1. Introduction
Erythritol is a four-carbon polyol, which is used as a sugar substitute. It gained popularity as a sweetener because of interesting properties such as a very low caloric value, little impact on insulin release [
1], and indications that its usage might lower the prevalence of caries-related mouth bacteria [
2]. Unlike many synthetic sweeteners, it can be found naturally in food such as fruits or honey [
3]. Yet, the role of this compound in metabolism is still unclear. Erythritol is often described as a zero-calorie sweetener, due to its fast absorption in the small intestine and subsequent excretion of most of the intake with the urine [
4]. Initially, it was believed that erythritol was not metabolized in the human body. However, the most recent studies revealed that a small fraction of the intake could be processed to erythronate [
5]. Moreover, erythritol could be synthesized from glucose in the blood and mammalian tissues [
5,
6], and in some cases elevated erythritol levels in blood might be a marker of upcoming changes in the health condition [
5,
7]. Thus, there is a need for better understanding of the role of this polyol in metabolism.
As erythritol is becoming a more widely available food additive, it is necessary to ensure that long-term usage of this sweetener does not have any negative impact on the human body and microflora. It is a difficult task because many aspects of erythritol metabolism remain unclear even in single-celled organisms. In this study, we focused on the role of erythritol in the yeast
Yarrowia lipolytica. This yeast is capable of both the synthesis [
8] and utilization [
9] of erythritol; thus it might become a model of erythritol metabolism in eukaryotic cells. Other yeast genera that are known to produce significant amounts of erythritol are
Trigonopsis,
Pichia,
Moniliella,
Pseudozyma,
Aureobasidium, and
Trichoderma [
3]. Some of them have interesting industrial applications or are a part of the human microbiome [
10].
Y. lipolytica is a dimorphic, oleaginous yeast that is being increasingly studied due to its great potential for industrial applications [
11]. It produces erythritol under hyperosmotic conditions [
12]. The biosynthesis of erythritol proceeds mainly via the pentose phosphate pathway (PPP) [
3]. Erythrose-4-phosphate, the metabolite of PPP, is dephosphorylated by a still unknown enzyme to erythrose, which is later converted to erythritol by erythrose reductase [
13]. After depletion of the original carbon source in the growth medium, the concentration of erythritol starts to decrease. There are four enzymes known to be involved in erythritol utilization: erythritol dehydrogenase Eyd1 [
14], kinase Eyk1 [
15], and two isomerases, Eyi1 and Eyi2 [
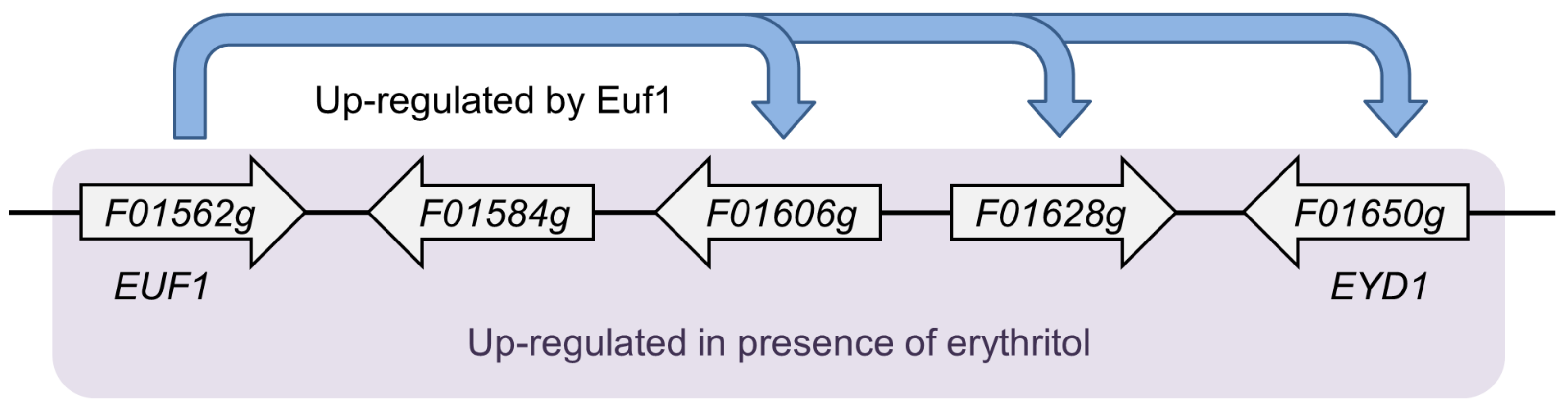
16]. The genes encoding these proteins are regulated by the transcription factor Euf1. All of them, including
EUF1, are arranged in a cluster in the genome called the erythritol utilization cluster (
Figure 1). Euf1 is most likely not the only factor regulating erythritol utilization, since in its absence the
euf1Δ strain may still use erythritol as a carbon source to a small extent [
9].
The correlation between high osmotic pressure and erythritol production indicates that it might be an unusual stress response. After years of studies carried out on the model yeast
Saccharomyces cerevisiae [
17] and the pathogenic yeast
Candida albicans [
18], it is known that yeast responses to various environmental stresses are regulated by the network of signaling pathways. The response to osmotic stress is transmitted through the high osmolarity glycerol (HOG) pathway. The central part of the pathway is a cascade of protein kinases (MAPK cascade) and its final element is the protein kinase Hog1. Activation of the HOG pathway leads to the phosphorylation of Hog1. In this activated form, it interacts with numerous cell elements in both the cytoplasm and the nucleus, which leads to adaptation to the environment. A significant part of the response is intracellular accumulation of small molecules with a neutral charge known as osmolytes. That balances the osmotic pressure on both sides of the membrane and prevents the efflux of water. In the case of
S. cerevisiae the main osmolyte is glycerol [
19].
Y. lipolytica has a homologue of kinase Hog1 and its deletion leads to a high sensitivity to osmotic stress. Moreover, the strain
yl-hog1Δ shows a few other anomalies, e.g., increased filamentous growth and higher resistance to cell wall disturbing agents [
20]. Similar changes have been previously observed in
hog1Δ deletion strains of
S. cerevisiae [
21] and
C. albicans [
22], which indicate the similarities of signaling pathways. However, the final responses are different, as the osmolyte produced by
Y. lipolytica is erythritol rather than glycerol. The intracellular concentration of erythritol rises several-fold in hyperosmotic conditions [
23], but surprisingly a significant amount of this polyol is also secreted to the environment.
In this study, we investigated the importance of erythritol in osmoprotection. We used an osmo-sensitive strain that carries the deletion of yl-Hog1, and combined it with modifications of genes involved in erythritol metabolism.
2. Materials and Methods
2.1. Strains and Culture Conditions
Strains used in the study were
Y. lipolytica AJD, the
ura3Δ derivative of
Y. lipolytica A101, and its further derivatives. They were obtained from the Department of Biotechnology and Food Microbiology at Wrocław University of Environmental and Life Sciences, Poland. Strains of
Escherichia coli DH5α were the carriers of plasmids containing deletion or overexpression cassettes and were used for transformation procedures. The full list of used strains and plasmids is in
Table 1.
E. coli strains used in the transformation procedures were cultivated in LB medium. YPD medium, containing 1% (w/v) yeast extract (Merck, Darmstadt, Germany), 1% (w/v) peptone (Merck, Darmstadt, Germany), and 2% (w/v) glucose (Chempur, Piekary Śląskie, Poland), was used for yeast inoculum preparation. YNB (Sigma-Aldrich, Darmstadt, Germany) agar plates with 2% (w/v) glucose were used for standard yeast transformations. YPD agar plates with Hygromycin B (800 µL/100 mL) were used for yeast transformations to restore auxotrophy. YPD agar plates with 2% (w/v) NaCl were used to screen for osmo-sensitive strains. YNB agar plates with varying concentrations of carbon sources were used in stress sensitivity tests. The carbon sources were: glucose, glycerol (Wratislavia-Biodiesel, Wroclaw, Poland), or erythritol (Młyn Oliwski, Gdańsk, Poland). The plates for stress sensitivity tests were also supplemented with various concentrations of additions such as erythritol, mannitol (Sigma-Aldrich, Darmstadt, Germany), and L-proline (Sigma-Aldrich, Darmstadt, Germany). Glycerol, glucose, and erythritol used in the media were sterilized by filtration and added to other compounds, just before the preparation of the agar plates. All yeast strains were cultivated at 30 °C.
Shake-flask experiments were performed in 0.3 L flasks with baffles containing 0.05 L of medium kept on a rotary shaker at 240 rpm. Media used for these experiments were: YNB medium with 100 g/L glycerol and optional supplementation of 10 g/L erythritol. 3 g L−1 CaCO3 was also added in order to prevent a drop in pH during culture.
2.2. Analytical Methods
Osmolality of some media was measured on the osmometer OS 3000 Marcel, Poland. The samples were diluted 10-fold before measurement. The concentrations of polyols and organic acids were determined by HPLC using a HyperRez Carbohydrate H+ Column (Thermo Scientific, Waltham, MA, USA) coupled to a UV (λ = 210 nm) (Dionex, Sunnyvale, CA, USA) and a refractive index detector (Shodex, Ogimachi, Japan). 0.25% trifluoroacetic acid was used as a mobile phase solvent. The samples were diluted 10-fold before the measurement. Data were analyzed with the Chromeleon program. Samples for biomass measurements (5 mL) were collected from shake-flask cultures and harvested by filtration on 0.45 µm pore size membranes. The biomass was determined gravimetrically after drying in a drier at 105 °C.
2.3. Deletion and Overexpression Cassettes
Plasmids containing deletion or overexpression cassettes prepared in previous studies were purified from
E. coli strains on a Plasmid Mini kit (A&A Biotechnology, Gdynia, Poland). The
yl-hog1 deletion cassette was carried on plasmid pQE80-ptHog1+Ura [
20], the
euf1 deletion cassette was on plasmid pUC-ura-ΔF01562 [
9], and the
EUF1 overexpression cassette was on plasmid pAD-F01562 [
9].
The deletion cassette for the gene
eyd1 was prepared as follows. The gene YALI0F01650 promoter region was amplified by PCR with Phusion high-fidelity DNA polymerase (Thermo Scientific, Darmstadt, Germany) and primers p1650-F-PmeI (CCAGTTTAAACCTCCAAGAGCTGCCACTGTAG) and p1650-R-NotI (TTAGCGGCCGCGGAAACTGTTGTCAGTATTTG). The obtained PCR product was digested with the enzymes MssI and NotI and inserted into corresponding sites of the plasmid pUC-Ura [
9] with the ligation enzyme T4 DNA Ligase (Thermo Scientific, Darmstadt, Germany), resulting in the vector pUC-Ura-pF01650. The proper integration was tested by PCR reaction with primers p1650-col-F (GGAATAAGGCTGGTCATGATGG) and p1650-col-R (CAGAGTACGGCTTCTCAATCG). Transformation of
E. coli strains was performed using standard chemical protocols [
27].
The YALI0F01650 terminator fragment was amplified with primers t1650-F-ApaI (ATTGGGCCCTCATCCCGACTGACAGCTAAC) and t1650-R-HindIII (GATAAGCTTTCAGC GGGAAGCTGGAGAG). The PCR product was digested with the enzymes ApaI and HindIII and inserted into the plasmid pUC-URA-pF01650. The final plasmid pUC-Ura-ΔF01650 was tested for proper integration with PCR reaction with the primers t1650-col-F (AATAATCTCGGGACAGCAATG) and t1650-col-R (TTGTGTAGGGCTTACTTGATG).
2.4. Yeast Transformation
Transformation of
Y. lipolytica was performed according to the lithium acetate method [
28]. The strain AJD with auxotrophy for uracil was transformed with one of the plasmids containing an overexpression or deletion cassette and transformants were plated out on selective media without uracil. They were tested for proper integration by gDNA extraction and PCR reaction. The uracil auxotrophy was restored by transformation with the replicative plasmid pUB4-Cre1 [
25] with the Cre-lox recombinase system. The transformants were plated out on YPD medium with hygromycin B. After restoring the auxotrophy and loss of the replicative plasmid, strains were transformed with the plasmid pQE80-ptHog1+Ura [
20]. The proper integration was confirmed with PCR reaction on isolated gDNA and increased sensitivity to growth on YPD plates with 2% NaCl.
The strain AJD was also transformed with the plasmid pAD. The plasmid was used for preparation of overexpression cassettes. However, in this case the cassette was empty and the plasmid was only allowed to integrate the gene URA3 into the rDNA sequence. The resulting AJDU strain was used as a control, instead of the original A101, because removal of URA3 from the original locus caused some differences in growth rate between A101 and AJD strains.
2.5. Stress Sensitivity Tests
Y. lipolytica strains were grown to the exponential phase (OD600 = 0.6) in liquid YPD medium. The dilutions 1, 10−1, 10−2, and 10−3 were spotted on YNB agar plates, with 0.5 M, 0.75 M, or 1 M concentration of one of the carbon sources—glucose, glycerol, or erythritol. YNB media with glucose and glycerol were also supplemented with erythritol (0.01–0.1 M), mannitol (0.1 M), or L-proline. Photos of the plates were taken after 24, 48, and 72 h of cultivation. The tests were performed in duplicate.
3. Results
Previous research on yl-Hog1 in
Y. lipolytica has shown that the deletion strain
yl-hog1Δ is very sensitive to osmotic stress induced by addition of NaCl to the growth media [
20]. In this study, we focused on high concentrations of carbon sources as a stressing factor. The initially chosen carbon sources were glucose, glycerol, and erythritol. The osmotic pressure of the medium is related to the molar concentration of its compounds. Thus, we compared media with the same molar concentration of the carbon source. It should be noted that glycerol, glucose, and erythritol have different molar masses, and the concentrations given in g L
−1 are significantly different. There was also a possibility that the chosen carbon sources might interact differently with other compounds of the media, or there might be losses during the sterilization process. To prevent such factors influencing further research, the osmolality values of YNB medium with 0.5 M carbon sources were measured. The result was 780 ± 34 mOsm kg
−1 H
2O for medium with glycerol, 733 ± 41 mOsm kg
−1 H
2O for glucose and 743 ± 49 mOsm kg
−1 H
2O for erythritol.
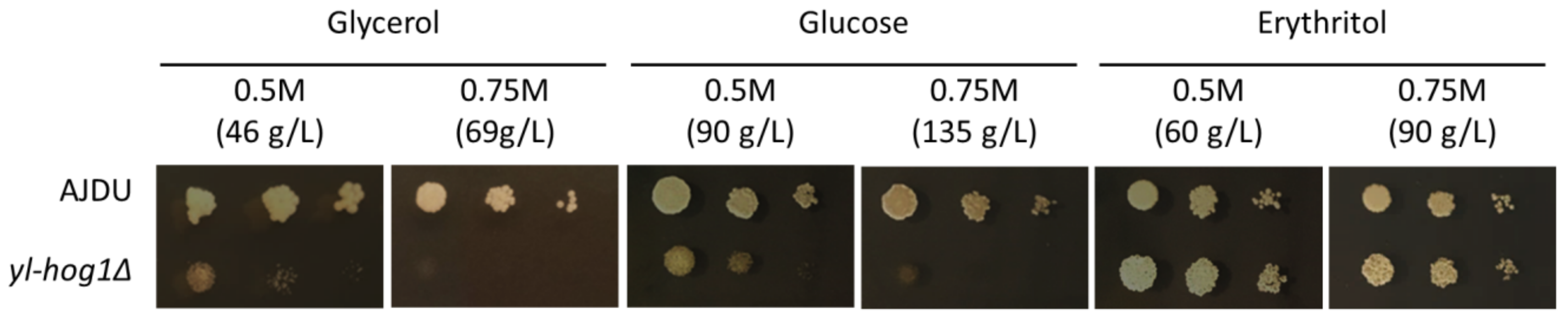
First, it has to be determined what concentration of carbon sources might inhibit the growth of strain
yl-hog1Δ. The sensitivity of
Y. lipolytica AJDU (control) and AJD
yl-hog1Δ was tested on YNB plates containing 0.5 M or 0.75 M of one of the substrates: glucose, glycerol, or erythritol (
Figure 2). The cultures were carried out for at least 72 h. Photos presented in the figures were taken in the 48th hour, as the changes between tested strains were the most clearly visible.
Changes in the concentration had little effect on the growth of the control AJDU strain, but the strain
yl-hog1Δ was very vulnerable to a rise in substrate concentration. The impact of media containing glucose and glycerol was similar: 0.5 M was enough to severely diminish the growth compared to the control strain and the growth was completely inhibited in 0.75 M. However, the results for erythritol-containing media were surprisingly different. On 0.5 M erythritol, the strain
yl-hog1Δ grew better than the control. Colonies on 0.75 M were slightly smaller, but the growth was still not weaker than the control. To ensure that the weaker growth of strain
yl-hog1Δ on glycerol and glucose was caused by their too high concentrations, we additionally prepared plates with 0.3 M carbon sources (
Supplementary Materials, Figure S1). With these lower concentrations, the development of strain
yl-hog1Δ on glycerol and glucose was significantly improved.
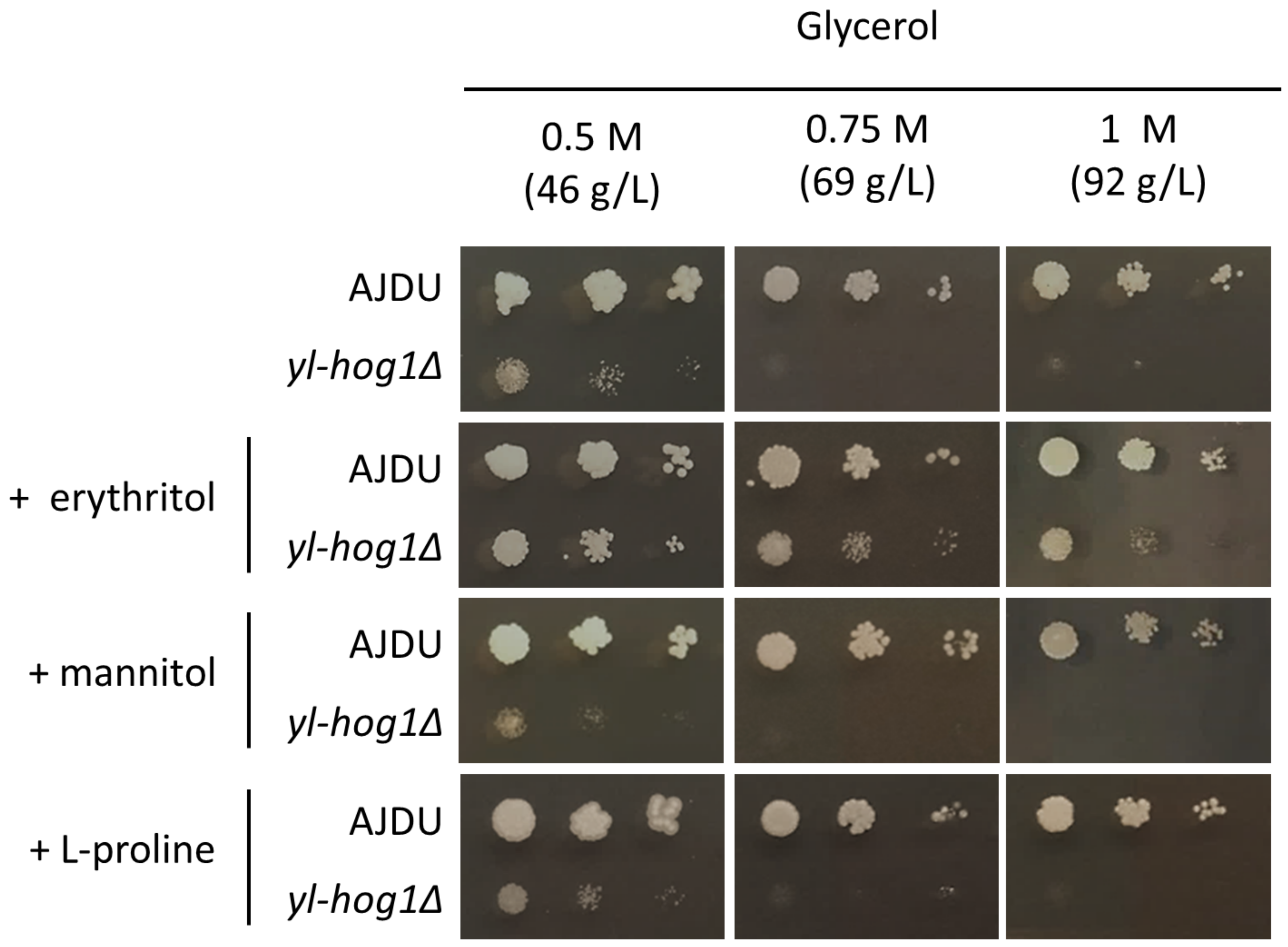
The next question was whether erythritol might improve the growth of
yl-hog1Δ when mixed with another substrate that generates hyperosmotic stress. To test this possibility, YNB agar plates with 0.5 M, 0.75 M, or 1 M glycerol and 0.1 M erythritol supplementation were prepared. As observed in a previous experiment, 0.75 M concentration of glycerol could completely inhibit the development of
yl-hog1Δ. However, when the erythritol was also present in the medium, the deletion strain could endure not only 0.75 M glycerol, but also 1 M (
Figure 3). The addition of erythritol did not completely overcome the osmotic stress though, as the strain
yl-hog1Δ was still significantly weaker than the control.
There was a possibility that the beneficial influence on
yl-hog1Δ might be achieved by supplementing the hyperosmotic media with other compounds with osmoprotective properties. The most common osmolyte for yeast is glycerol, but in the case of
yl-hog1Δ it was used as a stressing factor. From the group of other potential osmoprotectors we decided to use mannitol and L-proline. Mannitol, like erythritol and glycerol, is a polyol. Moreover, it is one of the main byproducts of erythritol production by
Y. lipolytica [
23]. Proline is an osmoprotectant for many groups of organisms [
29,
30], and a several-fold rise of its intracellular concentration was also observed in
Y. lipolytica cells subjected to osmotic stress [
31]. Mannitol was added to YNB-glycerol medium to the concentration 0.1 M, similar to erythritol, but the supplementation did not improve the growth of
yl-hog1Δ. L-proline was added in a lower concentration (0.02 M), because as an amino acid, it is a source of not only carbon, but also nitrogen. Thus, the supplementation was expected to have a large impact on yeast growth. Colonies of the control strain were indeed bigger, but
yl-hog1Δ was still not able to grow in higher concentrations of glycerol. To ensure that the difference between erythritol and proline supplementation was not the result of a lower concentration of proline, YNB-glycerol plates with erythritol supplementation from 0.01 to 0.05 M were prepared. The concentration of 0.02 M erythritol could still improve the growth of
yl-hog1Δ, although to a lesser extent than 0.1 M concentrations.
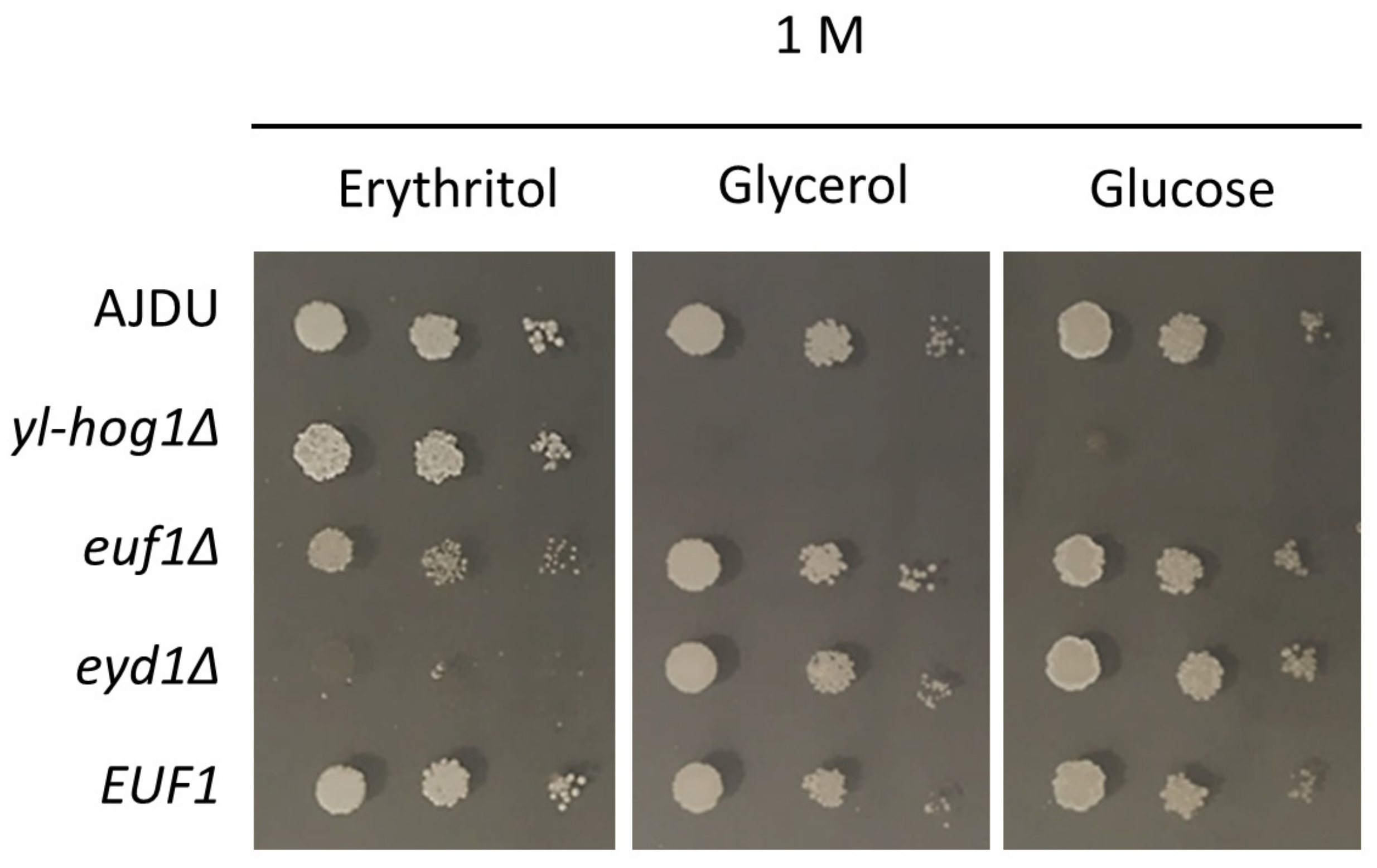
Next, we attempted to learn more about the mechanism of the protection. Osmolytes are known to be accumulated inside cells to balance the high osmotic pressure of the environment. To test the importance of erythritol accumulation, we targeted two proteins associated with its utilization: the transcription factor Euf1 and the enzyme Eyd1.
The two deletion strains
euf1Δ and
eyd1Δ were created on an AJD background.
EUF1 was also overexpressed in the strain
Y. lipolytica EUF1. The properties of the new transformants were tested on YNB plates with 1 M erythritol, glucose or glycerol as a sole carbon source (
Figure 4). Both Euf1 and Eyd1 are known to be important for erythritol utilization. However, cultivation on erythritol as a single carbon source revealed the difference in the phenotype of deletion strains. The growth of
euf1Δ was impaired, but still possible, whereas the development of
eyd1Δ was completely inhibited. Overexpression of
EUF1 did not cause any significant changes compared to the control. To ensure that the weak growth of
euf1Δ and
eyd1Δ was not a result of stress caused by a high concentration of the substrate, all transformants were also grown on glucose and glycerol. In these conditions, the growth of strains
euf1Δ and
eyd1Δ and
EUF1 did not differ from the control.
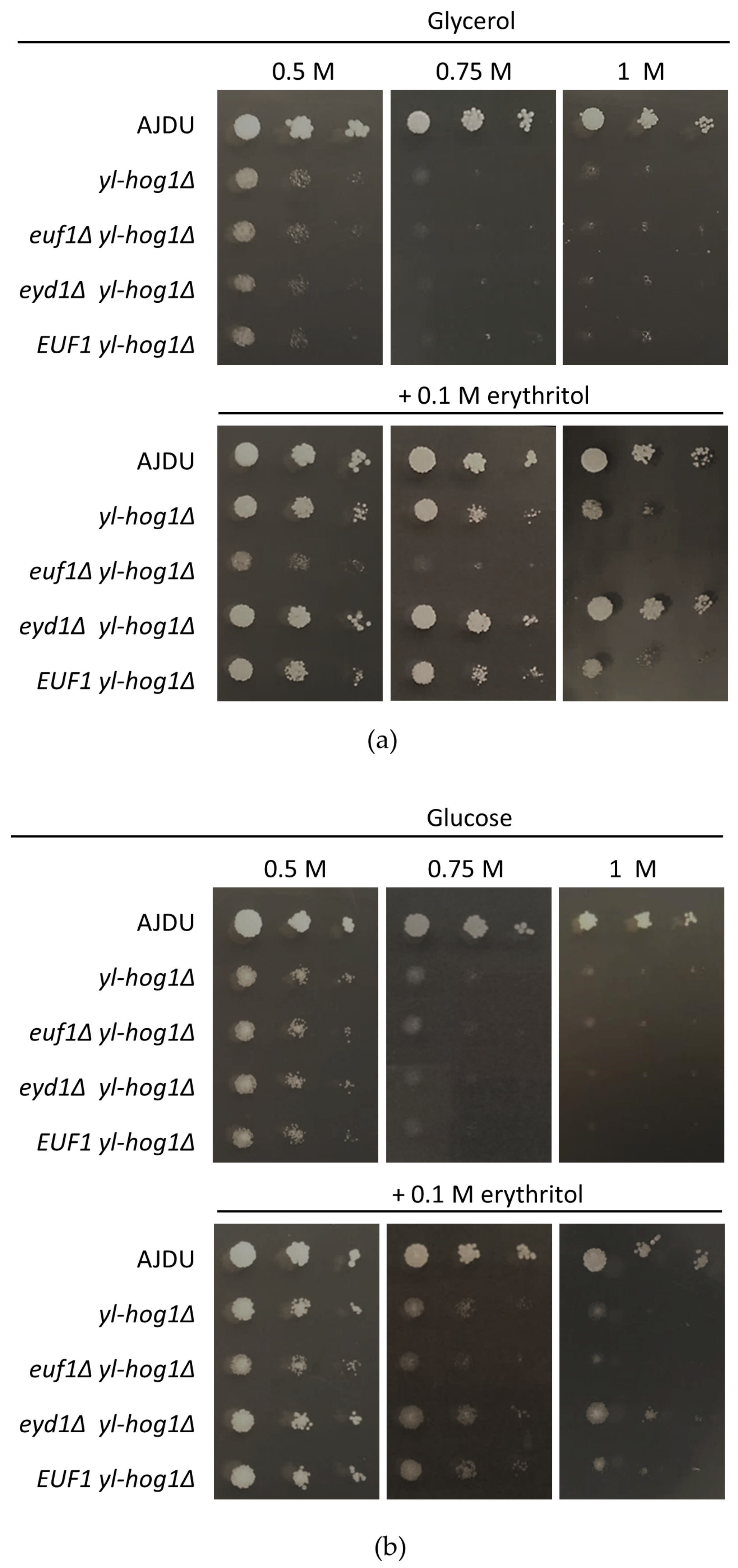
After establishing that
EUF1 and
EYD1 deletions do not influence the sensitivity to osmotic stress, the deletion yl-Hog1 was added. The strains
euf1Δ yl-hog1Δ,
eyd1Δ yl-hog1Δ, and
EUF1 yl-hog1Δ were all unable to grow at glucose and glycerol levels higher than 0.5 M (
Figure 5). However, the addition of erythritol revealed significant differences. The strain
euf1Δ yl-hog1Δ did not react to supplementation. Its growth was exactly the same as on plates without erythritol: weak on 0.5 M carbon sources and completely inhibited at higher concentrations. It was a striking contrast to all other tested strains. On the other hand,
eyd1Δ yl-hog1Δ reacted to the supplementation more strongly than
yl-hog1Δ. On the plates with glycerol as a main carbon source, the growth of
eyd1Δ yl-hog1Δ was comparable to the control strain, regardless of the glycerol concentrations (
Figure 5a). On glucose plates,
eyd1Δ yl-hog1Δ could withstand higher glucose concentrations than other deletion strains, but there was still strong growth inhibition compared to the control strain (
Figure 5b). The last created strain,
EUF1 yl-hog1Δ, did not differ from
yl-hog1Δ in all tested conditions.
The result that should be underlined is the significant difference between the effects of the erythritol supplementation on glucose and glycerol media. When glucose was used as the main carbon source (
Figure 5b), the positive influence of erythritol was significantly weaker, although still visible.
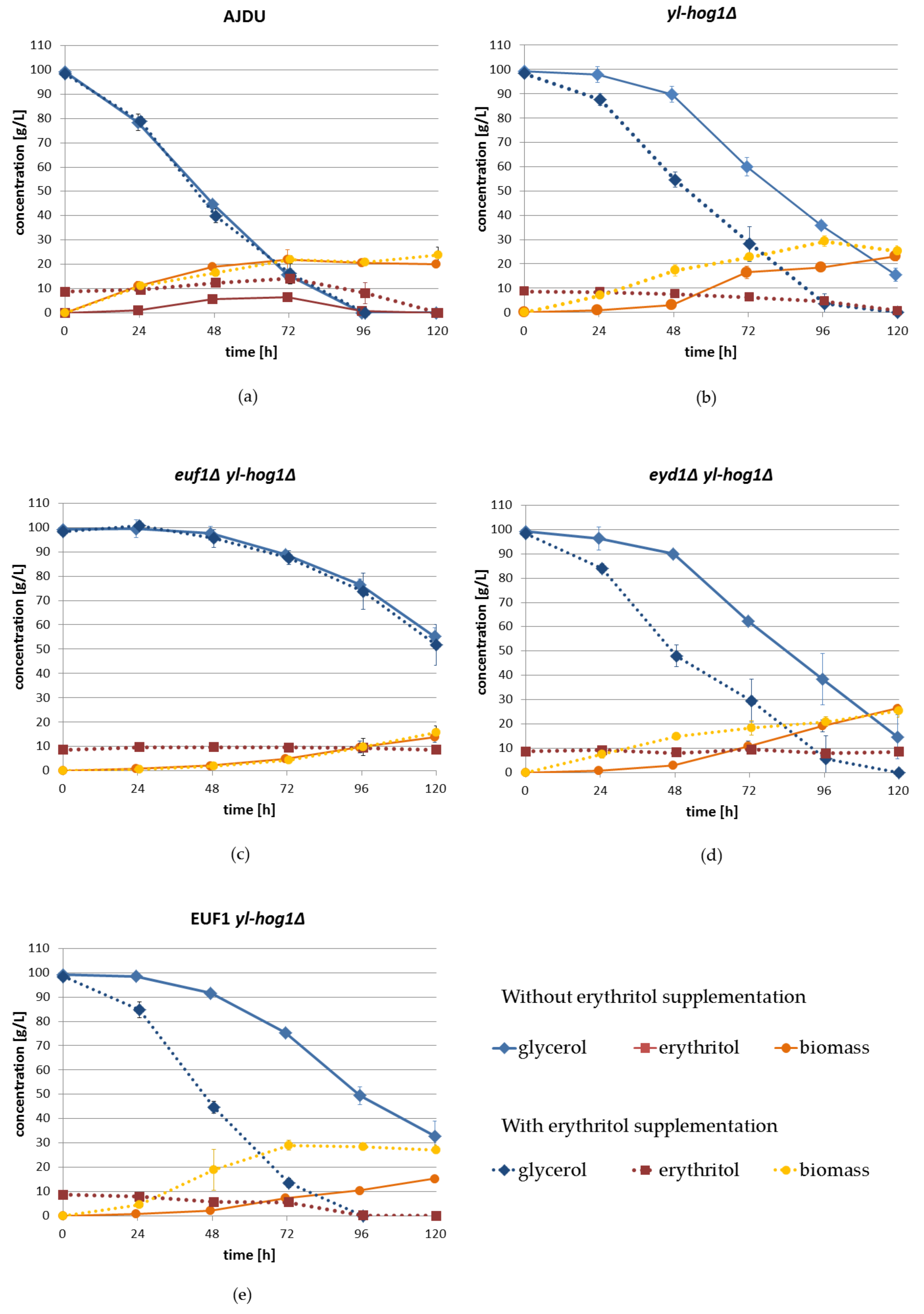
To further investigate the impact of erythritol supplementation on growth, substrate utilization, and secretion, a shake-flask experiment with liquid YNB medium with 100 g L
−1 glycerol (1.08 M) was prepared. Erythritol was supplemented in the concentration 10 g L
−1 (0.08 M). The unit g L
−1 was used because it is easier to monitor the carbon flux. Similar to previous experiments, the erythritol supplementation did not significantly influence the cultures of
Y. lipolytica AJDU (
Figure 6a). The addition of erythritol did not change the speed of glycerol utilization. Erythritol was produced, regardless of its initial presence in the medium. Its concentration reached 6.5 ± 0.3 g L
−1 without supplementation and 14.1 ± 0.9 g L
−1 with supplementation. In both cases erythritol was utilized after the depletion of glycerol.
In shake-flask cultures, all strains with deletion of yl-hog1 were found to be less sensitive to the osmotic stress generated by glycerol than in plate cultures (
Figure 6). They were able to adapt to the conditions, even without erythritol supplementation, but there was still a significant delay compared to the control AJDU strain.
During the first 48 h of cultivation without supplementation, the increase in biomass and utilization of glycerol by the strain
yl-hog1Δ were low, but significant changes appeared on the third day of cultivation (
Figure 6b). By the end of the experiment (120 h), the biomass reached 23.2 ± 2.1 g L
−1, and 15.5 ± 2.6 g L
−1 of glycerol remained. The addition of erythritol accelerated the rate of glycerol utilization and the biomass increase. Both glycerol and erythritol were completely exhausted by the end of the experiment. Biomass reached its highest value at 96 h, 29.3 ± 2.2 g L
−1, which was higher than that of the AJDU control.
Similar to the agar plate sensitivity tests, in shake-flask cultures the strain
euf1Δ yl-hog1Δ was also unable to use erythritol supplementation to improve adaptation to the osmotic stress (
Figure 6c). The average rates of glycerol utilization and biomass growth were very similar in cultures both with and without the addition of erythritol. Moreover, the lack of euf1 resulted in significant deterioration of the adaptability in comparison to the strain
yl-hog1Δ. After 48 h, the utilization rate of glycerol remained very slow, and at the end of the experiment there was still 55.2 ± 3.6 g of L
−1 of glycerol in culture without erythritol and 51.7 ± 8.4 g L
−1 in cultures with supplementation. Biomass growth was also extremely slow compared to all other strains, and at 120 h it was 13.7 ± 2.0 g L
−1 and 15.6 ± 2.7 g L
−1, respectively.
The results for the strain
eyd1Δ yl-hog1Δ were similar to those for
yl-hog1Δ. In the absence of erythritol, at the end of the experiment, there was 14.7 ± 7.3 g L
−1 remaining glycerol, while with supplementation, glycerol was completely depleted at 120 h (
Figure 6d). The concentration of supplemented erythritol remained constant throughout the entire experiment, due to the deletion of
EYD1. The inability to utilize erythritol could be the reason for lower final biomass (25.43 ± 2.6 g L
−1) than for
yl-hog1Δ.Erythritol supplementation had the most significant impact on glycerol utilization by strain EUF1
yl-hog1Δ (
Figure 6e). In the absence of erythritol, the adaptation was slower compared to
yl-hog1Δ and
eyd1Δ yl-hog1Δ. After 120 h there was 32.7 ± 6.3 g L
−1 of glycerol and biomass concentration was 15.3 ± 1 g L
−1. When erythritol was in the medium, both glycerol and erythritol were completely depleted in 96 h. Moreover, the biomass reached a high concentration of 29 ± 1.2 g L
−1 in just 72 h and remained stable until the end of the experiment.
Shake-flask experiments revealed interesting differences in erythritol assimilation. The strains yl-hog1Δ and EUF1 yl-hog1Δ utilized glycerol and erythritol simultaneously. In contrast, the strain AJDU first produced erythritol, and the concentration of this polyol started to decrease only when glycerol was completely depleted. In cultures of euf1Δ yl-hog1Δ and eyd1Δ yl-hog1Δ the concentrations of erythritol remained stable. In cultures without erythritol supplementation of strains with yl-hog1 deletion, small amounts of this polyol (0.01–0.1 g L−1) were detected in the supernatant after 24 h of incubation until the end of the experiment.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}