The Q223R Polymorphism of the Leptin Receptor Gene as a Predictor of Weight Gain in Childhood Obesity and the Identification of Possible Factors Involved

 ,
,  ,
,  , , and
, , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Ethical Statement
2.3. Outcome Variables
2.3.1. Anthropometric Measurements
2.3.2. Dietary and Physical Activity Data
2.3.3. Selection of Single Nucleotide Polymorphisms
2.4. DNA Extraction and Genotyping
2.5. Statistical Analysis
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bischoff, S.C.; Boirie, Y.; Cederholm, T.; Chourdakis, M.; Cuerda, C.; Delzenne, N.M.; Deutz, N.E.; Fouque, D.; Genton, L.; Gil, C.; et al. Towards a multidisciplinary approach to understand and manage obesity and related diseases. Clin. Nutr. 2017, 36, 917–938. [Google Scholar] [CrossRef]
- Abarca-Gómez, L.; Abdeen, Z.A.; Hamid, Z.A.; Abu-Rmeileh, N.M.; Acosta-Cazares, B.; Acuin, C.; Adams, R.J.; Aekplakorn, W.; Afsana, K.; Aguilar-Salinas, C.A.; et al. Worldwide trends in body-mass index, underweight, overweight, and obesity from 1975 to 2016: A pooled analysis of 2416 population-based measurement studies in 128·9 million children, adolescents, and adults. Lancet 2017, 390, 2627–2642. [Google Scholar] [CrossRef]
- Zheng, M.; Lamb, K.E.; Grimes, C.; Laws, R.; Bolton, K.; Ong, K.K.; Campbell, K. Rapid weight gain during infancy and subsequent adiposity: A systematic review and meta-analysis of evidence. Obes. Rev. 2018, 19, 321–332. [Google Scholar] [CrossRef] [PubMed]
- Thiering, E.; Brüske, I.; Kratzsch, J.; Hoffmann, B.; Herbarth, O.; von Berg, A.; Schaaf, B.; Wichmann, H.-E.; Heinrich, J.; LISAplus Study Group. Peak growth velocity in infancy is positively associated with blood pressure in school-aged children. J. Hypertens. 2012, 30, 1114. [Google Scholar] [CrossRef] [PubMed]
- Sweeting, A.N.; Caterson, I.D. Approaches to obesity management. Intern. Med. J. 2017, 47, 734–739. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.J.; Geisler, C.; Blundell, J.; Dulloo, A.; Schutz, Y.; Krawczak, M.; Bosy-Westphal, A.; Enderle, J.; Heymsfield, S.B. The case of GWAS of obesity: Does body weight control play by the rules? Int. J. Obes. 2018, 42, 1395–1405. [Google Scholar] [CrossRef]
- Felix, J.F.; Bradfield, J.P.; Monnereau, C.; van der Valk, R.J.P.; Stergiakouli, E.; Chesi, A.; Gaillard, R.; Feenstra, B.; Thiering, E.; Kreiner-Møller, E.; et al. Genome-wide association analysis identifies three new susceptibility loci for childhood body mass index. Hum. Mol. Genet. 2016, 25, 389–403. [Google Scholar] [CrossRef]
- Thunders, M.; Mangai, S.; Cooper, R. Nutrigenetics, nutrigenomics, and the future of dietary advice. Food Nutr. Sci. 2013, 4, 999–1003. [Google Scholar] [CrossRef]
- Renta Neta Media de Los Hogares (Urban Audit)—Ayuntamiento de Madrid. Available online: http://www.madrid.es/portales/munimadrid/es/Inicio/El-Ayuntamiento/Estadistica/Areas-de-informacion-estadistica/Economia/Renta/Renta-neta-media-de-los-hogares-Urban-Audit-?vgnextfmt=default&vgnextoid=65e0c19a1666a510VgnVCM1000001d4a900aRCRD&vgnextchannel=ef863636b44b4210VgnVCM2000000c205a0aRCRD (accessed on 22 December 2019).
- Stewart, A.; Marfell-Jones, M.; International Society for Advancement of Kinanthropometry. International Standards for Anthropometric Assessment; International Society for the Advancement of Kinanthropometry: Lower Hutt, New Zealand, 2011; ISBN 978-0-620-36207-8. [Google Scholar]
- WHO. Physical Status: The Use and Interpretation of Anthropometry. Available online: http://www.who.int/childgrowth/publications/physical_status/en/ (accessed on 22 December 2019).
- Cole, T.J.; Bellizzi, M.C.; Flegal, K.M.; Dietz, W.H. Establishing a standard definition for child overweight and obesity worldwide: International survey. BMJ 2000, 320, 1240–1243. [Google Scholar] [CrossRef]
- European Food Safety Authority. General principles for the collection of national food consumption data in the view of a pan-European dietary survey. EFSA J. 2009, 7. [Google Scholar] [CrossRef]
- Ortega, R.; López-Sobaler, A.; Andrés, P.; Requejo, A.; Aparicio, A.; Molinero, L. DIAL Software for Assessing Diets and Food Calculations (for Windows, Version 3.3.6.0); Department of Nutrition (UCM) & Alce Ingeniería, S.L.: Madrid, Spain, 2019; Available online: http://www.alceingenieria.net/nutricion/descarga.htm (accessed on 5 April 2019).
- Ortega, R.; Requejo, A.; López-Sobaler, A. Modelos de cuestionario de actividad. In Nutriguía. Manual de Nutrición Clínica en Atención Primaria; Complutense: Madrid, Spain, 2006; p. 468. [Google Scholar]
- World Health Organization. Protein and energy requirements: A joint FAO/WHO memorandum. Bull. World Health Organ. 1979, 57, 65–79. [Google Scholar]
- Ortiz-Marrón, H.; Ortiz-Pinto, M.A.; Cuadrado-Gamarra, J.I.; Esteban-Vasallo, M.; Cortés-Rico, O.; Rey-Gayo, L.; Ordobás, M.; Galán, I.; Ortiz-Marrón, H.; Ortiz-Pinto, M.A.; et al. Persistence and Variation in Overweight and Obesity Among the Pre-school Population of the Community of Madrid After 2 Years of Follow-up. The ELOIN Cohort. Rev. Esp. Cardiol. 2018, 71, 902–909. [Google Scholar] [CrossRef] [PubMed]
- Agencia Española de Consumo, Seguridad Alimentaria y Nutrición. Ministerio de Sanidad, Servicios Sociales e Igualdad 2015. Estudio ALADINO 2015: Estudio de Vigilancia del Crecimiento, Alimentación, Actividad Física, Desarrollo Infantil y Obesidad en España. 2016. Available online: http://www.aecosan.msssi.gob.es/AECOSAN/docs/documentos/nutricion/observatorio/Estudio_ALADINO_2015.pdf (accessed on 5 April 2019).
- Frongillo, E.A.; Lampl, M. Early identification of children at risk of developing obesity. Arch. Pediatr. Adolesc. Med. 2011, 165, 1043–1044. [Google Scholar] [CrossRef] [PubMed]
- Corella, D.; Coltell, O.; Portolés, O.; Sotos-Prieto, M.; Fernández-Carrión, R.; Ramirez-Sabio, J.B.; Zanón-Moreno, V.; Mattei, J.; Sorlí, J.V.; Ordovas, J.M. A guide to applying the sex-gender perspective to nutritional genomics. Nutrients 2018, 11, 4. [Google Scholar] [CrossRef] [PubMed]
- Gruzdeva, O.; Borodkina, D.; Uchasova, E.; Dyleva, Y.; Barbarash, O. Leptin resistance: Underlying mechanisms and diagnosis. Diabetes Metab. Syndr. Obes. 2019, 12, 191–198. [Google Scholar] [CrossRef]
- Dornbush, S.; Aeddula, N.R. Physiology, Leptin. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2019. [Google Scholar]
- Daghestani, M.; Purohit, R.; Daghestani, M.; Daghistani, M.; Warsy, A. Molecular dynamic (MD) studies on Gln233Arg (rs1137101) polymorphism of leptin receptor gene and associated variations in the anthropometric and metabolic profiles of Saudi women. PLoS ONE 2019, 14, e0211381. [Google Scholar] [CrossRef]
- Eldosouky, M.K.; Abdu Allah, A.M.; AbdElmoneim, A.; Al-Ahmadi, N.S. Correlation between serum leptin and its gene expression to the anthropometric measures in overweight and obese children. Cell. Mol. Biol. 2018, 64, 84–90. [Google Scholar] [CrossRef]
- Mărginean, C.O.; Mărginean, C.; Voidăzan, S.; Meliţ, L.; Crauciuc, A.; Duicu, C.; Bănescu, C. Correlations between leptin gene polymorphisms 223 A/G, 1019 G/A, 492 G/C, 976 C/A, and anthropometrical and biochemical parameters in children with obesity: A prospective case-control study in a Romanian population-the nutrichild study. Medicine 2016, 95, e3115. [Google Scholar] [CrossRef]
- Murugesan, D.; Arunachalam, T.; Ramamurthy, V.; Subramanian, S. Association of polymorphisms in leptin receptor gene with obesity and type 2 diabetes in the local population of Coimbatore. Indian J. Hum. Genet. 2010, 16, 72–77. [Google Scholar] [CrossRef]
- Mattevi, V.S.; Zembrzuski, V.M.; Hutz, M.H. Association analysis of genes involved in the leptin-signaling pathway with obesity in Brazil. Int. J. Obes. 2002, 26, 1179–1185. [Google Scholar] [CrossRef]
- Quinton, N.; Lee, A.; Ross, R.; Eastell, R.; Blakemore, A. A single nucleotide polymorphism (SNP) in the leptin receptor is associated with BMI, fat mass and leptin levels in postmenopausal Caucasian women. Hum. Genet. 2001, 108, 233–236. [Google Scholar] [CrossRef] [PubMed]
- Verkerke, H.; Naylor, C.; Zabeau, L.; Tavernier, J.; Petri, W.A., Jr.; Marie, C. Kinetics of leptin binding to the Q223R leptin receptor. PLoS ONE 2014, 9, e94843. [Google Scholar] [CrossRef] [PubMed]
- Almeida, S.M.; Furtado, J.M.; Mascarenhas, P.; Ferraz, M.E.; Ferreira, J.C.; Monteiro, M.P.; Vilanova, M.; Ferraz, F.P. Association between LEPR, FTO, MC4R, and PPARG-2 polymorphisms with obesity traits and metabolic phenotypes in school-aged children. Endocrine 2018, 60, 466–478. [Google Scholar] [CrossRef] [PubMed]
- Pyrzak, B.; Wisniewska, A.; Kucharska, A.; Wasik, M.; Demkow, U. No association of LEPR Gln223Arg polymorphism with leptin, obesity or metabolic disturbances in children. Eur. J. Med. Res. 2009, 14, 201–204. [Google Scholar] [CrossRef]
- Riestra, P.; García-Anguita, A.; Schoppen, S.; López-Simón, L.; De Oya, M.; Garcés, C. Sex-specific association between leptin receptor polymorphisms and leptin levels and BMI in healthy adolescents. Acta Paediatr. 2010, 99, 1527–1530. [Google Scholar] [CrossRef]
- Bender, N.; Allemann, N.; Marek, D.; Vollenweider, P.; Waeber, G.; Mooser, V.; Egger, M.; Bochud, M. Association between Variants of the Leptin Receptor Gene (LEPR) and overweight: A systematic review and an analysis of the colaus study. PLoS ONE 2011, 6, e26157. [Google Scholar] [CrossRef]
- Péneau, S.; Giudici, K.V.; Gusto, G.; Goxe, D.; Lantieri, O.; Hercberg, S.; Rolland-Cachera, M.-F. Growth trajectories of body mass index during childhood: Associated factors and health outcome at adulthood. J. Pediatr. 2017, 186, 64–71. [Google Scholar] [CrossRef]
- Plank, L.D.; Obolonkin, V.; Smith, M.; Savila, F.; Jalili-Moghaddam, S.; Tautolo, E.-S.; Rush, E.C. Pacific Islands Families Study: Physical growth to age 14 and metabolic risk. Pediatr. Obes. 2019, 14, e12497. [Google Scholar] [CrossRef]
- Gallicchio, L.; Chang, H.H.; Christo, D.K.; Thuita, L.; Huang, H.Y.; Strickland, P.; Ruczinski, I.; Clipp, S.; Helzlsouer, K.J. Single nucleotide polymorphisms in obesity-related genes and all-cause and cause-specific mortality: A prospective cohort study. BMC Med. Genet. 2009, 10, 103. [Google Scholar] [CrossRef]
- van Rossum, C.T.M.; Hoebee, B.; van Baak, M.A.; Mars, M.; Saris, W.H.M.; Seidell, J.C. Genetic variation in the leptin receptor gene, leptin, and weight gain in young dutch adults. Obes. Res. 2003, 11, 377–386. [Google Scholar] [CrossRef]
- van Rossum, C.T.M.; Hoebee, B.; Seidell, J.C.; Bouchard, C.; van Baak, M.A.; de Groot, C.P.G.M.; Chagnon, M.; de Graaf, C.; Saris, W.H.M. Genetic factors as predictors of weight gain in young adult Dutch men and women. Int. J. Obes. Relat. Metab. Disord. 2002, 26, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Zandoná, M.R.; Rodrigues, R.O.; Albiero, G.; Campagnolo, P.D.B.; Vitolo, M.R.; Almeida, S.; Mattevi, V.S. Polymorphisms in LEPR, PPARG and APM1 genes: Associations with energy intake and metabolic traits in young children. Arq. Bras. Endocrinol. Metabol. 2013, 57, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Shalitin, S.; Phillip, M. Role of obesity and leptin in the pubertal process and pubertal growth--a review. Int. J. Obes. Relat. Metab. Disord. 2003, 27, 869–874. [Google Scholar] [CrossRef] [PubMed]
- Gajewska, J.; Kuryłowicz, A.; Mierzejewska, E.; Ambroszkiewicz, J.; Chełchowska, M.; Weker, H.; Puzianowska-Kuźnicka, M. Complementary effects of genetic variations in LEPR on body composition and soluble leptin receptor concentration after 3-month lifestyle intervention in prepubertal obese children. Nutrients 2016, 8, 328. [Google Scholar] [CrossRef] [PubMed]
- Ceddia, R.B. Direct metabolic regulation in skeletal muscle and fat tissue by leptin: Implications for glucose and fatty acids homeostasis. Int. J. Obes. 2005, 29, 1175–1183. [Google Scholar] [CrossRef] [PubMed]
- Walsh, S.; Haddad, C.J.; Kostek, M.A.; Angelopoulos, T.J.; Clarkson, P.M.; Gordon, P.M.; Moyna, N.M.; Visich, P.S.; Zoeller, R.F.; Seip, R.L.; et al. Leptin and leptin receptor genetic variants associate with habitual physical activity and the arm body composition response to resistance training. Gene 2012, 510, 66–70. [Google Scholar] [CrossRef] [PubMed]
- Tu, W.; Wagner, E.K.; Eckert, G.J.; Yu, Z.; Hannon, T.; Pratt, J.H.; He, C. Associations between menarche-related genetic variants and pubertal growth in male and female adolescents. J. Adolesc. Health 2015, 56, 66–72. [Google Scholar] [CrossRef]
- Foster, E.; Bradley, J. Methodological considerations and future insights for 24-hour dietary recall assessment in children. Nutr. Res. 2018, 51, 1–11. [Google Scholar] [CrossRef]



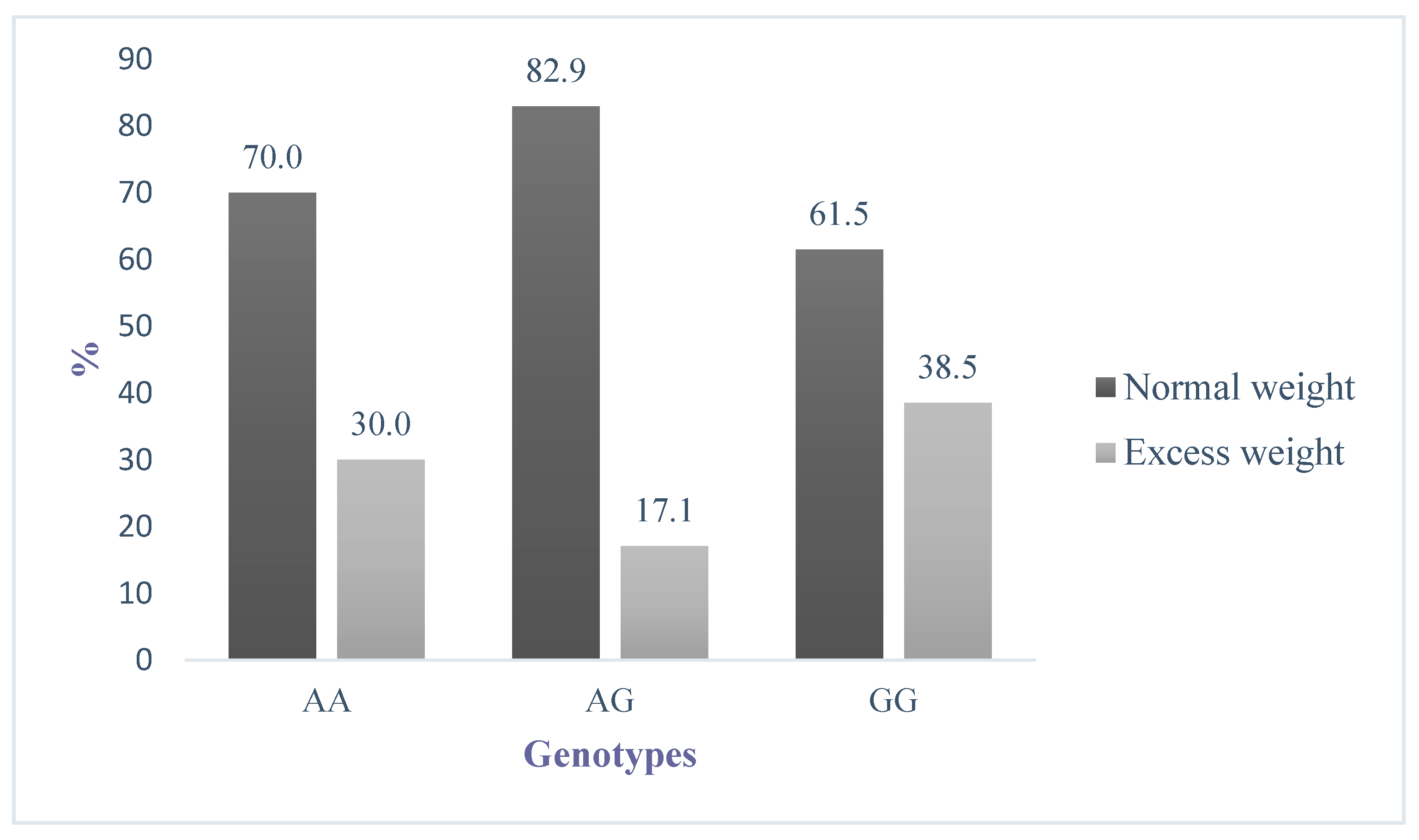{kind=link}
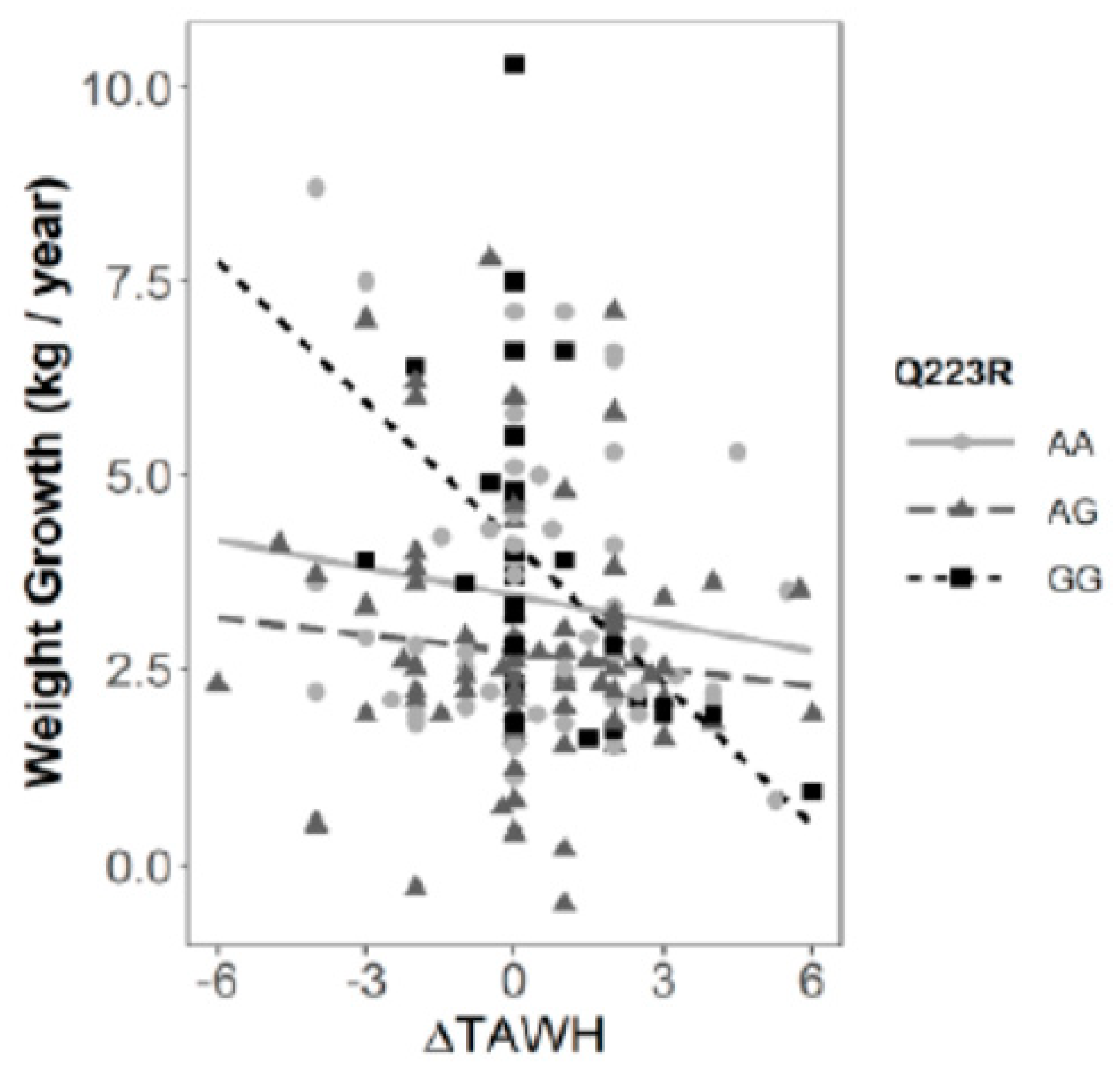{kind=link}
| Anthropometric Variables | AA 1 | AG 1 | GG 1 | β (CI) 2 | p |
|---|---|---|---|---|---|
| Weight growth (kg/year) | 3.52 ± 1.79 | 2.8 ± 1.49 | 3.99 ± 2.08 | −0.72 (−1.26, −0.18); 0.42 (−0.3, 1.13) | 0.001 |
| Height growth (cm/year) | 5.77 ± 0.85 | 5.56 ± 1.06 | 5.94 ± 1,00 | −0.21 (−0.52, 0.1); 0.23 (−0.18, 0.64) | 0.063 |
| %V BMI | 3.36 ± 5.04 | 1.63 ± 4.30 | 3.63 ± 4.87 | −1.73 (−3.23, −0.23); 0.24 (−1.76, 2.23) | 0.027 |
| %V FM | 2.37 ± 13.71 | 0,00 ± 14.72 | 1.91 ± 11.76 | −2.21 (−6.72, 2.3); −1.09 (−7.06, 4.89) | 0.623 |
| %V MM | 6.81 ± 4.87 | 7.74 ± 5.05 | 6.76 ± 5.04 | 0.62 (−0.92, 2.15); 0.51 (−1.43, 2.45) | 0.723 |
| %V WC | 3.8 ± 4.35 | 1.51 ± 3.91 | 3.7 ± 4.13 | −2.3 (−3.61, −0.99); −0.16 (−1.91, 1.6) | <0.001 |
| %V Triceps fold | 2.76 ± 17.55 | −1.93 ± 15.68 | 3.57 ± 12.86 | −4.78 (−9.88, 0.33); 1.17 (−5.64, 7.99) | 0.075 |
| Q223R LEPR YEAR 1 1 | Q223R LEPR YEAR 2 1 | |||||
|---|---|---|---|---|---|---|
| AA (WT) | AG | GG | AA (WT) | AG | GG | |
| Anthropometric variables | ||||||
| Weight (kg) | 26.61 ± 5.80 | 25.79 ± 5.48 | 29.06 ± 7.27 | 29.55 ± 6.58 | 28.52 ± 6.33 | 32.63 ± 8.83 |
| Height (cm) | 124.8 ± 6.39 | 124.08 ± 6.28 | 126.77 ± 6.40 | 130.13 ± 6.65 | 129.74 ± 6.43 | 132.47 ± 6.32 |
| BMI (kg/m2) | 16.98 ± 2.79 | 16.58 ± 2.30 | 17.85 ± 3.06 | 17.33 ± 3.00 | 16.8 ± 2.49 | 18.32 ± 3.60 |
| Fat mass (%) | 20.86 ± 7.50 | 19.69 ± 6.40 | 22.79 ± 8.40 | 21.07 ± 7.89 | 19.41 ± 6.38 | 22.46 ± 9.17 |
| Muscle mass (%) | 28.07 ± 3.14 | 27.92 ± 3.05 | 28.29 ± 2.27 | 29.57 ± 2.79 | 29.86 ± 2.45 | 30.14 ± 1.95 |
| WC (cm) | 59.57 ± 6.96 | 58.88 ± 6.69 | 62.77 ± 8.79 | 61.42 ± 8.09 | 59.47 ± 7.29 | 64.09 ± 9.79 |
| Triceps fold (mm) | 12.75 ± 5.28 | 11.68 ± 4.60 | 14.27 ± 5.53 | 12.89 ± 5.70 | 11.35 ± 4.97 | 14.4 ± 6.53 |
| Dietetic variables | ||||||
| Caloric Intake (kJ) | 8046.57 ± 1385.79 | 7634.55 ± 1376.41 | 7548.55 ± 1502.81 | 8322.52 ± 1637.54 | 8110.42 ± 1495.15 | 7668.04 ± 1670.83 |
| Lipids (g) | 85.45 ± 21.09 | 78.64 ± 18.66 | 77.27 ± 23.17 | 86.81 ± 23.27 | 85.77 ± 22.27 | 75.5 ± 16.95 |
| SFA (g) | 28.71 ± 8.38 | 27.03 ± 7.57 | 26.84 ± 8.30 | 29.1 ± 8.38 | 28.64 ± 8.50 | 25.39 ± 6.48 |
| Proteins (g) | 78.89 ± 16.05 | 74.68 ± 13.35 | 75.59 ± 12.59 | 88.05 ± 21.33 | 86.45 ± 17.27 | 84.64 ± 28.65 |
| Carbohydrates (g) | 199.97 ± 34.53 | 194.82 ± 46.06 | 193.21 ± 36.53 | 204.27 ± 45.57 | 195.61 ± 42.8 | 195.24 ± 45.87 |
| Physical activity variables | ||||||
| TAWH (hours/week) | 4.14 ± 1.96 | 3.68 ± 1.75 | 3.33 ± 1.63 | 4.75 ± 2.35 | 4.09 ± 2.42 | 4.3 ± 2.05 |
| IPAC | 1.59 ± 0.12 | 1.59 ± 0.10 | 1.57 ± 0.11 | 1.52 ± 0.08 | 1.53 ± 0.09 | 1.51 ± 0.08 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marcos-Pasero, H.; Aguilar-Aguilar, E.; Colmenarejo, G.; Ramírez de Molina, A.; Reglero, G.; Loria-Kohen, V. The Q223R Polymorphism of the Leptin Receptor Gene as a Predictor of Weight Gain in Childhood Obesity and the Identification of Possible Factors Involved. Genes 2020, 11, 560. https://doi.org/10.3390/genes11050560
Marcos-Pasero H, Aguilar-Aguilar E, Colmenarejo G, Ramírez de Molina A, Reglero G, Loria-Kohen V. The Q223R Polymorphism of the Leptin Receptor Gene as a Predictor of Weight Gain in Childhood Obesity and the Identification of Possible Factors Involved. Genes. 2020; 11(5):560. https://doi.org/10.3390/genes11050560
Chicago/Turabian StyleMarcos-Pasero, Helena, Elena Aguilar-Aguilar, Gonzalo Colmenarejo, Ana Ramírez de Molina, Guillermo Reglero, and Viviana Loria-Kohen. 2020. "The Q223R Polymorphism of the Leptin Receptor Gene as a Predictor of Weight Gain in Childhood Obesity and the Identification of Possible Factors Involved" Genes 11, no. 5: 560. https://doi.org/10.3390/genes11050560
APA StyleMarcos-Pasero, H., Aguilar-Aguilar, E., Colmenarejo, G., Ramírez de Molina, A., Reglero, G., & Loria-Kohen, V. (2020). The Q223R Polymorphism of the Leptin Receptor Gene as a Predictor of Weight Gain in Childhood Obesity and the Identification of Possible Factors Involved. Genes, 11(5), 560. https://doi.org/10.3390/genes11050560