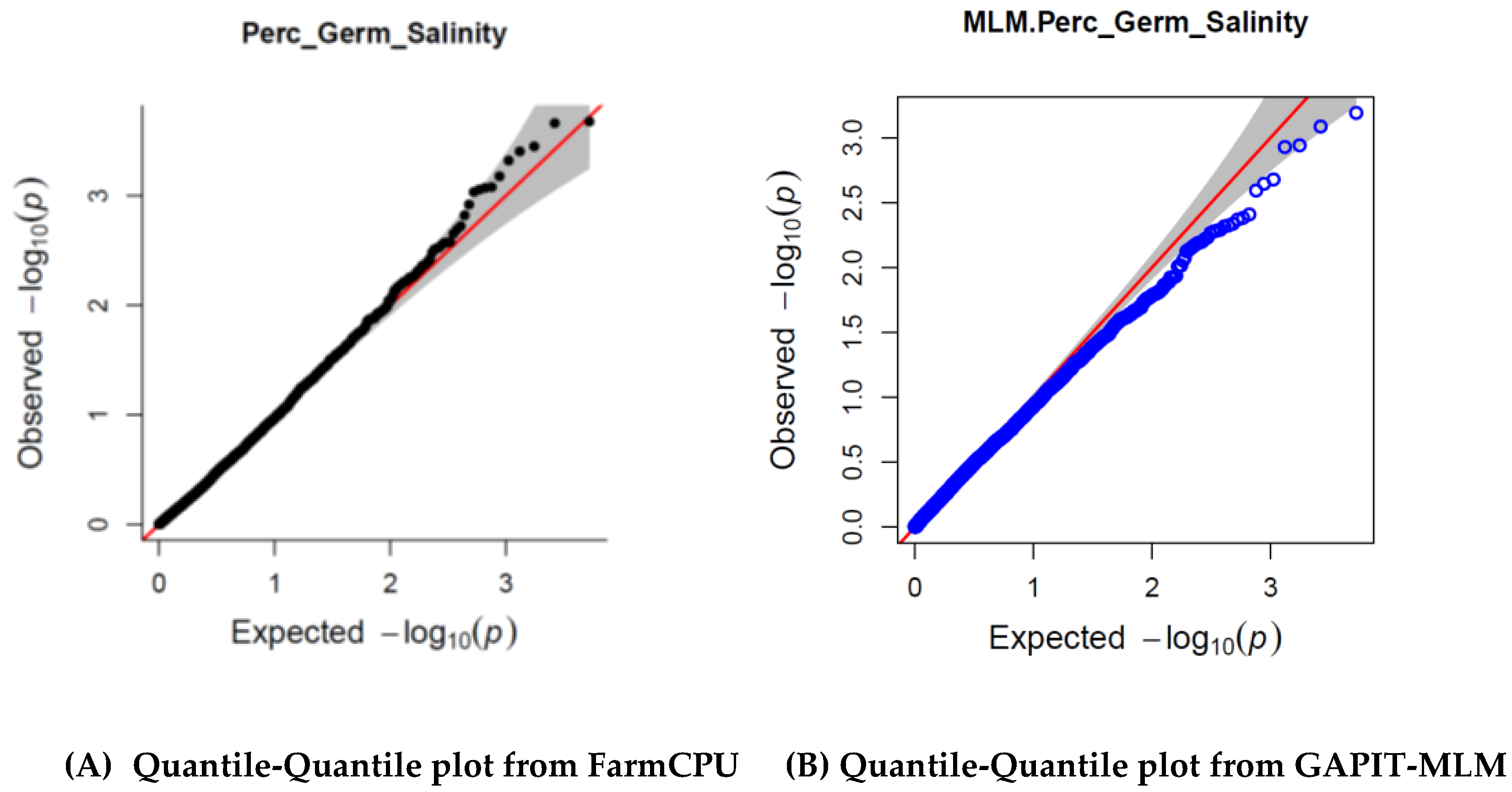
3.3. GWAS Model
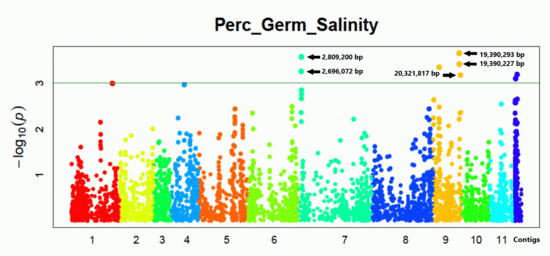
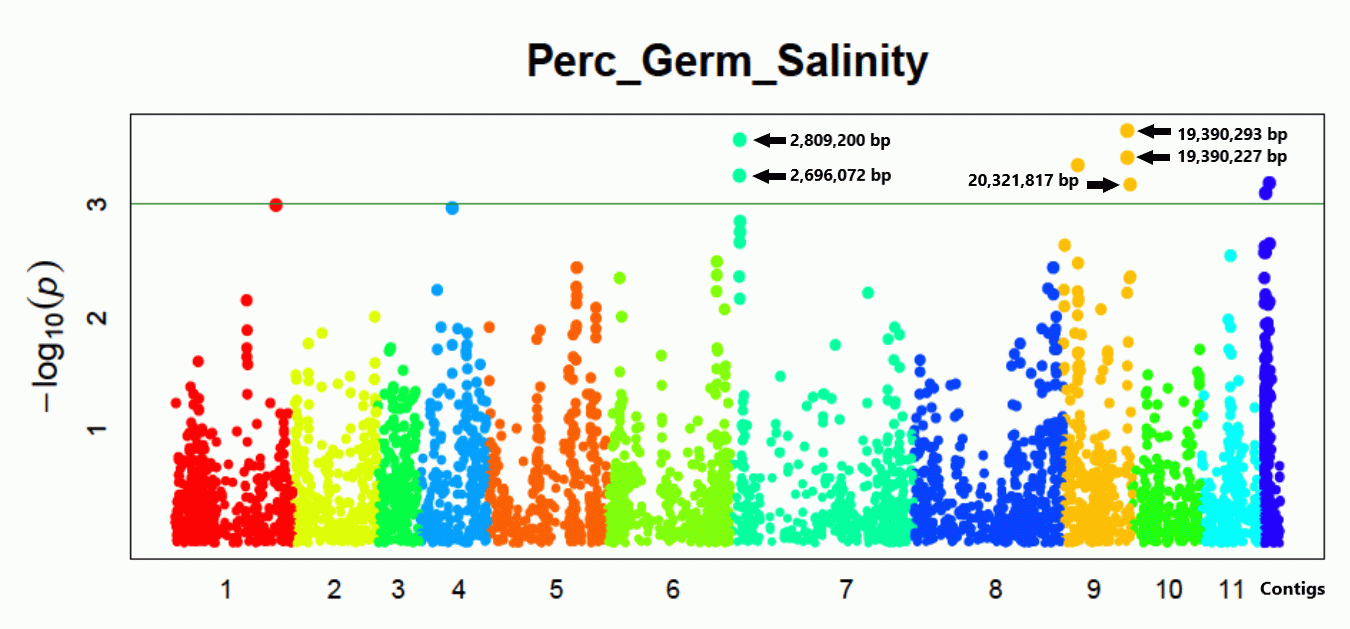
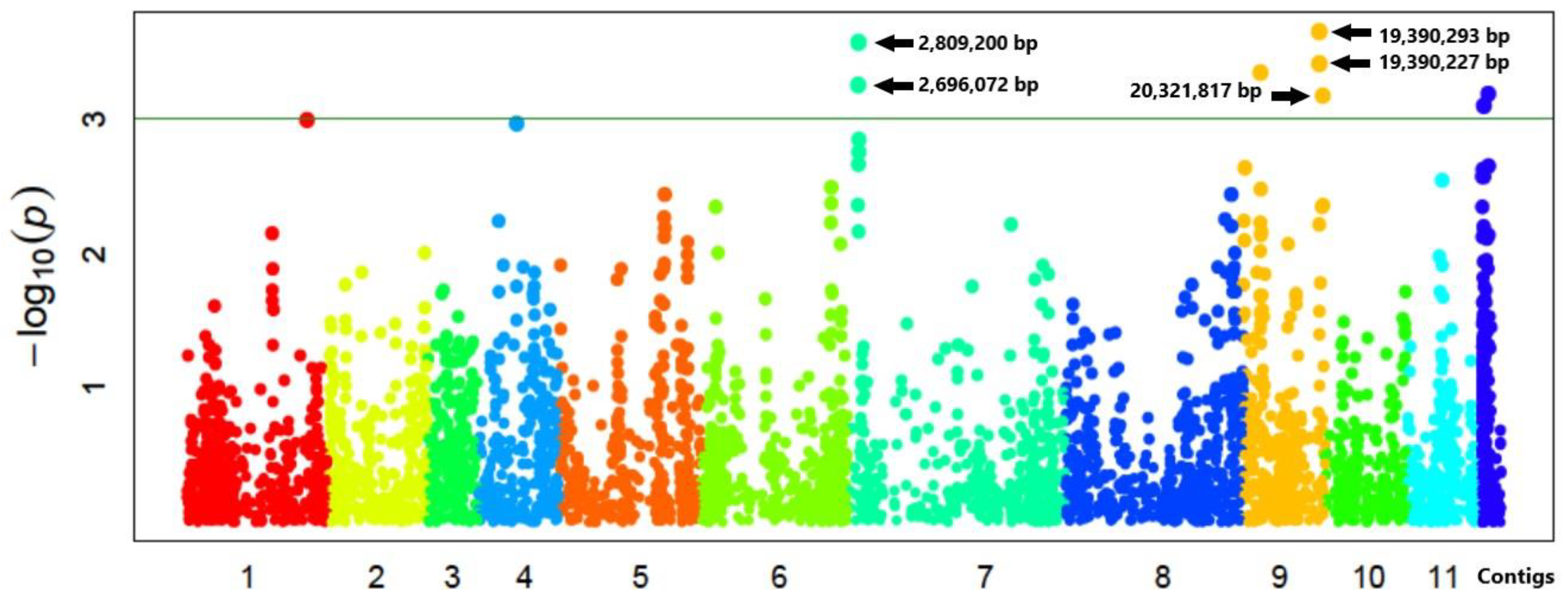
3.4. SNPs and Genes Associated with High Germination Rate under Salt Stress
4. Discussion
4.1. Optimal Salt Concentration to Distinguish Salt Tolerant and Susceptible Accessions Based on Germination Rate
4.2. Difference of Average Tolerance among Accessions Originating from Different Regions
4.3. Salt-Stress Loci and Putative Tolerance Genes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Tester, M.; Davenport, R. Na+ tolerance and Na+ transport in higher plants. Ann. Bot. 2003, 91, 503–527. [Google Scholar] [CrossRef]
- Pirasteh-Anosheh, H.; Ranjbar, G.; Pakniyat, H.; Emam, Y. Physiological mechanisms of salt stress tolerance in plants. In Plant-Environment Interaction; Baluska, F., Ed.; John Wiley & Sons: Chichester, UK, 2016; pp. 141–160. [Google Scholar]
- Munns, R.; Tester, M. Mechanisms of Salinity Tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef]
- Farooq, M.; Hussain, M.; Wakeel, A.; Siddique, K.H.M. Salt stress in maize: Effects, resistance mechanisms, and management. A review. Agron. Sustain. Dev. 2015, 35, 461–481. [Google Scholar] [CrossRef]
- Ma, Q.; Kang, J.; Long, R.; Zhang, T.; Xiong, J.; Zhang, K.; Wang, T.; Yang, Q.; Sun, Y. Comparative proteomic analysis of alfalfa revealed new salt and drought stress-related factors involved in seed germination. Mol. Biol. Rep. 2017, 44, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Acosta-Motos, J.R.; Ortuño, M.F.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.J.; Hernandez, J.A. Plant Responses to Salt Stress: Adaptive Mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef]
- Julkowska, M.M. Changing Gears in Salt Stress Tolerance: AtCAMTA6’s Role in Transcriptional Regulation of Ion Transport. Plant Physiol. 2019, 180, 684–685. [Google Scholar] [CrossRef]
- Lu, Y.; Lei, J.-Q.; Zeng, F.-J.; Zhang, B.; Liu, G.-J.; Liu, B.; Li, X.-Y. Effect of NaCl-induced changes in growth, photosynthetic characteristics, water status and enzymatic antioxidant system of Calligonum caput-medusae seedlings. Photosynthetica 2017, 55, 96–106. [Google Scholar] [CrossRef]
- Nadeem, M.; Li, J.; Yahya, M.; Wang, M.; Ali, A.; Cheng, A.; Wang, X.; Ma, C. Grain Legumes and Fear of Salt Stress: Focus on Mechanisms and Management Strategies. Int. J. Mol. Sci. 2019, 20, 799. [Google Scholar] [CrossRef]
- El Sayed, H.; El Sayed, A. Influence of NaCl and Na2SO4 treatments on growth development of broad bean (Vicia Faba, L.) plant. J. Life Sci. 2011, 5, 513–523. [Google Scholar]
- Gupta, B.; Huang, B. Mechanism of salinity tolerance in plants: Physiological, biochemical, and molecular characterization. Int. J. Genom. 2014, 2014, 701596. [Google Scholar] [CrossRef]
- Manchanda, G.; Garg, N. Salinity and its effects on the functional biology of legumes. Acta Physiol. Plant. 2008, 30, 595–618. [Google Scholar] [CrossRef]
- Passioura, J.B.; Munns, R. Rapid environmental changes that affect leaf water status induce transient surges or pauses in leaf expansion rate. Funct. Plant Biol. 2000, 27, 941–948. [Google Scholar] [CrossRef]
- Carillo, P.; Grazia, M.; Pontecorvo, G.; Fuggi, A.; Woodrow, P. Salinity Stress and Salt Tolerance. In Abiotic Stress in Plants—Mechanisms and Adaptations; Shanker, A., Venkateswarlu, B., Eds.; Indian Council of Agricultural Research: Basar, India, 2011. [Google Scholar]
- Ashraf, M.; Foolad, M.R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Hasegawa, P.M.; Bressan, R.A.; Zhu, J.-K.; Bohnert, H.J. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Biol. 2000, 51, 463–499. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, A.; Murata, N. The role of glycine betaine in the protection of plants from stress: Clues from transgenic plants. Plant Cell Environ. 2002, 25, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Mansour, M. Nitrogen containing compounds and adaptation of plants to salinity stress. Biol. Plant. 2000, 43, 491–500. [Google Scholar] [CrossRef]
- Dash, M.; Panda, S.K. Salt Stress Induced Changes in Growth and Enzyme Activities in Germinating Phaseolus Mungo Seeds. Biol. Plant. 2001, 44, 587–589. [Google Scholar] [CrossRef]
- Bonilla, I.; El-Hamdaoui, A.; Bolaños, L. Boron and calcium increasePisum sativum seed germination and seedling development under salt stress. Plant Soil 2004, 267, 97–107. [Google Scholar] [CrossRef]
- Kaya, M.; Kaya, G.; Kaya, M.D.; Atak, M.; Saglam, S.; Khawar, K.M.; Ciftci, C.Y. Interaction between seed size and NaCl on germination and early seedling growth of some Turkish cultivars of chickpea (Cicer arietinum L.). J. Zhejiang Univ. Sci. B 2008, 9, 371–377. [Google Scholar] [CrossRef]
- Nichols, P.G.H.; Malik, A.I.; Stockdale, M.; Colmer, T.D. Salt tolerance and avoidance mechanisms at germination of annual pasture legumes: Importance for adaptation to saline environments. Plant Soil 2008, 315, 241–255. [Google Scholar] [CrossRef]
- Tavakkoli, E.; Rengasamy, P.; McDonald, G.K. High concentrations of Na+ and Cl- ions in soil solution have simultaneous detrimental effects on growth of faba bean under salinity stress. J. Exp. Bot. 2010, 61, 4449–4459. [Google Scholar] [CrossRef] [PubMed]
- Iseki, K.; Takahashi, Y.; Muto, C.; Naito, K.; Tomooka, N. Diversity and Evolution of Salt Tolerance in the Genus Vigna. PLoS ONE 2016, 11, e0164711. [Google Scholar] [CrossRef] [PubMed]
- Garg, G. Response in germination and seedling growth in Phaseolus mungo under salt and drought stress. J. Environ. Biol. 2010, 31, 261–264. [Google Scholar] [PubMed]
- Nair, R.M.; Pandey, A.K.; War, A.R.; Hanumantharao, B.; Shwe, T.; Alam, A.; Pratap, A.; Malik, S.R.; Karimi, R.; Mbeyagala, E.K.; et al. Biotic and Abiotic Constraints in Mungbean Production—Progress in Genetic Improvement. Front. Plant Sci. 2019, 10, 1340. [Google Scholar] [CrossRef]
- Sehrawat, N.; Bhat, K.V.; Kaga, A.; Tomooka, N.; Yadav, M.; Jaiwal, P.K. Development of new gene-specific markers associated with salt tolerance for mungbean (Vigna radiata L. Wilczek). Span. J. Agric. Res. 2014, 12, 732–741. [Google Scholar] [CrossRef]
- Rabie, G.H. Influence of arbuscular mycorrhizal fungi and kinetin on the response of mungbean plants to irrigation with seawater. Mycorrhiza 2005, 15, 225–230. [Google Scholar] [CrossRef]
- Ahmed, S. Effect of salinity on the yield and yield components of mungbean. Pak. J. Bot. 2009, 41, 263–268. [Google Scholar]
- Zuccarini, P. Effects of silicon on photosynthesis, water relations and nutrient uptake of Phaseolus vulgaris under NaCl stress. Biol. Plant. 2008, 52, 157–160. [Google Scholar] [CrossRef]
- Cabot, C.; Sibole, J.V.; Barceló, J.; Poschenrieder, C. Abscisic Acid Decreases Leaf Na+ Exclusion in Salt-Treated Phaseolus vulgaris L. J. Plant Growth Regul. 2009, 28, 187–192. [Google Scholar] [CrossRef]
- Taïbi, K.; Taïbi, F.; Ait Abderrahim, L.; Ennajah, A.; Belkhodja, M.; Mulet, J.M. Effect of salt stress on growth, chlorophyll content, lipid peroxidation and antioxidant defence systems in Phaseolus vulgaris L. S. Afr. J. Bot. 2016, 105, 306–312. [Google Scholar] [CrossRef]
- Torche, Y.; Blair, M.; Saida, C. Biochemical, physiological and phenological genetic analysis in common bean (Phaseolus vulgaris L.) under salt stress. Ann. Agric. Sci. 2018, 63, 153–161. [Google Scholar] [CrossRef]
- Sehrawat, N.; Yadav, M.; Bhat, K.V.; Sairam, R.K.; Jaiwal, P.K. Effect of salinity stress on mungbean [Vigna radiata (L.) Wilczek] during consecutive summer and spring seasons. J. Agric. Sci. Belgrad. 2015, 60, 23–32. [Google Scholar] [CrossRef]
- Breria, C.M.; Hsieh, C.H.; Yen, J.-Y.; Nair, R.; Lin, C.-Y.; Huang, S.-M.; Noble, T.J.; Schafleitner, R. Population Structure of the World Vegetable Center Mungbean Mini Core Collection and Genome-Wide Association Mapping of Loci Associated with Variation of Seed Coat Luster. Trop. Plant Biol. 2019, 13, 1–12. [Google Scholar] [CrossRef]
- Schafleitner, R.; Nair, R.M.; Rathore, A.; Wang, Y.W.; Lin, C.Y.; Chu, S.H.; Lin, P.Y.; Chang, J.C.; Ebert, A.W. The AVRDC—The World Vegetable Center mungbean (Vigna radiata) core and mini core collections. BMC Genom. 2015, 16, 344. [Google Scholar] [CrossRef]
- Salt Tolerance Screening of the AVRDC Mungbean (Vigna Radiata) Collection.pdf. Available online: https://worldveg.tind.io//record/57265 (accessed on 18 January 2020).
- Liu, X.; Huang, M.; Fan, B.; Buckler, E.S.; Zhang, Z. Iterative Usage of Fixed and Random Effect Models for Powerful and Efficient Genome-Wide Association Studies. PLoS Genet. 2016, 12, e1005767. [Google Scholar] [CrossRef]
- Tang, Y.; Liu, X.; Wang, J.; Li, M.; Wang, Q.; Tian, F.; Su, Z.; Pan, Y.; Liu, D.; Lipka, A.E.; et al. GAPIT Version 2: An Enhanced Integrated Tool for Genomic Association and Prediction. Plant Genome 2016, 9, 1–9. [Google Scholar] [CrossRef]
- Kang, Y.J.; Satyawan, D.; Shim, S.; Lee, T.; Lee, J.; Hwang, W.J.; Kim, S.K.; Lestari, P.; Laosatit, K.; Kim, K.H.; et al. Draft genome sequence of adzuki bean, Vigna angularis. Sci. Rep. 2015, 5, 8069. [Google Scholar] [CrossRef]
- Kang, Y.J.; Kim, S.K.; Kim, M.Y.; Lestari, P.; Kim, K.H.; Ha, B.K.; Jun, T.H.; Hwang, W.J.; Lee, T.; Lee, J.; et al. Genome sequence of mungbean and insights into evolution within Vigna species. Nat. Commun. 2014, 5, 5443. [Google Scholar] [CrossRef]
- Maas, E.V. Plant growth response to salt stress. In Towards the Rational Use of High Salinity Tolerant Plants: Vol 1 Deliberations about High Salinity Tolerant Plants and Ecosystems; Lieth, H., Al Masoom, A.A., Eds.; Springer: Dordrecht, The Netherlands, 1993; pp. 279–291. [Google Scholar]
- Flowers, T.J.; Gaur, P.M.; Gowda, C.L.; Krishnamurthy, L.; Samineni, S.; Siddique, K.H.; Turner, N.C.; Vadez, V.; Varshney, R.K.; Colmer, T.D. Salt sensitivity in chickpea. Plantcell Environ. 2010, 33, 490–509. [Google Scholar] [CrossRef]
- Kotula, L.; Khan, H.A.; Quealy, J.; Turner, N.C.; Vadez, V.; Siddique, K.H.; Clode, P.L.; Colmer, T.D. Salt sensitivity in chickpea (Cicer arietinum L.): Ions in reproductive tissues and yield components in contrasting genotypes. Plantcell Environ. 2015, 38, 1565–1577. [Google Scholar] [CrossRef]
- Khan, H.A.; Siddique, K.H.; Munir, R.; Colmer, T.D. Salt sensitivity in chickpea: Growth, photosynthesis, seed yield components and tissue ion regulation in contrasting genotypes. J. Plant Physiol. 2015, 182, 1–12. [Google Scholar] [CrossRef]
- Khan, H.A.; Siddique, K.H.M.; Colmer, T.D. Vegetative and reproductive growth of salt-stressed chickpea are carbon-limited: Sucrose infusion at the reproductive stage improves salt tolerance. J. Exp. Bot. 2017, 68, 2001–2011. [Google Scholar] [CrossRef]
- Ren, S.; Lyle, C.; Jiang, G.; Penumala, A. Soybean Salt Tolerance 1 (GmST1) Reduces ROS Production, Enhances ABA Sensitivity, and Abiotic Stress Tolerance in Arabidopsis thaliana. Front. Plant Sci. 2016, 7, 445. [Google Scholar] [CrossRef] [PubMed]
- Shu, K.; Qi, Y.; Chen, F.; Meng, Y.; Luo, X.; Shuai, H.; Zhou, W.; Ding, J.; Du, J.; Liu, J.; et al. Salt Stress Represses Soybean Seed Germination by Negatively Regulating GA Biosynthesis While Positively Mediating ABA Biosynthesis. Front. Plant Sci. 2017, 8, 1372. [Google Scholar] [CrossRef] [PubMed]
- Palit, P.; Sasmal, B.C.; Bhattacharryya, A.C. Germplasm diversity and estimate of genetic advance of four morpho-physiological traits in a world collection of jute. Euphytica 1996, 90, 49–58. [Google Scholar]
- Loqué, D.; von Wirén, N. Regulatory levels for the transport of ammonium in plant roots. J. Exp. Bot. 2004, 55, 1293–1305. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.Y.; Siddiqi, M.Y.; Ruth, T.J.; Glass, A. Ammonium Uptake by Rice Roots (II. Kinetics of 13NH4+ Influx across the Plasmalemma). Plant Physiol. 1993, 103, 1259–1267. [Google Scholar] [CrossRef]
- Karasawa, T.; Hayakawa, T.; Mae, T.; Ojima, K.; Yamaya, T. Characteristics of ammonium uptake by rice cells in suspension culture. Soil Sci. Plant Nutr. 1994, 40, 333–338. [Google Scholar] [CrossRef]
- Kronzucker, H.J.; Britto, D.T.; Davenport, R.J.; Tester, M. Ammonium toxicity and the real cost of transport. Trends Plant Sci. 2001, 6, 335–337. [Google Scholar] [CrossRef]
- Howitt, S.M.; Udvardi, M.K. Structure, function and regulation of ammonium transporters in plants. Biochimica et Biophysica Acta (BBA) -Biomembranes 2000, 1465, 152–170. [Google Scholar] [CrossRef]
- Bu, Y.; Takano, T.; Liu, S. The role of ammonium transporter (AMT) against salt stress in plants. Plant Signal. Behav. 2019, 14, 1625696. [Google Scholar] [CrossRef] [PubMed]
- Bu, Y.; Sun, B.; Zhou, A.; Zhang, X.; Lee, I.; Liu, S. Identification and Characterization of a PutAMT1; 1 Gene from Puccinellia tenuiflora. PLoS ONE 2013, 8, e83111. [Google Scholar] [CrossRef] [PubMed]
- Ning, X.; Sun, Y.; Wang, C.; Zhang, W.; Sun, M.; Hu, H.; Liu, J.; Yang, L. A Rice CPYC-Type Glutaredoxin OsGRX20 in Protection against Bacterial Blight, Methyl Viologen and Salt Stresses. Front. Plant Sci. 2018, 9, 111. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Zhou, S.; Luo, Y.; Ma, C.; Gong, Y.; Zhou, Y.; Gao, S.; Huang, Z.; Yan, L.; Hu, Y.; et al. The heat shock protein 40 LeDnaJ regulates stress resistance and indole-3-acetic acid biosynthesis in Lentinula edodes. Fungal Genet. Biol. 2018, 118, 37–44. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Tolerance Level | Polystyrene Sheets | Petri Dish | Overall |
|---|---|---|---|
| Susceptible (0 to < 25%) | 11 | 17 | 5 |
| Moderately susceptible (25% to < 50%) | 122 | 122 | 135 |
| Moderately tolerant (50% to < 75%) | 125 | 127 | 140 |
| Tolerant (75% to 100%) | 39 | 31 | 17 |
| Parameter | Arid Semi-Arid | Tropical Subtropical |
|---|---|---|
| Mean | 45.413 | 51.838 |
| Variance | 82.922 | 118.403 |
| Observations | 246 | 38 |
| Hypothesized Mean Difference | 0.000 | - |
| df | 45 | - |
| t Stat | −3.458 | - |
| P(T ≤ t) two-tail | 0.001 | - |
| t Critical two-tail | 2.014 | - |
| Parameter | Germination % | Agro-Climatic Region | Subgroup |
|---|---|---|---|
| Germination % | 1 | ||
| Agro-climatic region | 0.228 ** | 1 | |
| Subgroup | −0.117 ** | −0.403 | 1 |
| Chr. | Position (bp) | Effect | LOD | Region Length (kbp) | Gene | Description Based on Arabidopsis |
|---|---|---|---|---|---|---|
| 7 | 2,809,200 | 3.803 | 3.660 | 113.128 | Vradi07g01630 | ammonium transporter protein (AMT) |
| 7 | 2,696,072 | −3.629 | 3.323 | |||
| 9 | 19,390,293 | −3.069 | 3.675 | 931.590 | Vradi09g09510 | OsGrx_S16-glutaredoxin subgroup II |
| 9 | 19,390,227 | 2.892 | 3.450 | Vradi09g09600 | dnaJ domain protein (HSP40s-heat shock protein) | |
| 9 | 20,321,817 | 2.963 | 3.178 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Breria, C.M.; Hsieh, C.-H.; Yen, T.-B.; Yen, J.-Y.; Noble, T.J.; Schafleitner, R. A SNP-Based Genome-Wide Association Study to Mine Genetic Loci Associated to Salinity Tolerance in Mungbean (Vigna radiata L.). Genes 2020, 11, 759. https://doi.org/10.3390/genes11070759
Breria CM, Hsieh C-H, Yen T-B, Yen J-Y, Noble TJ, Schafleitner R. A SNP-Based Genome-Wide Association Study to Mine Genetic Loci Associated to Salinity Tolerance in Mungbean (Vigna radiata L.). Genes. 2020; 11(7):759. https://doi.org/10.3390/genes11070759
Chicago/Turabian StyleBreria, Caleb Manamik, Ching-Hsiang Hsieh, Tsair-Bor Yen, Jo-Yi Yen, Thomas J. Noble, and Roland Schafleitner. 2020. "A SNP-Based Genome-Wide Association Study to Mine Genetic Loci Associated to Salinity Tolerance in Mungbean (Vigna radiata L.)" Genes 11, no. 7: 759. https://doi.org/10.3390/genes11070759
APA StyleBreria, C. M., Hsieh, C.-H., Yen, T.-B., Yen, J.-Y., Noble, T. J., & Schafleitner, R. (2020). A SNP-Based Genome-Wide Association Study to Mine Genetic Loci Associated to Salinity Tolerance in Mungbean (Vigna radiata L.). Genes, 11(7), 759. https://doi.org/10.3390/genes11070759