Comparative Characterization of Oil Body Proteins from Hemp, Plum, and Jujube Seed and Their Application in Curcumin-Loaded Artificial Oleosomes


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Extraction of Oleosomes from Hemp, Plum, and Jujube Seed
2.3. Isolation of OBPs from Oleosomes
2.4. Physicochemical Properties of OBPs
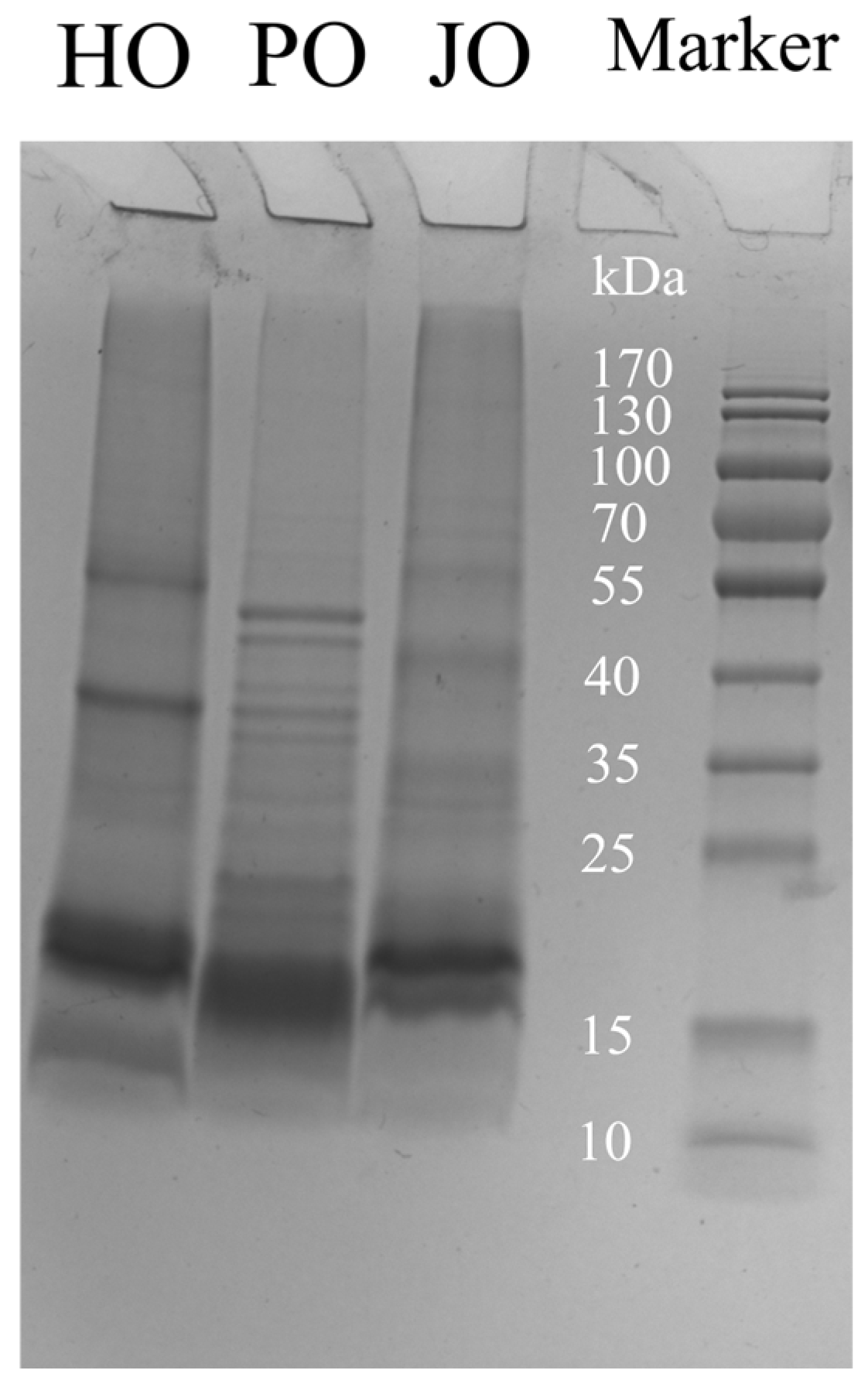
2.4.1. Analysis of Subunit Composition
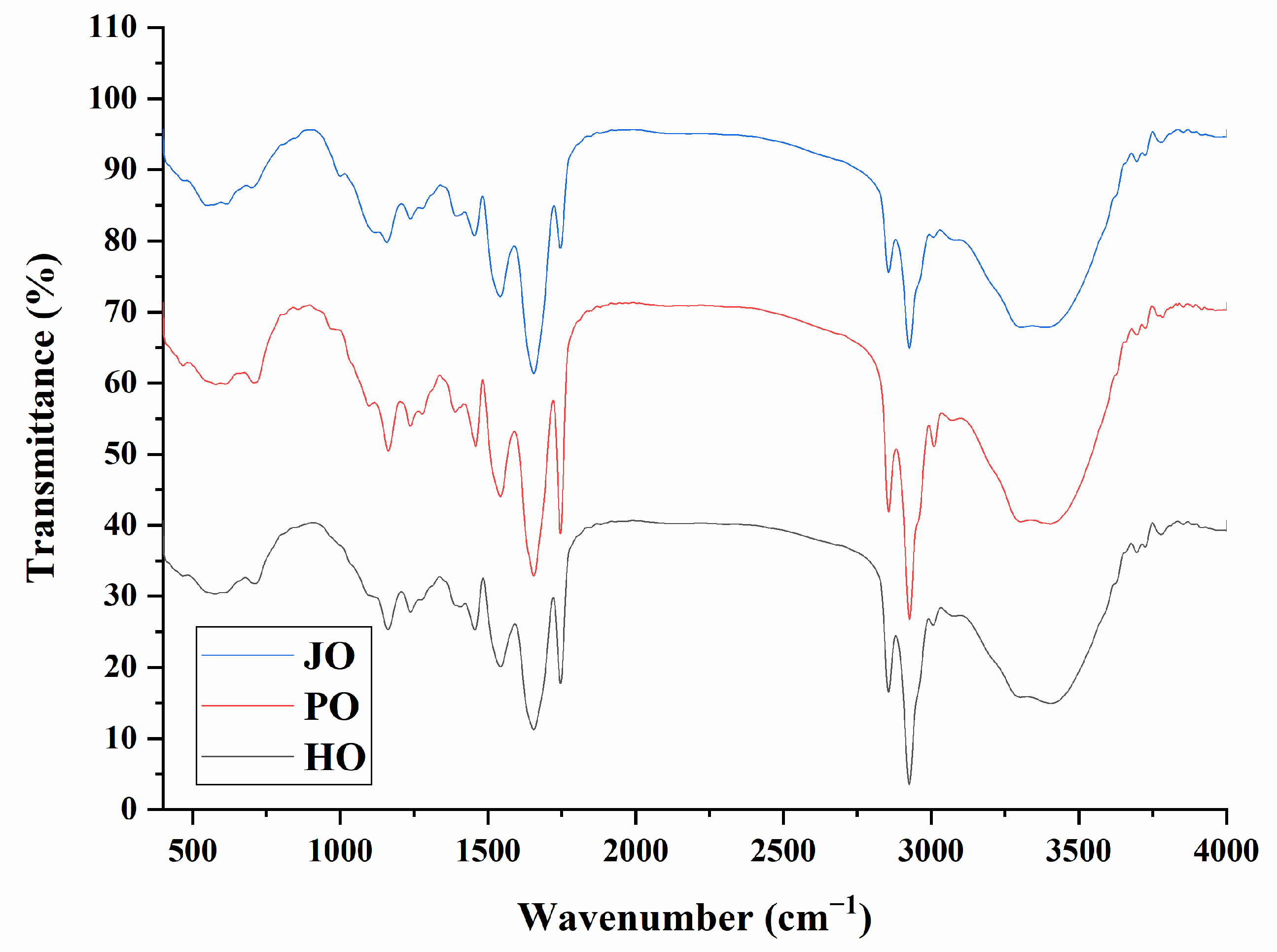
2.4.2. FTIR Spectroscopy for Secondary Structure Analysis
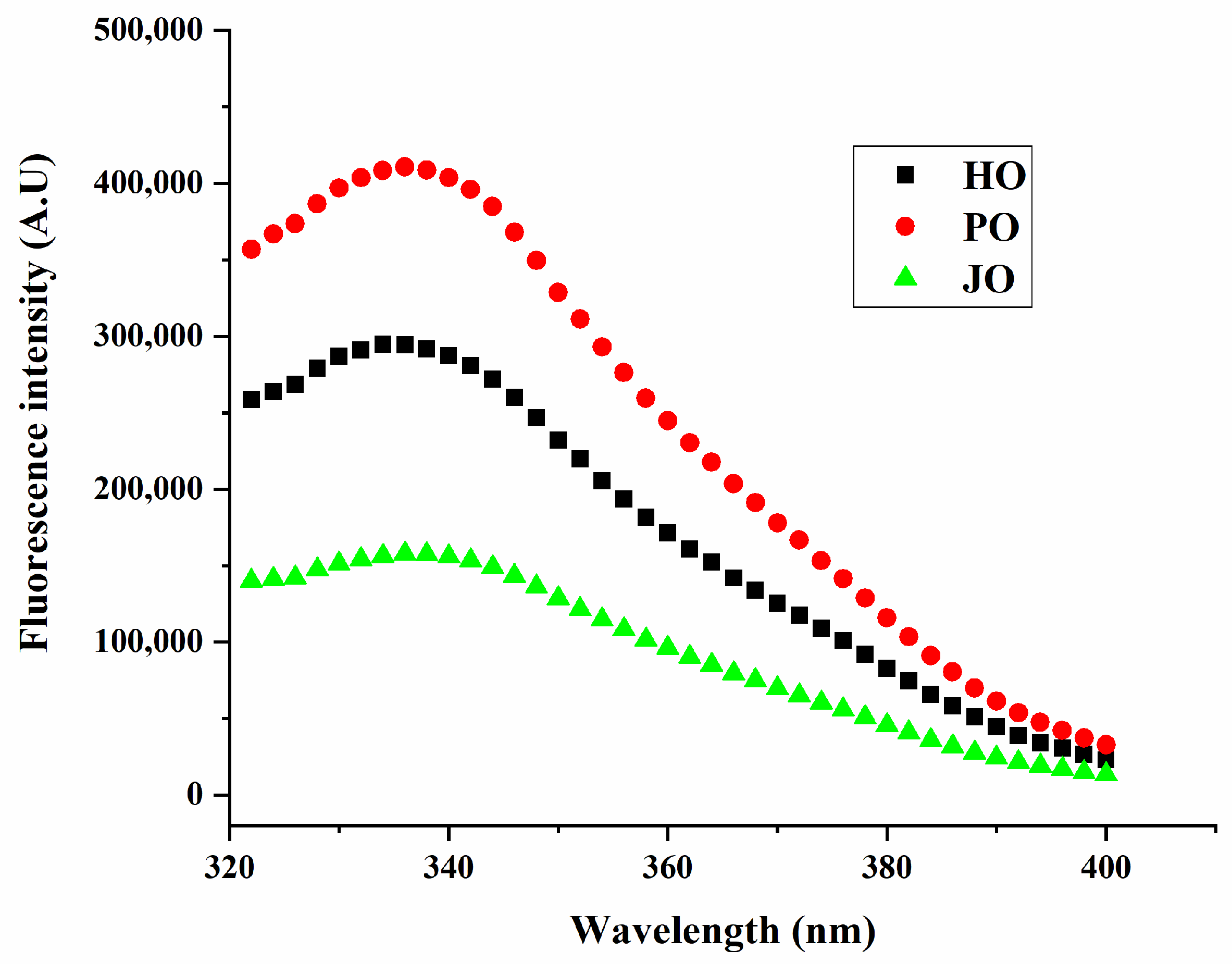
2.4.3. Intrinsic Fluorescence Spectroscopy for Tertiary Structure
2.4.4. Particle Size and Zeta Potential of OBPs
2.4.5. Microstructure of OBPs
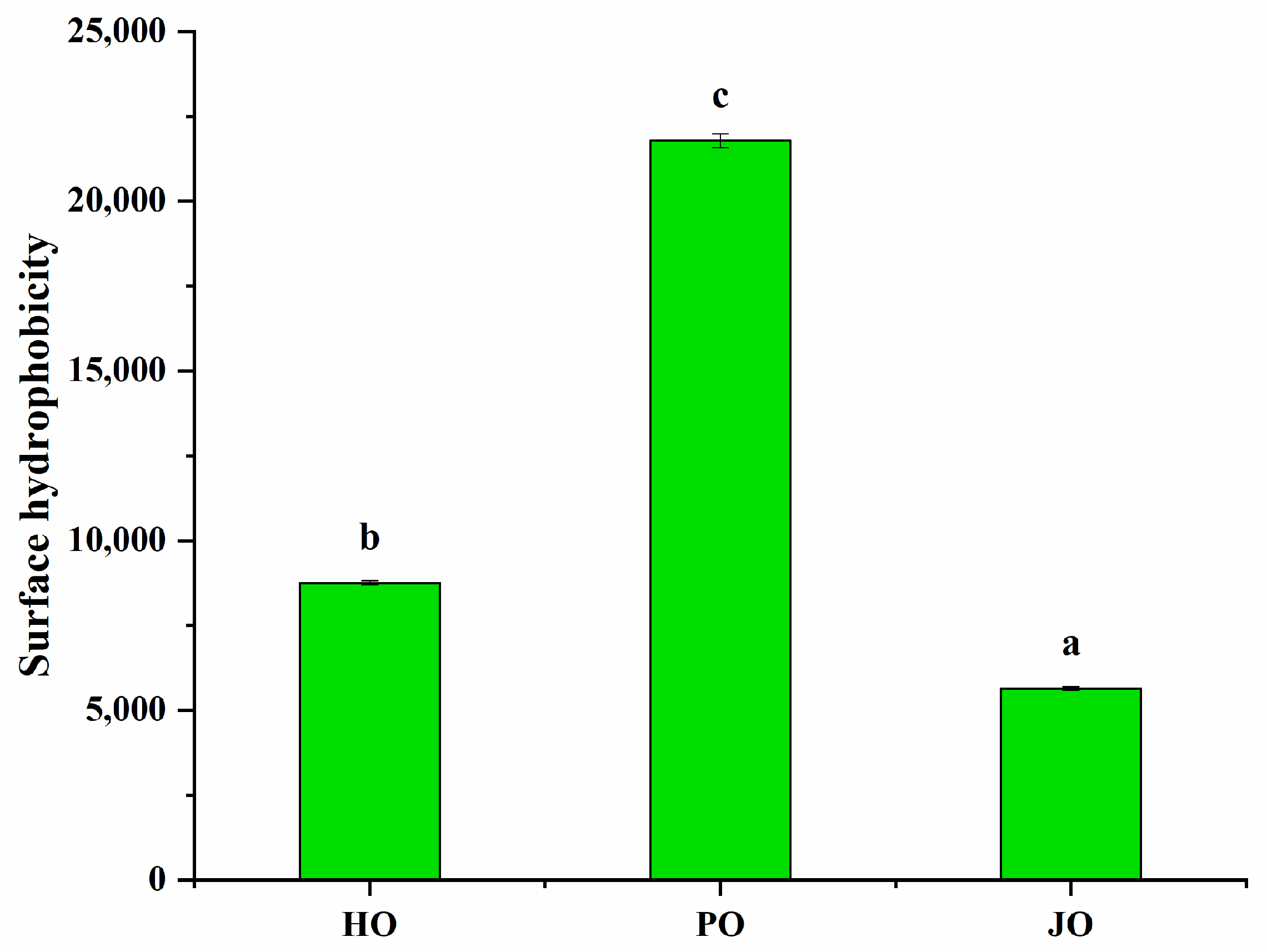
2.4.6. Surface Hydrophobicity of OBPs
2.4.7. Solubility of OBPs
2.4.8. Thermal Stability of OBPs
2.4.9. Emulsifying Properties of OBPs
2.5. Preparation of Artificial Oleosomes (AOs) Loaded with Curcumin
2.6. Physicochemical Properties of AOs Loaded with Curcumin
2.6.1. Encapsulation Efficiency
2.6.2. Particle Size and Zeta Potential
2.6.3. Microstructure
2.6.4. Free Radical Scavenging Activity
2.6.5. Bio-Accessibility of Curcumin
2.6.6. Storage, Temperature, and Salt Stability
2.7. Statistical Analysis
3. Results and Discussion
3.1. Structure of OBPs
3.1.1. Subunits
3.1.2. FTIR Spectrum
3.1.3. Secondary Structures
3.1.4. Intrinsic Fluorescence Emission Spectra
3.2. Physicochemical Properties of OBPs
3.2.1. Particle Size and Zeta Potential
3.2.2. Microstructure
3.2.3. Surface Hydrophobicity
3.2.4. Solubility
3.2.5. Thermal Stability
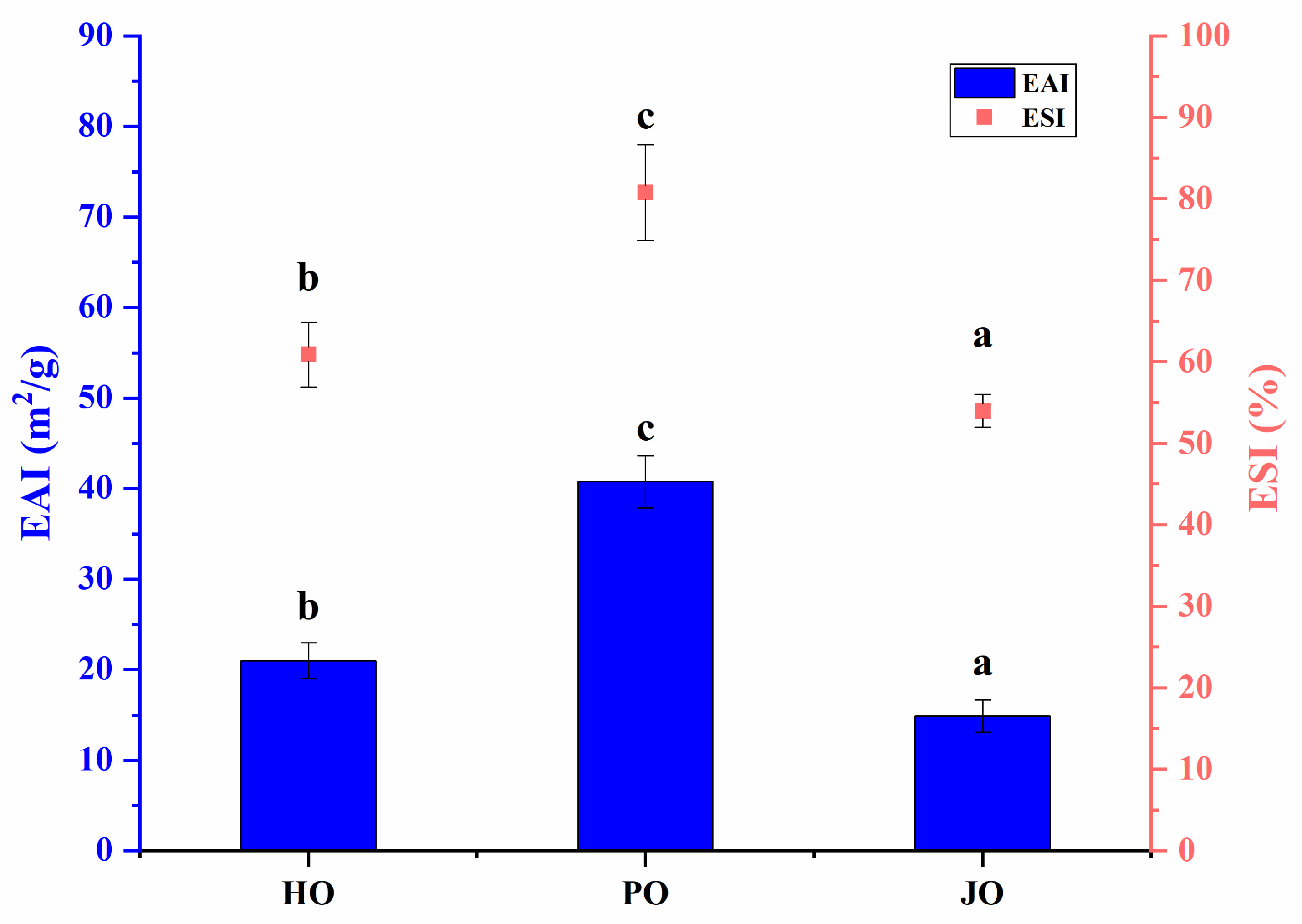
3.2.6. Emulsifying Properties
3.3. Physicochemical Properties of AOs Loaded with Curcumin
3.3.1. Appearance
3.3.2. Encapsulation Efficiency
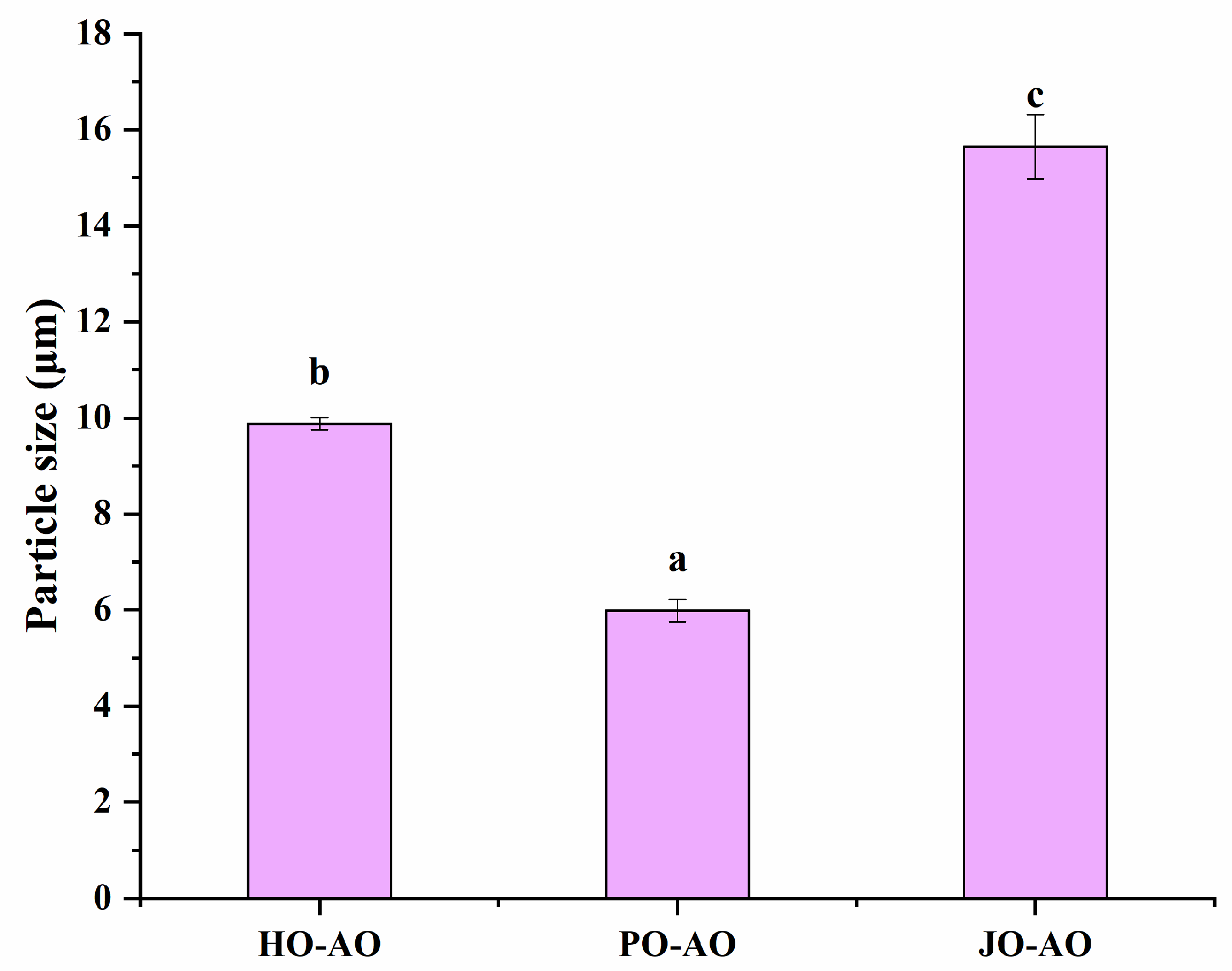
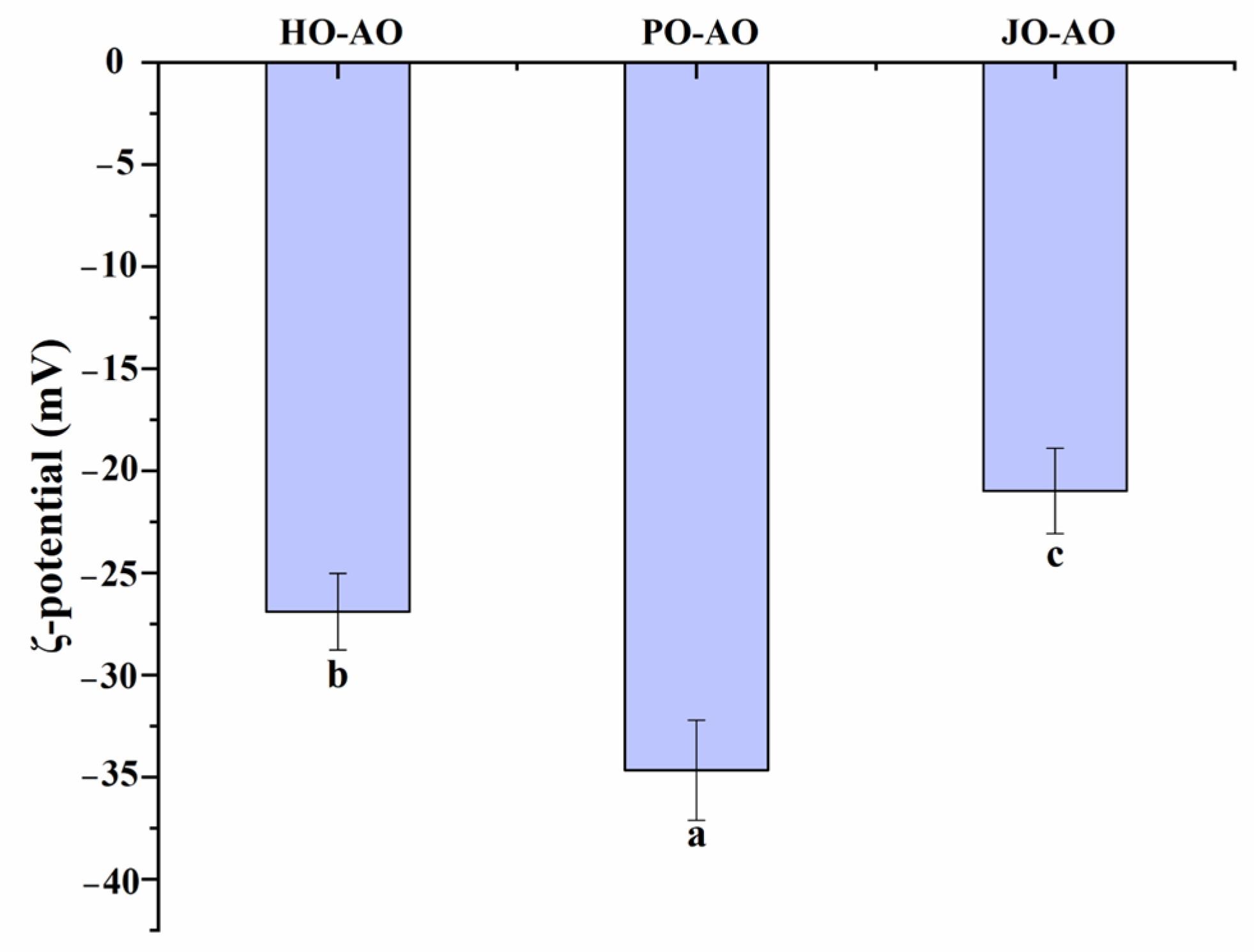
3.3.3. Particle Size and Zeta Potential
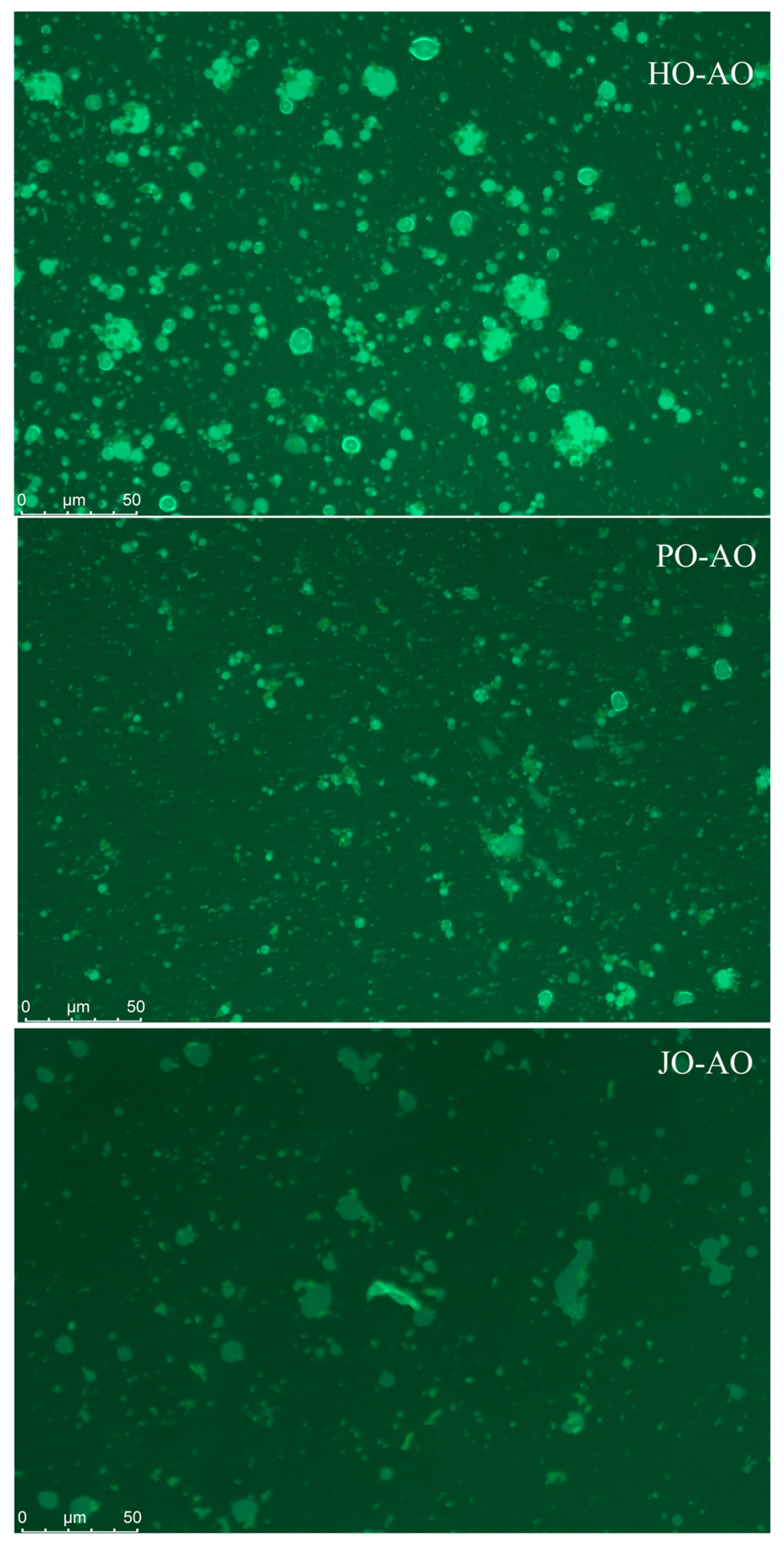
3.3.4. Microstructure
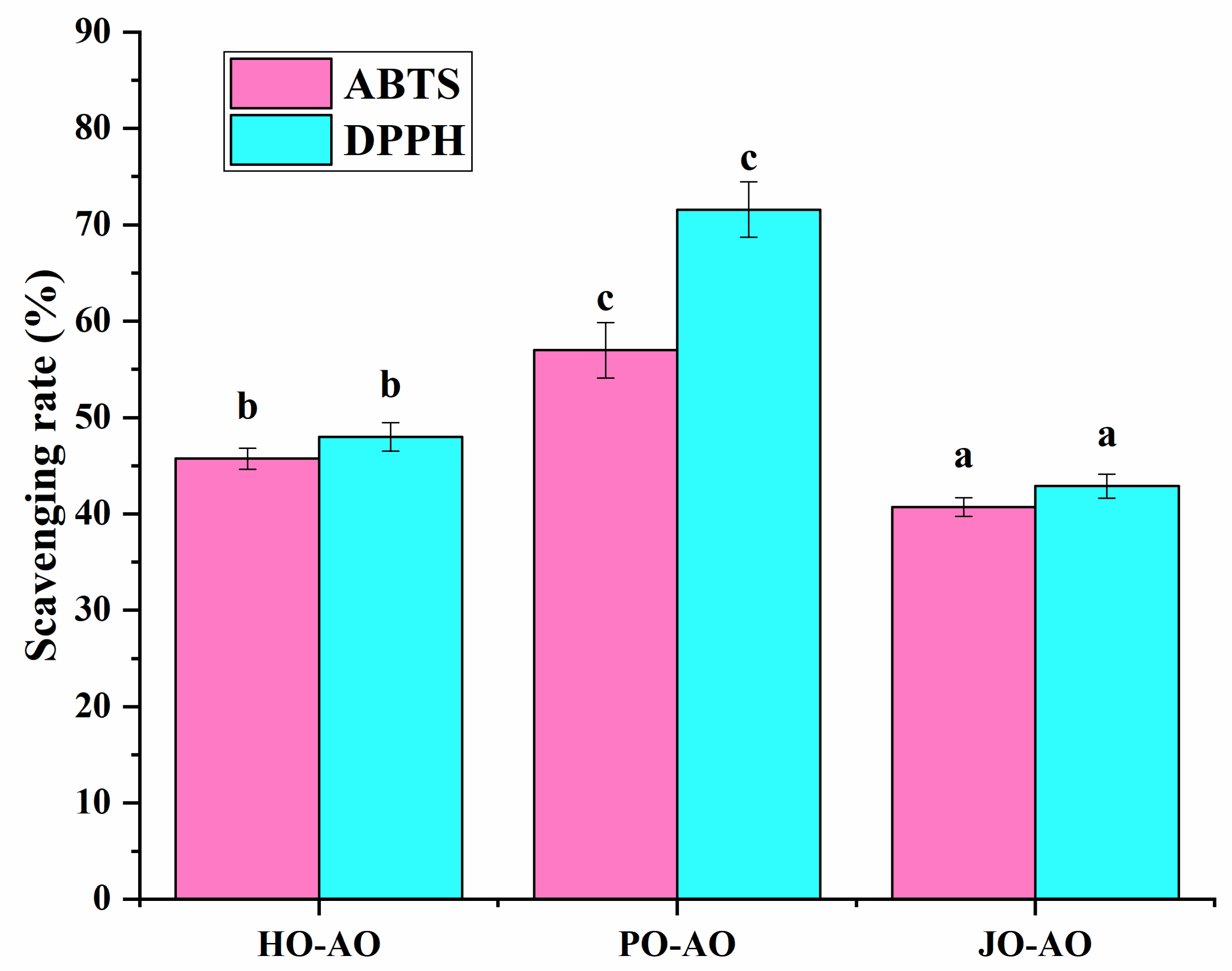
3.3.5. Free Radical Scavenging Activity
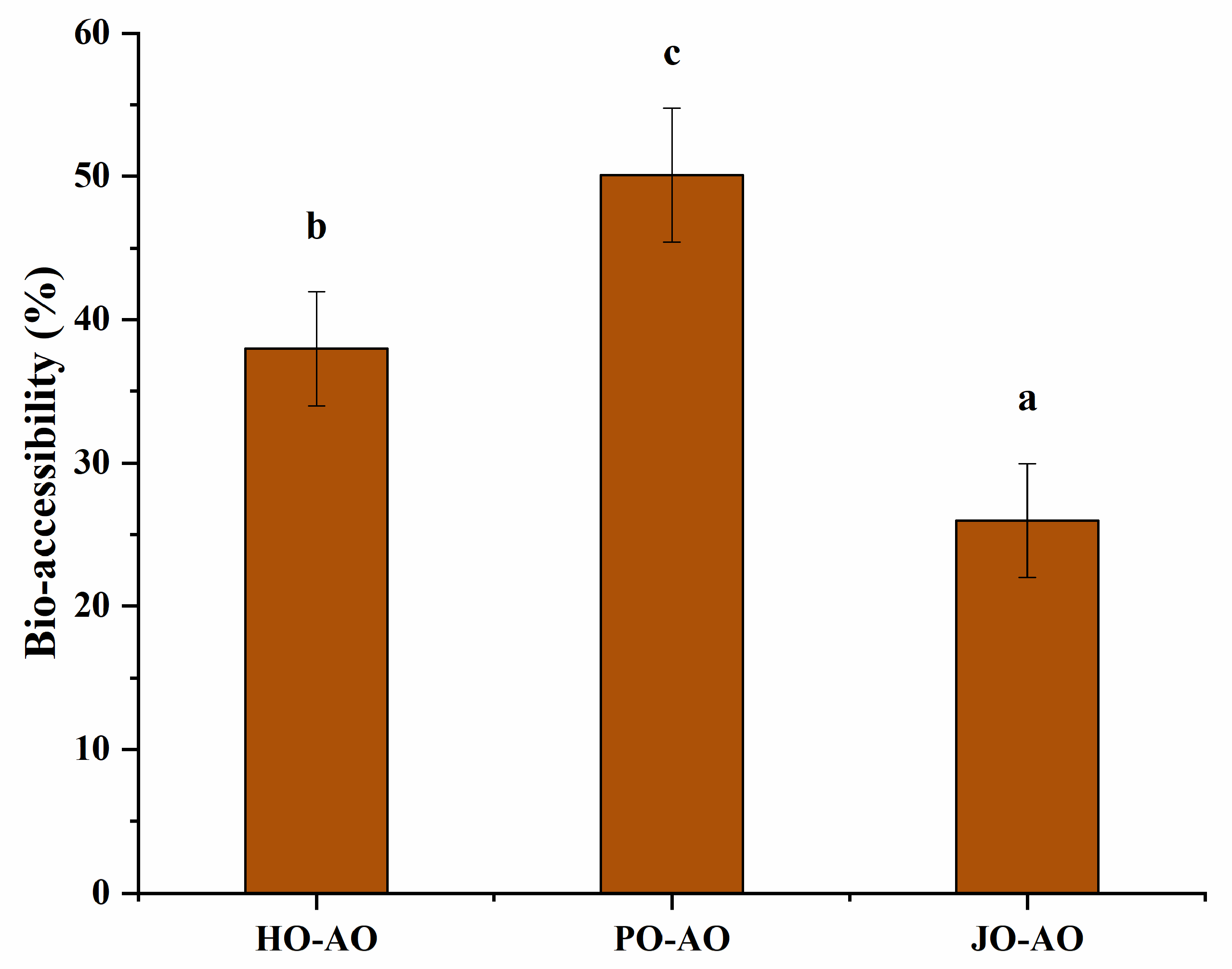
3.3.6. Bio-Accessibility of Curcumin
3.3.7. Storage, Temperature, and Salt Stability
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yuan, R.; Liu, J.; Ukwatta, R.H.; Xue, F.; Xiong, X.; Li, C. Artificial oil bodies: A review on composition, properties, biotechnological applications, and improvement methods. Food Chem. X 2024, 21, 101109. [Google Scholar] [CrossRef] [PubMed]
- Abdullah; Weiss, J.; Zhang, H. Recent advances in the composition, extraction and food applications of plant-derived oleosomes. Trends Food Sci. Technol. 2020, 106, 322–332. [Google Scholar] [CrossRef]
- Yuan, R.; Yi, M.; Xue, F.; Xiong, X.; Adhikari, B.; Li, C. Comparative analysis of physicochemical properties and functionalities of artificial oil bodies stabilized by different seed oil body proteins. Int. J. Biol. Macromol. 2025, 307, 141893. [Google Scholar] [CrossRef]
- Cao, Y.; Zhao, L.; Ying, Y.; Kong, X.; Hua, Y.; Chen, Y. The characterization of soybean oil body integral oleosin isoforms and the effects of alkaline pH on them. Food Chem. 2015, 177, 288–294. [Google Scholar] [CrossRef]
- Deleu, M.; Vaca-Medina, G.; Fabre, J.-F.; Roïz, J.; Valentin, R.; Mouloungui, Z. Interfacial properties of oleosins and phospholipids from rapeseed for the stability of oil bodies in aqueous medium. Colloids Surf. B Biointerfaces 2010, 80, 125–132. [Google Scholar] [CrossRef]
- Macias, M.L.; Gómez, F.; Diaz, A.; Mayorga, C.; Aranda, A.; González, M.; Blanca-López, N.; Canto, G.; Osorio, M.; Garrido, M.; et al. Basophil Response To Storage Proteins and Oleosins From Sunflower Seed. J. Allergy Clin. Immunol. 2014, 133, AB111. [Google Scholar] [CrossRef]
- Khongrum, J.; Mairuae, N.; Thanchomnang, T.; Zhang, M.; Bai, G.; Palachai, N. Synergistic Neuroprotection Through Epigenetic Modulation by Combined Curcumin-Enriched Turmeric Extract and L-Ascorbic Acid in Oxidative Stress-Induced SH-SY5Y Cell Damage. Foods 2025, 14, 892. [Google Scholar] [CrossRef]
- Xiao, H.; Wang, L.; Bu, N.; Duan, J.; Pang, J. Electrospun Photodynamic Antibacterial Konjac Glucomannan/Polyvinylpyrrolidone Nanofibers Incorporated with Lignin-Zinc Oxide Nanoparticles and Curcumin for Food Packaging. Foods 2024, 13, 2007. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, H.; Feng, Z.; Lu, Z.; Su, C.; Zhao, Y.; Yu, J.; Dushkin, A.V.; Su, W. Preparation of pectin-tannic acid coated core-shell nanoparticle for enhanced bioavailability and antihyperlipidemic activity of curcumin. Food Hydrocoll. 2021, 119, 106858. [Google Scholar] [CrossRef]
- Bhinge, S.D.; Nayakal, P.D.; Bhutkar, M.A.; Jagtap, A.; Gade, V.; Jadhav, N.R.; Jadhav, S.T. Synergistic anti-inflammatory effects of curcumin and ginger oil: A potential alternative to steroid therapy. S. Afr. J. Bot. 2025, 180, 369–377. [Google Scholar] [CrossRef]
- Rahman, M.; Farooq, S. Role of peanut oleosomes in the delivery of curcumin embedded in interpenetrating emulsion-filled gels made with whey protein and chitosan. Colloids Surf. A Physicochem. Eng. Asp. 2025, 707, 135962. [Google Scholar] [CrossRef]
- Liu, X.; Shi, Y.; Wang, M.; Adhikari, B.; Xue, F. Covalent conjugation of hemp protein isolates with curcumin via ultrasound to improve its structural and functional properties. Food Chem. 2025, 482, 144096. [Google Scholar] [CrossRef] [PubMed]
- Xue, F.; Li, C.; Zhu, X.; Wang, L.; Pan, S. Comparative studies on the physicochemical properties of soy protein isolate-maltodextrin and soy protein isolate-gum acacia conjugate prepared through Maillard reaction. Food Res. Int. 2013, 51, 490–495. [Google Scholar] [CrossRef]
- Hu, M.; Du, X.; Liu, G.; Tan, Z.; Zhang, S.; Qi, B.; Li, Y. Investigation of structure–stability correlations of reconstructed oil bodies. LWT 2022, 165, 113740. [Google Scholar] [CrossRef]
- Song, Y.; Su, S.; Yang, T.; Li, B.; Li, L.; Zhang, X. Enhanced bioaccessibility of curcumin in Pickering emulsions stabilized by solid lipid particles. LWT 2023, 188, 115481. [Google Scholar] [CrossRef]
- Liu, Y.; Fei, Y.; Li, C.; Cheng, J.; Xue, F. Impact of Probiotic Fermentation on the Physicochemical Properties of Hemp Seed Protein Gels. Polymers 2024, 16, 3032. [Google Scholar] [CrossRef]
- Liu, X.; Xue, F.; Adhikari, B. Production of hemp protein isolate-polyphenol conjugates through ultrasound and alkali treatment methods and their characterization. Future Foods 2023, 7, 100210. [Google Scholar] [CrossRef]
- Sun, Y.; Zhong, M.; Liao, Y.; Kang, M.; Li, Y.; Qi, B. Interfacial characteristics of artificial oil body emulsions (O/W) prepared using extrinsic and intrinsic proteins: Inspired by natural oil body. LWT 2023, 173, 114270. [Google Scholar] [CrossRef]
- Liu, X.; Xue, F.; Adhikari, B. Hemp protein isolate-polysaccharide complex coacervates and their application as emulsifiers in oil-in-water emulsions. Food Hydrocoll. 2023, 137, 108352. [Google Scholar] [CrossRef]
- Franzoi, M.; Costa, A.; Vigolo, V.; Penasa, M.; De Marchi, M. Effect of pasteurization on coagulation properties of bovine milk and the role of major composition traits and protein fractions. J. Food Compos. Anal. 2022, 114, 104808. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, S.; Jiang, W.; Zhao, H.; Fu, J. Ability of casein hydrolysate-carboxymethyl chitosan conjugates to stabilize a nanoemulsion: Improved freeze-thaw and pH stability. Food Hydrocoll. 2020, 101, 105452. [Google Scholar] [CrossRef]
- Liao, Y.; Wang, Z.; Pei, Y.; Yan, S.; Chen, T.; Qi, B.; Li, Y. Unveiling the applications of membrane proteins from oil bodies: Leading the way in artificial oil body technology and other biotechnological advancements. Crit. Rev. Food Sci. Nutr. 2025, 65, 2295–2322. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Gao, Y.; Pei, Y.; Ma, L.; Yan, S.; Qi, B.; Li, Y. Characterization of the soybean oil body interface: Revealing the mechanism of changes in the interfacial protein composition, structure, and function of oil bodies extracted under different denaturation conditions. Food Hydrocoll. 2025, 162, 110930. [Google Scholar] [CrossRef]
- Farooq, S.; Ahmad, M.I.; Zheng, S.; Ali, U.; Abdullah; Xue, J.; Li, Y.; Shixiu, C.; Zhang, H. Fabrication and characterization of double-network meat analogs based on camellia oleosomes and soy protein-wheat gluten induced by transglutaminase and calcium alginate. Int. J. Biol. Macromol. 2025, 310, 143322. [Google Scholar] [CrossRef]
- Peng, Y.; Shan, Z.; Jia, W.; Li, M.; Wen, X.; Ni, Y. Comparative analysis of twin-screw pressing and blending methods for walnut oleosome extraction: Yield, physical stability, and functionalities. J. Food Eng. 2025, 386, 112292. [Google Scholar] [CrossRef]
- Di, D.; Zhang, R.; Li, H.; Qiu, M.; Jiang, S.; Luo, S.; Sun, H.; He, S. Insights from structural characterization for understanding isolation and impact of roasting on allergenic potential of sesame Oleosins. Food Chem. 2025, 481, 144095. [Google Scholar] [CrossRef]
- Dou, L.; Li, B.; Zhang, K.; Chu, X.; Hou, H. Physical properties and antioxidant activity of gelatin-sodium alginate edible films with tea polyphenols. Int. J. Biol. Macromol. 2018, 118, 1377–1383. [Google Scholar] [CrossRef]
- Xue, F.; Li, C.; Adhikari, B. Physicochemical properties of soy protein isolates-cyanidin-3-galactoside conjugates produced using free radicals induced by ultrasound. Ultrason. Sonochem. 2020, 64, 104990. [Google Scholar] [CrossRef]
- Chen, N.; Wang, Z.; Zhu, J.; Ning, Y.; Jiang, L.; Yan, S.; Qi, B. Effect of extraction pH on the emulsion stability and surface protein structure of soybean oil body. Food Chem. 2025, 473, 143029. [Google Scholar] [CrossRef]
- Martínez-Velasco, A.; Lobato-Calleros, C.; Hernández-Rodríguez, B.E.; Román-Guerrero, A.; Alvarez-Ramirez, J.; Vernon-Carter, E.J. High intensity ultrasound treatment of faba bean (Vicia faba L.) protein: Effect on surface properties, foaming ability and structural changes. Ultrason. Sonochem. 2018, 44, 97–105. [Google Scholar] [CrossRef]
- Du, Y.; Zhang, Q.; Zhao, X.; Chen, F. Effect of reverse micelle on physicochemical properties of soybean 7S globulins. J. Food Eng. 2020, 282, 110026. [Google Scholar] [CrossRef]
- Zhang, Y.; Ma, L.; Cai, L.; Liu, Y.; Li, J. Effect of combined ultrasonic and alkali pretreatment on enzymatic preparation of angiotensin converting enzyme (ACE) inhibitory peptides from native collagenous materials. Ultrason. Sonochem. 2017, 36, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Xue, F.; Zhao, X.; Li, C.; Adhikari, B. Modification of plum seed protein isolate via enzymatic hydrolysis, polyphenol conjugation and polysaccharide complexation to enhance emulsification and encapsulation of essential oils. Int. J. Biol. Macromol. 2025, 306, 141812. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, M.; Xue, F.; Adhikari, B. Application of ultrasound treatment to improve the technofunctional properties of hemp protein isolate. Future Foods 2022, 6, 100176. [Google Scholar] [CrossRef]
- Pan, C.-P.; Muiño, P.L.; Barkley, M.D.; Callis, P.R. Correlation of Tryptophan Fluorescence Spectral Shifts and Lifetimes Arising Directly from Heterogeneous Environment. J. Phys. Chem. B 2011, 115, 3245–3253. [Google Scholar] [CrossRef]
- Plankensteiner, L.; Yang, J.; Bitter, J.H.; Vincken, J.-P.; Hennebelle, M.; Nikiforidis, C.V. High yield extraction of oleosins, the proteins that plants developed to stabilize oil droplets. Food Hydrocoll. 2023, 137, 108419. [Google Scholar] [CrossRef]
- Huang, C.-Y.; Huang, A.H.C. Unique Motifs and Length of Hairpin in Oleosin Target the Cytosolic Side of Endoplasmic Reticulum and Budding Lipid Droplet. Plant Physiol. 2017, 174, 2248–2260. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, S.-B. Structural and functional properties of self-assembled peanut protein nanoparticles prepared by ultrasonic treatment: Effects of ultrasound intensity and protein concentration. Food Chem. 2023, 413, 135626. [Google Scholar] [CrossRef]
- Plankensteiner, L.; Hennebelle, M.; Vincken, J.-P.; Nikiforidis, C.V. Insights into the emulsification mechanism of the surfactant-like protein oleosin. J. Colloid Interface Sci. 2024, 657, 352–362. [Google Scholar] [CrossRef]
- Hu, H.; Wu, J.; Li-Chan, E.C.Y.; Zhu, L.; Zhang, F.; Xu, X.; Fan, G.; Wang, L.; Huang, X.; Pan, S. Effects of ultrasound on structural and physical properties of soy protein isolate (SPI) dispersions. Food Hydrocoll. 2013, 30, 647–655. [Google Scholar] [CrossRef]
- Dabbour, M.; He, R.; Mintah, B.; Xiang, J.; Ma, H. Changes in functionalities, conformational characteristics and antioxidative capacities of sunflower protein by controlled enzymolysis and ultrasonication action. Ultrason. Sonochem. 2019, 58, 104625. [Google Scholar] [CrossRef] [PubMed]
- Resendiz-Vazquez, J.A.; Ulloa, J.A.; Urías-Silvas, J.E.; Bautista-Rosales, P.U.; Ramírez-Ramírez, J.C.; Rosas-Ulloa, P.; González-Torres, L. Effect of high-intensity ultrasound on the technofunctional properties and structure of jackfruit (Artocarpus heterophyllus) seed protein isolate. Ultrason. Sonochem. 2017, 37, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Qiao, Y.; Zhu, Y.; Shen, W.; Jin, W.; Peng, D.; Huang, Q. Assembly of oleosin during efficient extraction: Altering the sequence of defatting solvents. Food Chem. X 2025, 25, 102022. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Yan, C.; Lin, M.; He, C.; Xu, Y.; Huang, Y.; Zhou, Z. The effects of conjugation of walnut protein isolate with polyphenols on protein solubility, antioxidant activity, and emulsifying properties. Food Res. Int. 2022, 161, 111910. [Google Scholar] [CrossRef]
- Arzeni, C.; Martínez, K.; Zema, P.; Arias, A.; Pérez, O.E.; Pilosof, A.M.R. Comparative study of high intensity ultrasound effects on food proteins functionality. J. Food Eng. 2012, 108, 463–472. [Google Scholar] [CrossRef]
- Sun, Y.; Zhong, M.; Wu, L.; Huang, Y.; Li, Y.; Qi, B. Effects of ultrasound-assisted salt (NaCl) extraction method on the structural and functional properties of Oleosin. Food Chem. 2022, 372, 131238. [Google Scholar] [CrossRef]
- Lv, Y.; Chen, L.; Liu, F.; Xu, F.; Zhong, F. Improvement of the encapsulation capacity and emulsifying properties of soy protein isolate through controlled enzymatic hydrolysis. Food Hydrocoll. 2023, 138, 108444. [Google Scholar] [CrossRef]
- Qi, X.; Luo, Y.; Fei, W.; Shen, M.; Chen, Y.; Yu, Q.; Xie, J. Effects of enzyme hydrolysis-assisted fibrillation treatment on the solubility, emulsifying properties and antioxidant activity of rice protein. Int. J. Biol. Macromol. 2024, 279, 135378. [Google Scholar] [CrossRef]
- Yuliana, M.; Truong, C.T.; Huynh, L.H.; Ho, Q.P.; Ju, Y.-H. Isolation and characterization of protein isolated from defatted cashew nut shell: Influence of pH and NaCl on solubility and functional properties. LWT—Food Sci. Technol. 2014, 55, 621–626. [Google Scholar] [CrossRef]
- Guo, Y.; Luo, Y.; Ren, Z.; Zhang, X.; Duan, H.; Liu, Z.; Wang, X. Food-Grade Pickering Emulsions Stabilized by Ultrasound-Treated Foxtail Millet Prolamin: Characterization and In Vitro Release Behavior of Curcumin. Foods 2025, 14, 417. [Google Scholar] [CrossRef]
- Liu, Q.; Chen, T.; Chen, L.; Zhao, R.; Ye, X.; Wang, X.; Wu, D.; Hu, J. High Internal Phase Emulsions Stabilized with Ultrasound-Modified Spirulina Protein for Curcumin Delivery. Foods 2024, 13, 1324. [Google Scholar] [CrossRef] [PubMed]
- McClements, D.J.; Jafari, S.M. Improving emulsion formation, stability and performance using mixed emulsifiers: A review. Adv. Colloid Interface Sci. 2018, 251, 55–79. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Fan, X.; Yu, X.; Liu, T.; Guo, X.; Zhang, J. Enhancing curcumin stability and bioavailability through chickpea protein isolate–citrus pectin conjugate emulsions: Targeted delivery and gut microecology modulation. Int. J. Biol. Macromol. 2025, 300, 140295. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Jia, Q.; Li, Y.; Zhao, H.; Wu, Z. Chitosan/OSA starch composite-stabilized Pickering emulsions: Characterization and its application in curcumin. LWT 2025, 222, 117661. [Google Scholar] [CrossRef]
- Guo, Q.; Qi, X.; Chen, X.; Yang, Y.; Wang, S. Development of a stable curcumin-loaded emulsion with desired properties under a low oil phase proportion using starch-palmitic acid-β-lactoglobulin complexes. Int. J. Biol. Macromol. 2025, 309, 143114. [Google Scholar] [CrossRef]
- Zheng, L.; Chen, X.; Zheng, L.; Chang, Q.; Qin, Y.; Ding, X.; Zhang, W.; Zhao, P.; Xi, C. Curcumin-loaded WPI-FUC emulsion as a microcapsule core material enhances the applicability and stability of curcumin—Preparation and characterization of a new type of microcapsule. LWT 2025, 224, 117854. [Google Scholar] [CrossRef]
- Zhang, L.; Chen, D.-L.; Wang, X.-F.; Qian, J.-Y.; He, X.-D. Enzymatically modified quinoa starch-based Pickering emulsion: Effect of enzymolysis and emulsifying conditions. Int. J. Biol. Macromol. 2022, 219, 824–834. [Google Scholar] [CrossRef]
- Xia, B.; Liu, Y.; Dong, C.; Shen, Y.; Wang, C. Enhancing the usability of pea protein in emulsion applications through modification by various approaches: A comparative study. Food Res. Int. 2024, 188, 114477. [Google Scholar] [CrossRef]
- Ma, Z.; Bitter, J.H.; Boom, R.M.; Nikiforidis, C.V. Encapsulation of cannabidiol in hemp seed oleosomes. Food Res. Int. 2024, 195, 114948. [Google Scholar] [CrossRef]
- Ji, L.; Feng, W.; Chen, H.; Chu, Y.; Wong, A.; Zhu, Y.; Sinatra, G.; Bramante, F.; Carrière, F.; Stocks, M.J.; et al. Rapeseed oleosomes facilitate intestinal lymphatic delivery and oral bioavailability of cannabidiol. Int. J. Pharm. 2025, 668, 124947. [Google Scholar] [CrossRef]
- Kang, M.; Song, H.; Liao, Y.; Wang, S.; Yan, S.; Qi, B. Preparation of a curcumin oil bodies delivery system based on pH-Shift treatment: Physicochemical properties, protein structure, and digestive properties. LWT 2024, 200, 116218. [Google Scholar] [CrossRef]
- Lin, Z.; Zhou, P.; Deng, Y.; Liu, G.; Li, P.; Zeng, J.; Zhang, Y.; Tang, X.; Zhao, Z.; Zhang, M. Impact of homogenization methods on the interfacial protein composition and stability of peanut oil body emulsion with sodium caseinate and maltodextrin. LWT 2025, 215, 117286. [Google Scholar] [CrossRef]
- Karaca, A.C.; Low, N.; Nickerson, M. Emulsifying properties of chickpea, faba bean, lentil and pea proteins produced by isoelectric precipitation and salt extraction. Food Res. Int. 2011, 44, 2742–2750. [Google Scholar] [CrossRef]
- Dalla, E.; Koumentakou, I.; Bikiaris, N.; Balla, E.; Lykidou, S.; Nikolaidis, N. Formulation, Characterization and Evaluation of Innovative O/W Emulsions Containing Curcumin Derivatives with Enhanced Antioxidant Properties. Antioxidants 2022, 11, 2271. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Jiang, Y.; Shi, J. Novel Pickering emulsion stabilized by glycosylated whey protein isolate: Characterization, stability, and curcumin bioaccessibility. Food Chem. X 2024, 21, 101186. [Google Scholar] [CrossRef]
- Liu, C.; Deng, Z.; Wang, L.; Zhang, M.; Liu, J. Complexation between curcumin and walnut protein isolate modified by pH shifting combined with protein-glutaminase. Food Chem. 2025, 464, 141693. [Google Scholar] [CrossRef]
- Yang, Z.; Zeng, S.; Tang, J.; Yang, L.; Jin, B. Construction of high internal phase Pickering emulsion using dual protein hydrolysates-naringenin supramolecules and their contribution on curcumin bioaccessibility. J. Mol. Liq. 2025, 430, 127700. [Google Scholar] [CrossRef]
- Chen, Q.; Di, X.; Zhai, Y.; Zhao, Q.; Song, X. Influence of oil phases on the digestibility and curcumin delivery properties of Pickering emulsions. Food Chem. X 2025, 26, 102270. [Google Scholar] [CrossRef]




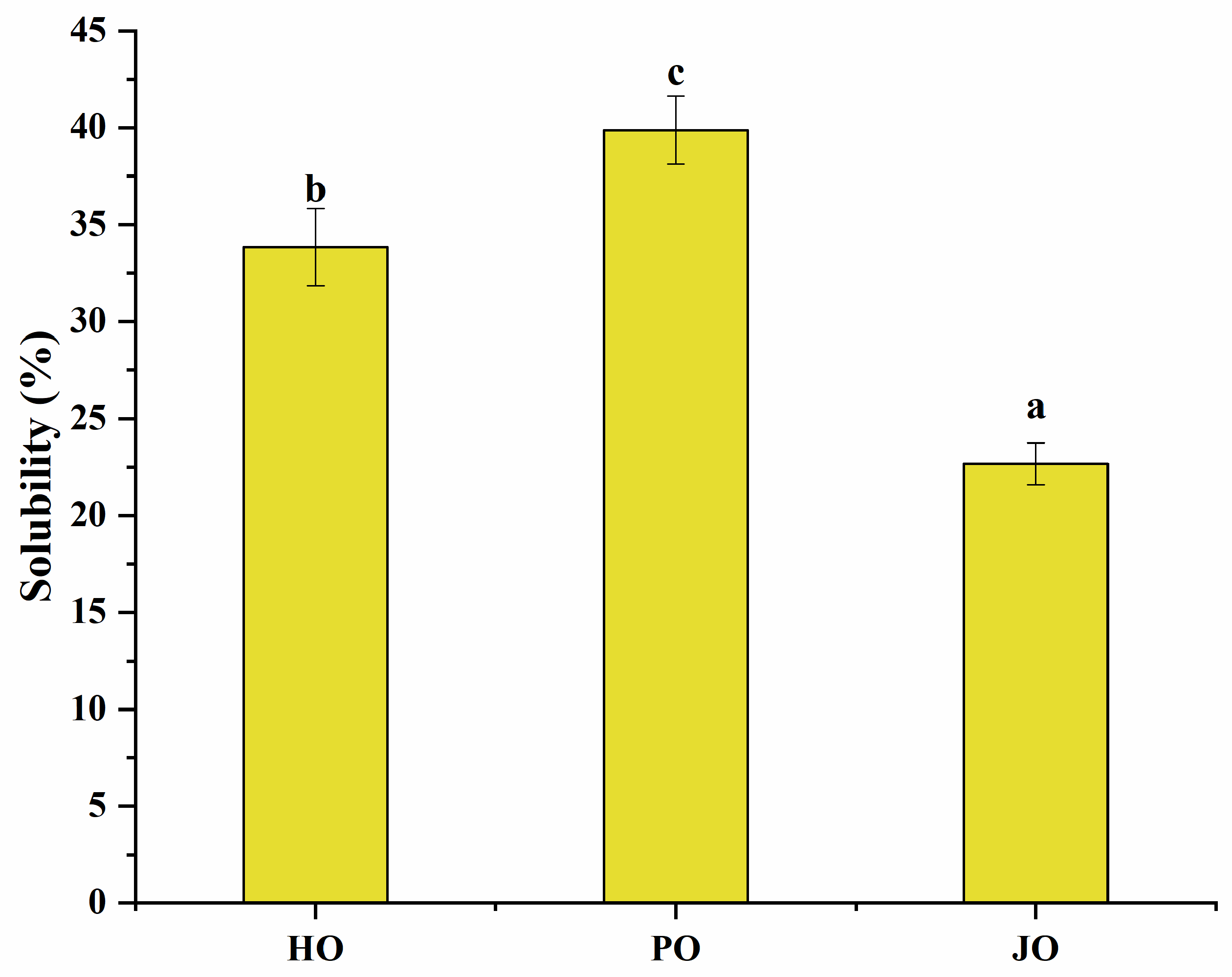




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, Y.; Hu, Q.; Xue, F. Comparative Characterization of Oil Body Proteins from Hemp, Plum, and Jujube Seed and Their Application in Curcumin-Loaded Artificial Oleosomes. Polymers 2025, 17, 1346. https://doi.org/10.3390/polym17101346
Cao Y, Hu Q, Xue F. Comparative Characterization of Oil Body Proteins from Hemp, Plum, and Jujube Seed and Their Application in Curcumin-Loaded Artificial Oleosomes. Polymers. 2025; 17(10):1346. https://doi.org/10.3390/polym17101346
Chicago/Turabian StyleCao, Yuhan, Qin Hu, and Feng Xue. 2025. "Comparative Characterization of Oil Body Proteins from Hemp, Plum, and Jujube Seed and Their Application in Curcumin-Loaded Artificial Oleosomes" Polymers 17, no. 10: 1346. https://doi.org/10.3390/polym17101346
APA StyleCao, Y., Hu, Q., & Xue, F. (2025). Comparative Characterization of Oil Body Proteins from Hemp, Plum, and Jujube Seed and Their Application in Curcumin-Loaded Artificial Oleosomes. Polymers, 17(10), 1346. https://doi.org/10.3390/polym17101346