The Mouse Cohesin-Associated Protein PDS5B Is Expressed in Testicular Cells and Is Associated with the Meiotic Chromosome Axes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
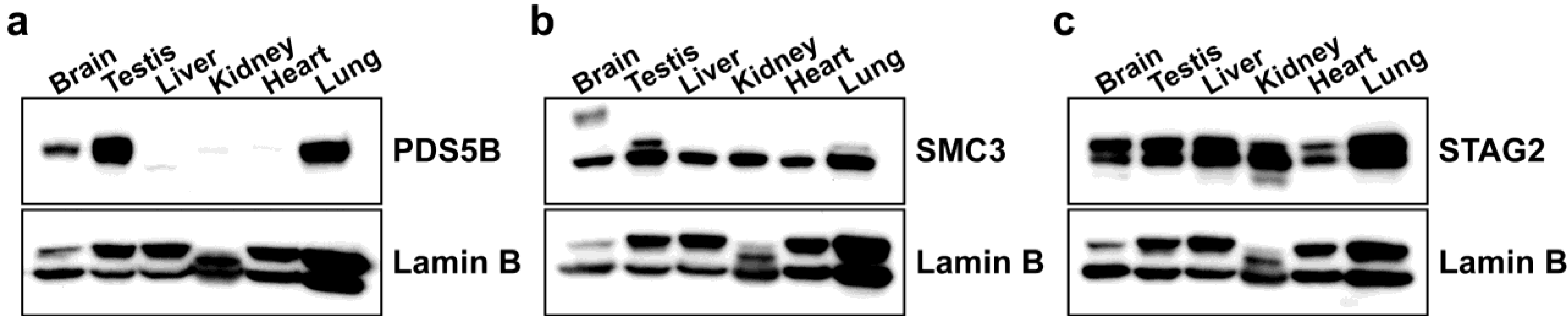
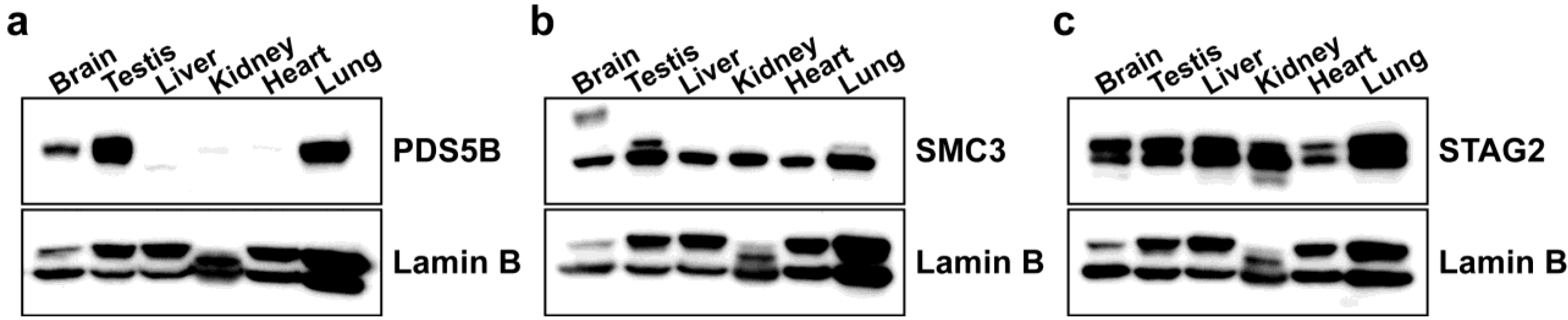
2.1. PDS5B Is Highly Expressed in Mouse Testis
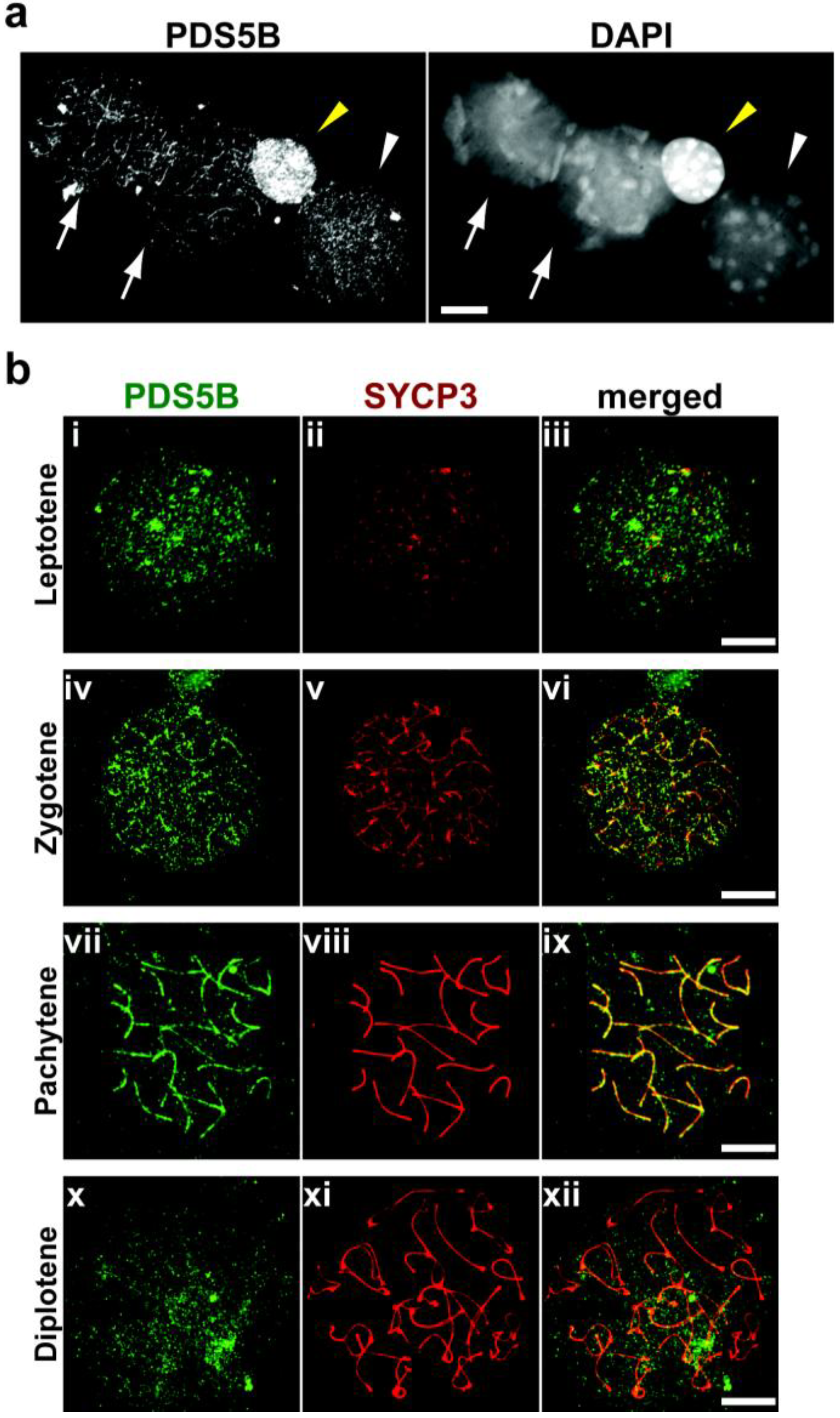
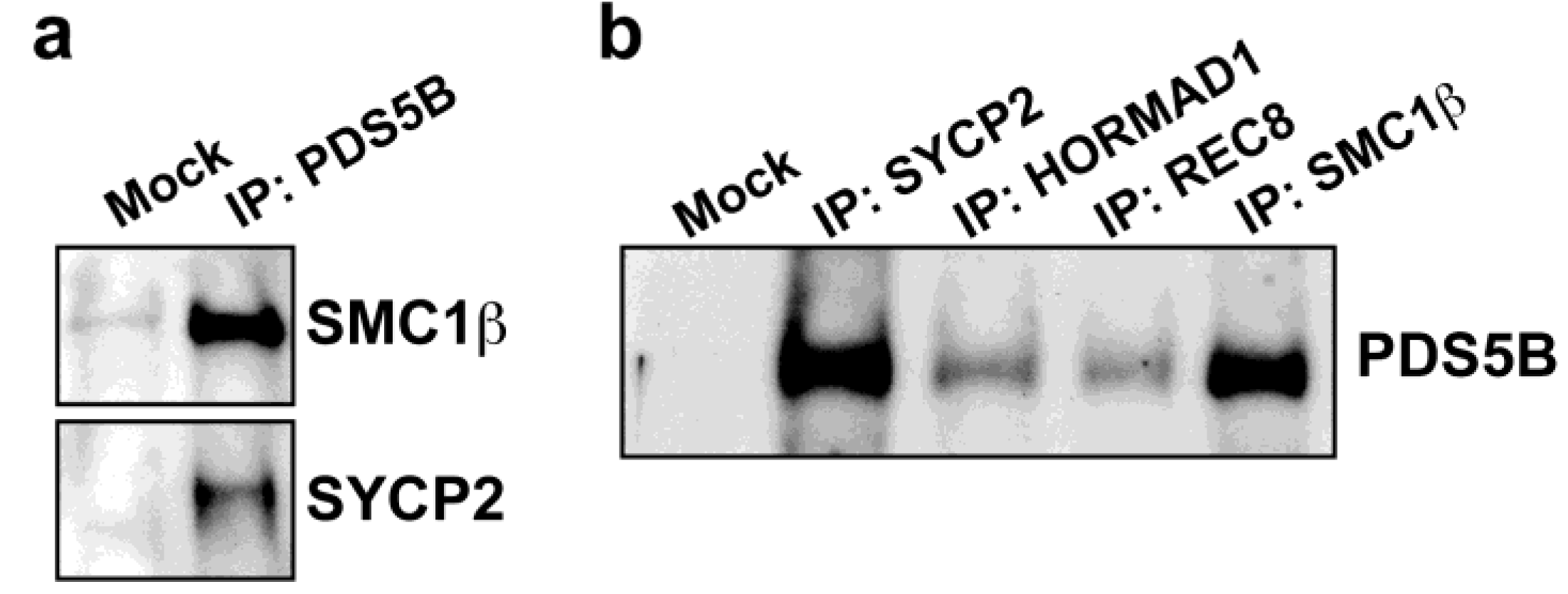
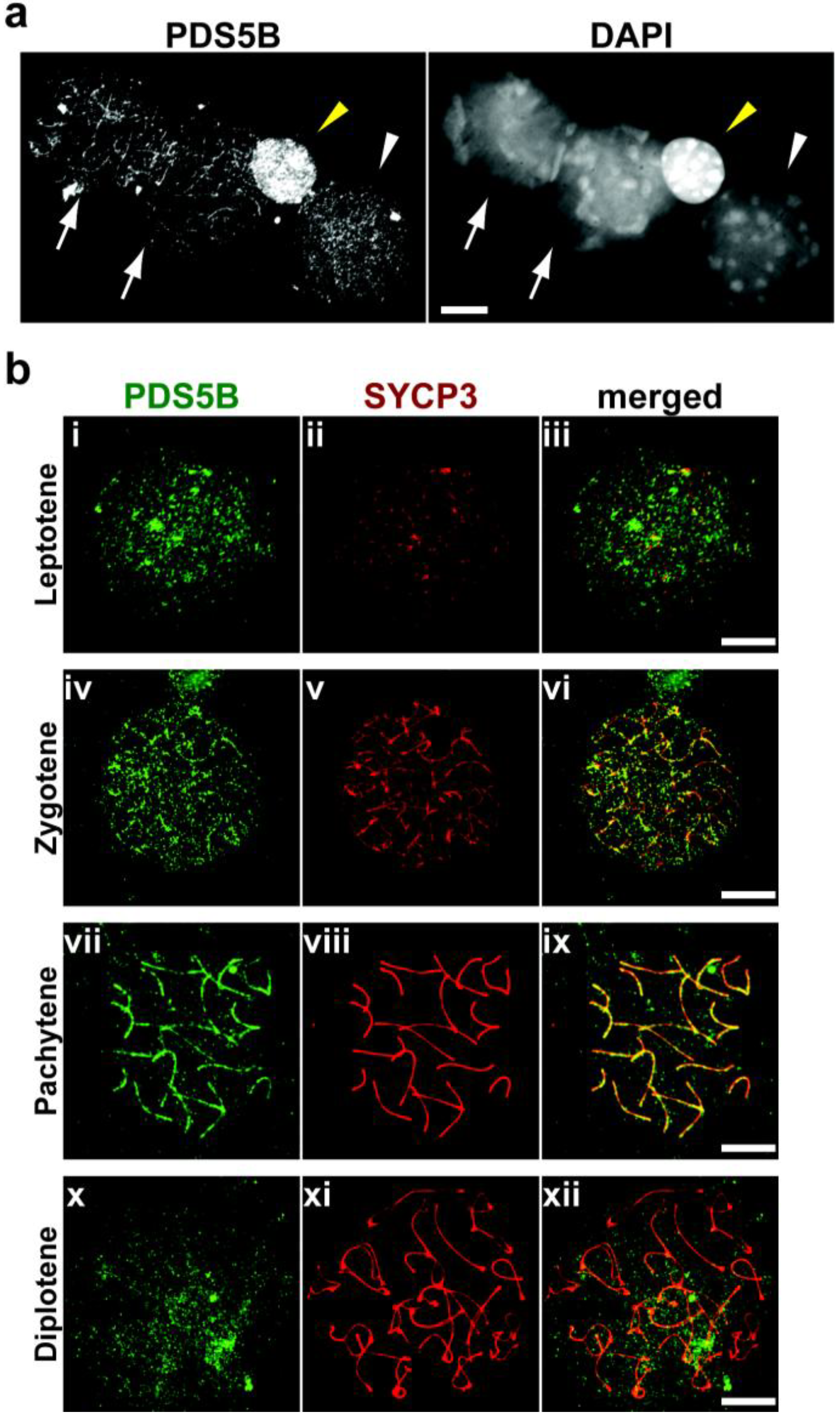
2.2. PDS5B Is Associated with the Meiotic Chromosome Axis
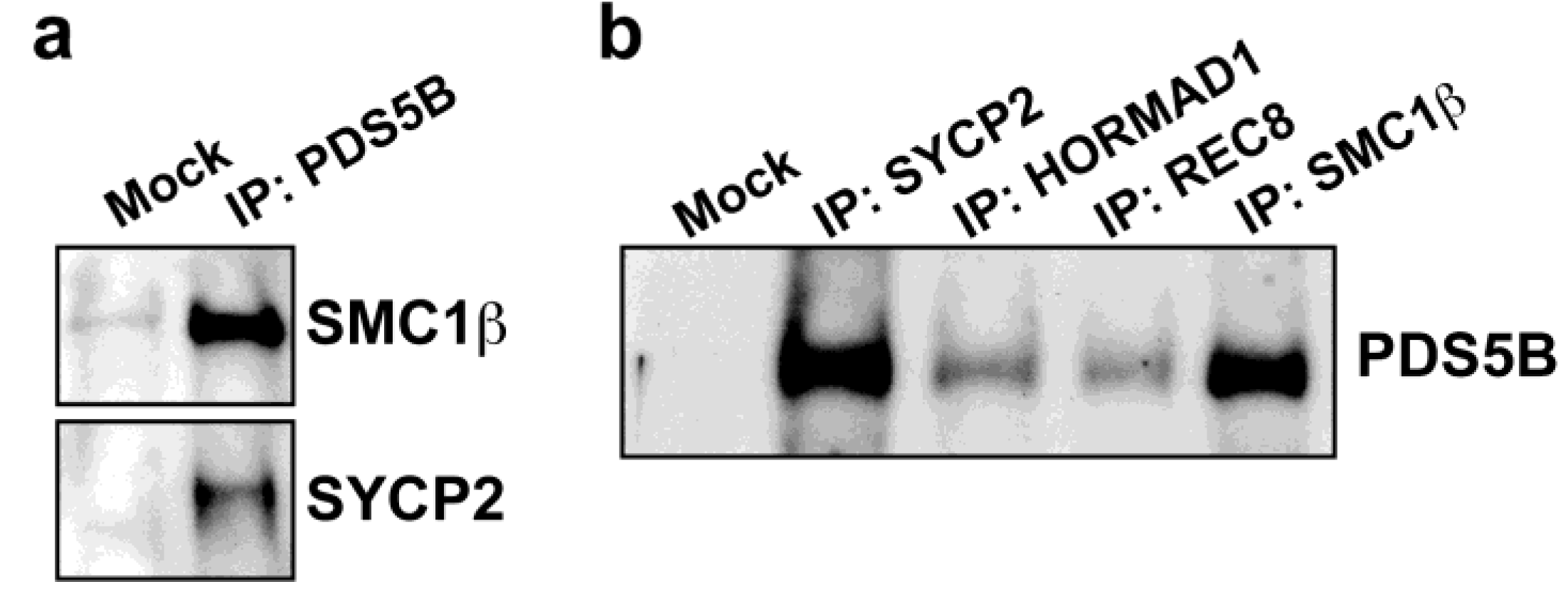
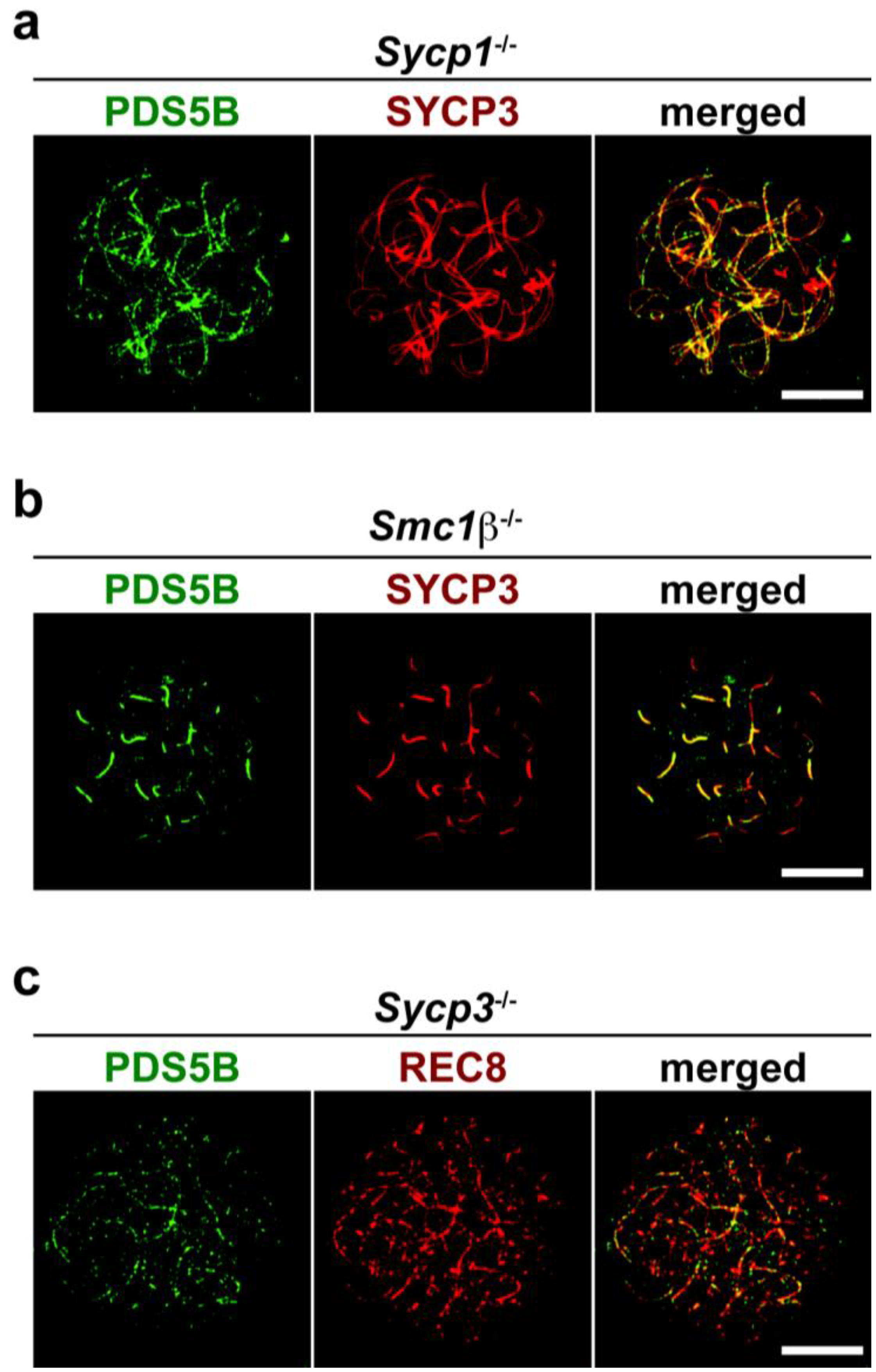
2.3. Axis Association of PDS5B Is Independent of SC Proteins
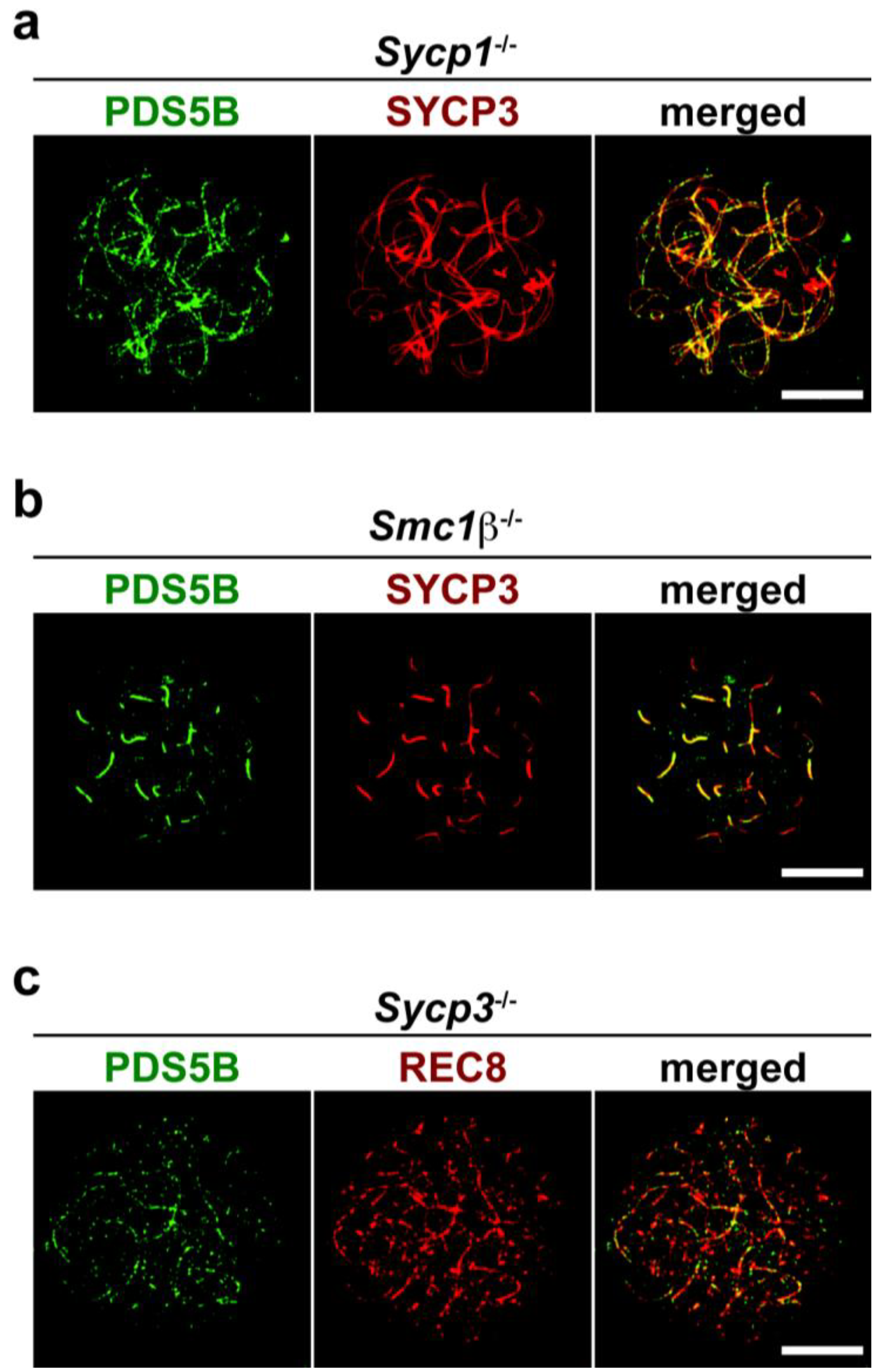
2.4. Possible Roles of the PDS5B Protein in Spermatogenesis
3. Experimental Section
3.1. Animals
3.2. Immunoblotting
3.3. Immunoprecipitation
3.4. Nuclear Spreads
4. Conclusions
Acknowledgements
Supplementary Files
References
- Petronczki, M.; Siomos, M.F.; Nasmyth, K. Un menage a quatre: The molecular biology of chromosome segregation in meiosis. Cell 2003, 112, 423–440. [Google Scholar] [CrossRef]
- Marston, A.L.; Amon, A. Meiosis: cell-cycle controls shuffle and deal. Nat. Rev. Mol. Cell Biol. 2004, 5, 983–997. [Google Scholar] [CrossRef]
- Keeney, S. Mechanism and control of meiotic recombination initiation. Curr. Top. Dev. Biol. 2001, 52, 1–53. [Google Scholar] [CrossRef]
- Bellani, M.A.; Romanienko, P.J.; Cairatti, D.A.; Camerini-Otero, R.D. SPO11 is required for sex-body formation, and Spo11 heterozygosity rescues the prophase arrest of Atm-/- spermatocytes. J. Cell Sci. 2005, 118, 3233–3245. [Google Scholar] [CrossRef]
- Zickler, D.; Kleckner, N. Meiotic chromosomes: integrating structure and function. Annu. Rev. Genet. 1999, 33, 603–754. [Google Scholar] [CrossRef]
- Page, S.L.; Hawley, R.S. The genetics and molecular biology of the synaptonemal complex. Annu. Rev. Cell Dev. Biol. 2004, 20, 525–558. [Google Scholar] [CrossRef]
- de Boer, E.; Heyting, C. The diverse roles of transverse filaments of synaptonemal complexes in meiosis. Chromosoma 2006, 115, 220–234. [Google Scholar] [CrossRef]
- Suja, J.A.; Barbero, J.L. Cohesin complexes and sister chromatid cohesion in mammalian meiosis. Genome Dyn. 2009, 5, 94–116. [Google Scholar] [CrossRef]
- Pelttari, J.; Hoja, M.R.; Yuan, L.; Liu, J.G.; Brundell, E.; Moens, P.; Santucci-Darmanin, S.; Jessberger, R.; Barbero, J.L.; Heyting, C.; Hoog, C. A meiotic chromosomal core consisting of cohesin complex proteins recruits DNA recombination proteins and promotes synapsis in the absence of an axial element in mammalian meiotic cells. Mol. Cell. Biol. 2001, 21, 5667–5677. [Google Scholar] [CrossRef]
- Bannister, L.A.; Reinholdt, L.G.; Munroe, R.J.; Schimenti, J.C. Positional cloning and characterization of mouse mei8, a disrupted allelle of the meiotic cohesin Rec8. Genesis 2004, 40, 184–194. [Google Scholar] [CrossRef]
- Revenkova, E.; Eijpe, M.; Heyting, C.; Hodges, C.A.; Hunt, P.A.; Liebe, B.; Scherthan, H.; Jessberger, R. Cohesin SMC1 beta is required for meiotic chromosome dynamics, sister chromatid cohesion and DNA recombination. Nat. Cell Biol. 2004, 6, 555–562. [Google Scholar] [CrossRef]
- Xu, H.; Beasley, M.D.; Warren, W.D.; van der Horst, G.T.; McKay, M.J. Absence of mouse REC8 cohesin promotes synapsis of sister chromatids in meiosis. Dev. Cell 2005, 8, 949–961. [Google Scholar] [CrossRef]
- Novak, I.; Wang, H.; Revenkova, E.; Jessberger, R.; Scherthan, H.; Hoog, C. Cohesin Smc1beta determines meiotic chromatin axis loop organization. J. Cell Biol. 2008, 180, 83–90. [Google Scholar] [CrossRef]
- Brar, G.A.; Hochwagen, A.; Ee, L.S.; Amon, A. The multiple roles of cohesin in meiotic chromosome morphogenesis and pairing. Mol. Biol. Cell 2009, 20, 1030–1047. [Google Scholar]
- Hartman, T.; Stead, K.; Koshland, D.; Guacci, V. Pds5p is an essential chromosomal protein required for both sister chromatid cohesion and condensation in Saccharomyces cerevisiae. J. Cell Biol. 2000, 151, 613–626. [Google Scholar] [CrossRef]
- Panizza, S.; Tanaka, T.; Hochwagen, A.; Eisenhaber, F.; Nasmyth, K. Pds5 cooperates with cohesin in maintaining sister chromatid cohesion. Curr. Biol. 2000, 10, 1557–1564. [Google Scholar] [CrossRef]
- Tanaka, K.; Hao, Z.; Kai, M.; Okayama, H. Establishment and maintenance of sister chromatid cohesion in fission yeast by a unique mechanism. EMBO J. 2001, 20, 5779–5790. [Google Scholar] [CrossRef]
- Losada, A.; Yokochi, T.; Hirano, T. Functional contribution of Pds5 to cohesin-mediated cohesion in human cells and Xenopus egg extracts. J. Cell Sci. 2005, 118, 2133–2141. [Google Scholar] [CrossRef]
- Shintomi, K.; Hirano, T. Releasing cohesin from chromosome arms in early mitosis: opposing actions of Wapl-Pds5 and Sgo1. Genes Dev. 2009, 23, 2224–2236. [Google Scholar] [CrossRef]
- van Heemst, D.; James, F.; Poggeler, S.; Berteaux-Lecellier, V.; Zickler, D. Spo76p is a conserved chromosome morphogenesis protein that links the mitotic and meiotic programs. Cell 1999, 98, 261–271. [Google Scholar] [CrossRef]
- Zhang, Z.; Ren, Q.; Yang, H.; Conrad, M.N.; Guacci, V.; Kateneva, A.; Dresser, M.E. Budding yeast PDS5 plays an important role in meiosis and is required for sister chromatid cohesion. Mol. Microbiol. 2005, 56, 670–680. [Google Scholar] [CrossRef]
- Ding, D.Q.; Sakurai, N.; Katou, Y.; Itoh, T.; Shirahige, K.; Haraguchi, T.; Hiraoka, Y. Meiotic cohesins modulate chromosome compaction during meiotic prophase in fission yeast. J. Cell Biol. 2006, 174, 499–508. [Google Scholar] [CrossRef]
- Jin, H.; Guacci, V.; Yu, H.G. Pds5 is required for homologue pairing and inhibits synapsis of sister chromatids during yeast meiosis. J. Cell Biol. 2009, 186, 713–725. [Google Scholar] [CrossRef]
- Wang, F.; Yoder, J.; Antoshechkin, I.; Han, M. Caenorhabditis elegans EVL-14/PDS-5 and SCC-3 are essential for sister chromatid cohesion in meiosis and mitosis. Mol. Cell. Biol. 2003, 23, 7698–7707. [Google Scholar] [CrossRef]
- Zhang, B.; Jain, S.; Song, H.; Fu, M.; Heuckeroth, R.O.; Erlich, J.M.; Jay, P.Y.; Milbrandt, J. Mice lacking sister chromatid cohesion protein PDS5B exhibit developmental abnormalities reminiscent of Cornelia de Lange syndrome. Development 2007, 134, 3191–3201. [Google Scholar] [CrossRef]
- Zhang, B.; Chang, J.; Fu, M.; Huang, J.; Kashyap, R.; Salavaggione, E.; Jain, S.; Kulkarni, S.; Deardorff, M.A.; Uzielli, M.L.; Dorsett, D.; Beebe, D.C.; Jay, P.Y.; Heuckeroth, R.O.; Krantz, I.; Milbrandt, J. Dosage effects of cohesin regulatory factor PDS5 on mammalian development: implications for cohesinopathies. PLoS One 2009, 4, e5232. [Google Scholar] [CrossRef]
- de Vries, F.A.; de Boer, E.; van den Bosch, M.; Baarends, W.M.; Ooms, M.; Yuan, L.; Liu, J.G.; van Zeeland, A.A.; Heyting, C.; Pastink, A. Mouse Sycp1 functions in synaptonemal complex assembly, meiotic recombination, and XY body formation. Genes Dev. 2005, 19, 1376–1389. [Google Scholar] [CrossRef]
- Gandhi, R.; Gillespie, P.J.; Hirano, T. Human Wapl is a cohesin-binding protein that promotes sister-chromatid resolution in mitotic prophase. Curr. Biol. 2006, 16, 2406–2417. [Google Scholar] [CrossRef]
- Kueng, S.; Hegemann, B.; Peters, B.H.; Lipp, J.J.; Schleiffer, A.; Mechtler, K.; Peters, J.M. Wapl controls the dynamic association of cohesin with chromatin. Cell 2006, 127, 955–967. [Google Scholar] [CrossRef]
- Rowland, B.D.; Roig, M.B.; Nishino, T.; Kurze, A.; Uluocak, P.; Mishra, A.; Beckouet, F.; Underwood, P.; Metson, J.; Imre, R.; Mechtler, K.; Katis, V.L.; Nasmyth, K. Building sister chromatid cohesion: Smc3 acetylation counteracts an antiestablishment activity. Mol. Cell 2009, 33, 763–774. [Google Scholar] [CrossRef]
- Sutani, T.; Kawaguchi, T.; Kanno, R.; Itoh, T.; Shirahige, K. Budding yeast Wpl1(Rad61)-Pds5 complex counteracts sister chromatid cohesion-establishing reaction. Curr. Biol. 2009, 19, 492–497. [Google Scholar] [CrossRef]
- Kuroda, M.; Oikawa, K.; Ohbayashi, T.; Yoshida, K.; Yamada, K.; Mimura, J.; Matsuda, Y.; Fujii-Kuriyama, Y.; Mukai, K. A dioxin sensitive gene, mammalian WAPL, is implicated in spermatogenesis. FEBS Lett. 2005, 579, 167–172. [Google Scholar] [CrossRef]
- Zhang, J.; Hakansson, H.; Kuroda, M.; Yuan, L. Wapl localization on the synaptonemal complex, a meiosis-specific proteinaceous structure that binds homologous chromosomes, in the female mouse. Reprod. Domest. Anim. 2008, 43, 124–126. [Google Scholar]
- Kitajima, T.S.; Sakuno, T.; Ishiguro, K.; Iemura, S.; Natsume, T.; Kawashima, S.A.; Watanabe, Y. Shugoshin collaborates with protein phosphatase 2A to protect cohesin. Nature 2006, 441, 46–52. [Google Scholar] [CrossRef]
- Lee, J.; Kitajima, T.S.; Tanno, Y.; Yoshida, K.; Morita, T.; Miyano, T.; Miyake, M.; Watanabe, Y. Unified mode of centromeric protection by shugoshin in mammalian oocytes and somatic cells. Nat. Cell Biol. 2008, 10, 42–52. [Google Scholar] [CrossRef]
- Llano, E.; Gomez, R.; Gutierrez-Caballero, C.; Herran, Y.; Sanchez-Martin, M.; Vazquez-Quinones, L.; Hernandez, T.; de Alava, E.; Cuadrado, A.; Barbero, J.L.; Suja, J.A.; Pendas, A.M. Shugoshin-2 is essential for the completion of meiosis but not for mitotic cell division in mice. Genes Dev. 2008, 22, 2400–2413. [Google Scholar] [CrossRef]
- Yuan, L.; Liu, J.G.; Zhao, J.; Brundell, E.; Daneholt, B.; Hoog, C. The murine SCP3 gene is required for synaptonemal complex assembly, chromosome synapsis, and male fertility. Mol. Cell 2000, 5, 73–83. [Google Scholar] [CrossRef]
- Fukuda, T.; Daniel, K.; Wojtasz, L.; Toth, A.; Hoog, C. A novel mammalian HORMA domain-containing protein, HORMAD1, preferentially associates with unsynapsed meiotic chromosomes. Exp. Cell Res. 2010, 316, 158–171. [Google Scholar] [CrossRef]
- Djabali, K.; Portier, M.M.; Gros, F.; Blobel, G.; Georgatos, S.D. Network antibodies identify nuclear lamin B as a physiological attachment site for peripherin intermediate filaments. Cell 1991, 64, 109–121. [Google Scholar] [CrossRef]
- Kouznetsova, A.; Novak, I.; Jessberger, R.; Hoog, C. SYCP2 and SYCP3 are required for cohesin core integrity at diplotene but not for centromere cohesion at the first meiotic division. J. Cell Sci. 2005, 118, 2271–2278. [Google Scholar] [CrossRef]
- Peters, A.H.; Plug, A.W.; van Vugt, M.J.; de Boer, P. A drying-down technique for the spreading of mammalian meiocytes from the male and female germline. Chromosome Res. 1997, 5, 66–68. [Google Scholar]
© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Fukuda, T.; Hoog, C. The Mouse Cohesin-Associated Protein PDS5B Is Expressed in Testicular Cells and Is Associated with the Meiotic Chromosome Axes. Genes 2010, 1, 484-494. https://doi.org/10.3390/genes1030484
Fukuda T, Hoog C. The Mouse Cohesin-Associated Protein PDS5B Is Expressed in Testicular Cells and Is Associated with the Meiotic Chromosome Axes. Genes. 2010; 1(3):484-494. https://doi.org/10.3390/genes1030484
Chicago/Turabian StyleFukuda, Tomoyuki, and Christer Hoog. 2010. "The Mouse Cohesin-Associated Protein PDS5B Is Expressed in Testicular Cells and Is Associated with the Meiotic Chromosome Axes" Genes 1, no. 3: 484-494. https://doi.org/10.3390/genes1030484
APA StyleFukuda, T., & Hoog, C. (2010). The Mouse Cohesin-Associated Protein PDS5B Is Expressed in Testicular Cells and Is Associated with the Meiotic Chromosome Axes. Genes, 1(3), 484-494. https://doi.org/10.3390/genes1030484