
Natural Compounds Interacting with Nicotinic Acetylcholine Receptors: From Low-Molecular Weight Ones to Peptides and Proteins

 ,
, 

 and
and
Abstract
:1. Introduction
2. Low-Molecular Weight Agonists and Antagonists of nAChRs
2.1. Short Summary of Well-Known Agonists and Antagonists of nAChRs

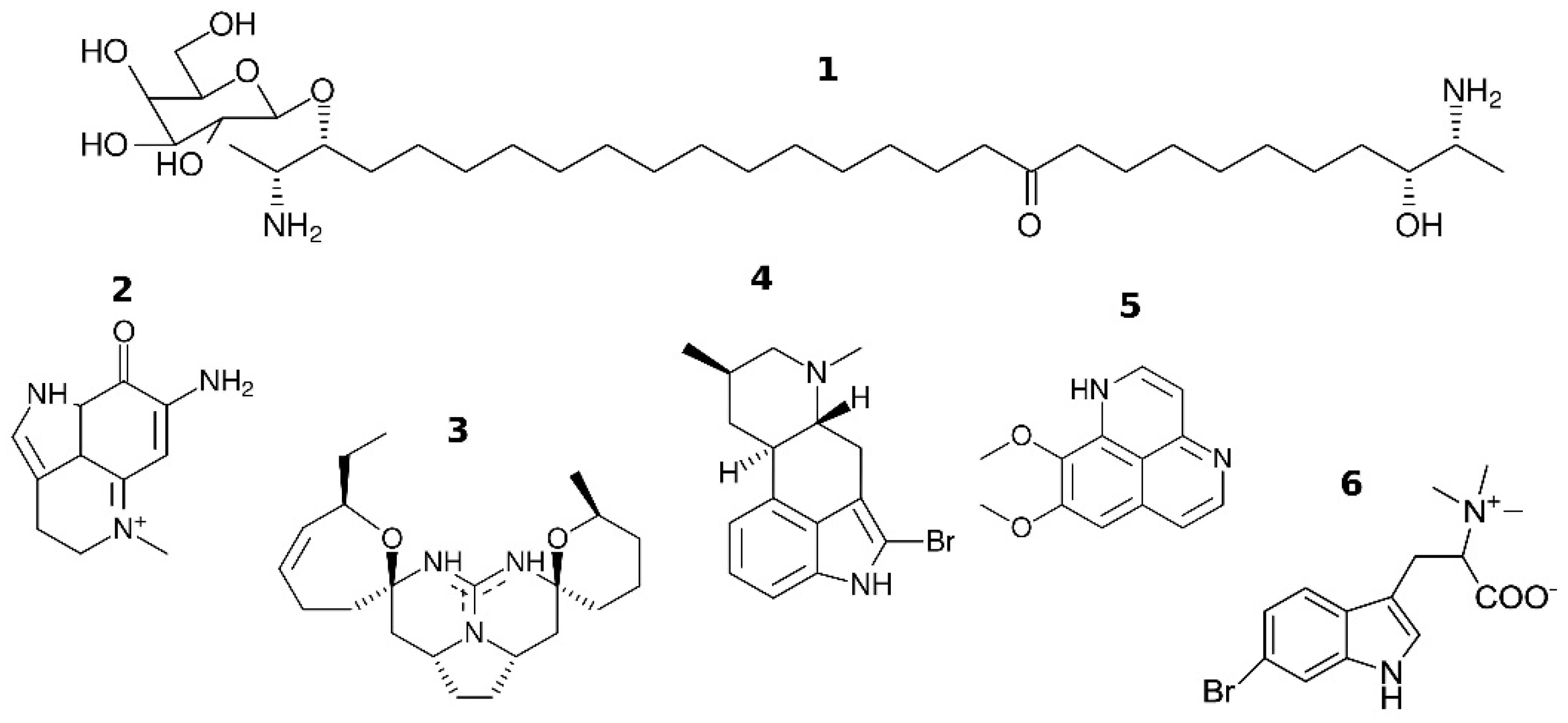{kind=link}
{kind=link}
| Compound | Source | Activity |
|---|---|---|
| acetylcholine | - | Prototypic agonist at all nicotinic receptors |
| choline | - | Agonist at α7 and muscle nAChRs |
| nicotine | Tobacco plant | Agonist at most nAChR subtypes; antagonist at α9 nAChR |
| epibatidine | Epipedobates frogs | Agonist at most nAChR subtypes; antagonist at α9 nAChR |
| cytisine | Plants of Fabaceae family | Partial agonist at neuronal nAChRs |
| anatoxin-a | Cyanobacteria | Non-selective agonist of nAChRs |
| anabaseine | Certain species of ants and marine worms | Agonist at neuronal nAChRs |
| d-tubocurarine | Chondodendron tomentosum plant | Non-selective antagonist |
| coniine | Conium maculatum plant | Antagonist at muscle nAChRs |
| pinnatoxins, 1,3 desmethyl spirolide, gymnodimines | dinoflagellates | Non-selective antagonists |
| pictamine | Clavelina picta ascidian | Antagonist at neuronal nAChRs |
2.2. nAChR Antagonists and Agonists of Marine Origin

3. α-Conotoxins and Other Peptides Interacting with nAChRs
3.1. Naturally-Occurring α-Conotoxins from Conus Marine Snails
3.2. Design and Synthesis of More Potent and Selective α-Conotoxin Analogs
| Substitutions in PnIA | Affinity (IC50, nM) for | ||||
|---|---|---|---|---|---|
| AChBPs in Competition with [125I]-αBgt | Human α7 nAChR | ||||
| L. stagnalis | A. californica | in competition with | in electrophysiology | ||
| [125I]-αBgt | [125I]-α-conotoxin | ||||
| α-conotoxin PnIA: GCCSPCNNPYC-NH2 | |||||
| [H5] | 220 | 3.1 | 26,000 | - | - |
| [H5, R14] | 2,900 | 1,400 | 21,000 | - | - |
| [L10] | 200 | 55 | 14,000 | - | - |
| [D5, L10] | 35,000 | 5,200 | >100,000 | - | - |
| [R5, L10] | 180 | 305 | 12,000 | - | - |
| [D7, L10] | >>100,000 | 63,000 | >>100,000 | - | - |
| [R7, L10] | 160,000 | 1,250 | >>100,000 | - | - |
| [L10, K14] | 8.2 | 47 | 7,200 | 1,800 1 | 260 |
| [D5, R7, L10] | 38,000 | 51 | 275,000 | - | - |
| [D5, R7, V10] | 6,400 | 45 | >100,000 | - | - |
| [R5, D7, L10] | 56,000 | 28,000 | >>100,000 | - | - |
| [R5, L10, R14] | 430 | 1,400 | 670 | 60 1 | - |
| [D5, R7, L10, R14] | 1,200 | 46 | 23,000 | - | - |
| [R5, D7, L10, R14] | 4,100 | 3,200 | 72,000 | - | - |
| [R9] | 58 | 41 | 2,400 | 1,490 2 | 27 |
| [R9, L10] | 18 | 47 | 270 | 36 2 | 17 |
| [R5, R9, L10, R14] | 1.2 | 24 | 860 | 390 2 | 27 |
3.3. nAChR -Inhibiting Peptides from Snake Venoms

4. Proteins Interacting with nAChRs
4.1. Monomeric and Dimeric α-Neurotoxins and Non-Conventional Toxins
4.2. Three-Finger Ly6 Proteins of the Non-Venomous Origin
4.3. Snake Venom Phospholipases A2 and C-Type Lectin-like Proteins as Inhibitors of nAChRs
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Changeux, J.P. The nicotinic acetylcholine receptor: The founding father of the pentameric ligand-gated ion channel superfamily. J. Biol. Chem. 2012, 287, 40207–40215. [Google Scholar] [CrossRef] [PubMed]
- Yakel, J.L. Cholinergic receptors: Functional role of nicotinic ACh receptors in brain circuits and disease. Pflugers. Arch. Eur. J. Physiol. 2013, 465, 441–450. [Google Scholar] [CrossRef]
- Corringer, P.J.; Poitevin, F.; Prevost, M.S.; Sauguet, L.; Delarue, M.; Changeux, J.P. Structure and pharmacology of pentameric receptor channels: From bacteria to brain. Structure 2012, 20, 941–956. [Google Scholar] [CrossRef] [PubMed]
- Dineley, K.T.; Pandya, A.A.; Yakel, J.L. Nicotinic ACh receptors as therapeutic targets in CNS disorders. Trends Pharmacol. Sci. 2015, 36, 96–108. [Google Scholar] [CrossRef] [PubMed]
- Lombardo, S.; Maskos, U. Role of the nicotinic acetylcholine receptor in Alzheimer’s disease pathology and treatment. Neuropharmacology 2014. [Google Scholar] [CrossRef]
- Farrugia, M.E.; Vincent, A. Autoimmune mediated neuromuscular junction defects. Curr. Opin. Neurol. 2010, 23, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Unwin, N. Refined structure of the nicotinic acetylcholine receptor at 4A resolution. J. Mol. Biol. 2005, 346, 967–989. [Google Scholar] [CrossRef] [PubMed]
- Unwin, N. Nicotinic acetylcholine receptor and the structural basis of neuromuscular transmission: Insights from Torpedo postsynaptic membranes. Q. Rev. Biophys. 2013, 46, 283–322. [Google Scholar] [CrossRef] [PubMed]
- Brejc, K.; van Dijk, W.J.; Klaassen, R.V.; Schuurmans, M.; van Der Oost, J.; Smit, A.B.; Sixma, T.K. Crystal structure of an ACh-binding protein reveals the ligand-binding domain of nicotinic receptors. Nature 2001, 411, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Celie, P.H.N.; Kasheverov, I.E.; Mordvintsev, D.Y.; Hogg, R.C.; van Nierop, P.; van Elk, R.; van Rossum-Fikkert, S.E.; Zhmak, M.N.; Bertrand, D.; Tsetlin, V.; et al. Crystal structure of nicotinic acetylcholine receptor homolog AChBP in complex with an alpha-conotoxin PnIA variant. Nat. Struct. Mol. Biol. 2005, 12, 582–588. [Google Scholar] [CrossRef] [PubMed]
- Celie, P.H.N.; van Rossum-Fikkert, S.E.; van Dijk, W.J.; Brejc, K.; Smit, A.B.; Sixma, T.K. Nicotine and carbamylcholine binding to nicotinic acetylcholine receptors as studied in AChBP crystal structures. Neuron 2004, 41, 907–914. [Google Scholar] [CrossRef] [PubMed]
- Hansen, S.B.; Sulzenbacher, G.; Huxford, T.; Marchot, P.; Taylor, P.; Bourne, Y. Structures of Aplysia AChBP complexes with nicotinic agonists and antagonists reveal distinctive binding interfaces and conformations. EMBO J. 2005, 24, 3635–3646. [Google Scholar] [CrossRef] [PubMed]
- Li, S.-X.; Huang, S.; Bren, N.; Noridomi, K.; Dellisanti, C.D.; Sine, S.M.; Chen, L. Ligand-binding domain of an α7-nicotinic receptor chimera and its complex with agonist. Nat. Neurosci. 2011, 14, 1253–1259. [Google Scholar] [CrossRef] [PubMed]
- Brams, M.; Pandya, A.; Kuzmin, D.; van Elk, R.; Krijnen, L.; Yakel, J.L.; Tsetlin, V.; Smit, A.B.; Ulens, C. A structural and mutagenic blueprint for molecular recognition of strychnine and d-tubocurarine by different cys-loop receptors. PLoS Biol. 2011, 9, e1001034. [Google Scholar] [CrossRef] [PubMed]
- Bourne, Y.; Talley, T.T.; Hansen, S.B.; Taylor, P.; Marchot, P. Crystal structure of a Cbtx-AChBP complex reveals essential interactions between snake alpha-neurotoxins and nicotinic receptors. EMBO J. 2005, 24, 1512–1522. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Li, S.-X.; Bren, N.; Cheng, K.; Gomoto, R.; Chen, L.; Sine, S.M. Complex between α-bungarotoxin and an α7 nicotinic receptor ligand-binding domain chimaera. Biochem. J. 2013, 454, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Ulens, C.; Hogg, R.C.; Celie, P.H.; Bertrand, D.; Tsetlin, V.; Smit, A.B.; Sixma, T.K. Structural determinants of selective alpha-conotoxin binding to a nicotinic acetylcholine receptor homolog AChBP. Proc. Natl. Acad. Sci. USA 2006, 103, 3615–3620. [Google Scholar] [CrossRef] [PubMed]
- Dutertre, S.; Ulens, C.; Büttner, R.; Fish, A.; van Elk, R.; Kendel, Y.; Hopping, G.; Alewood, P.F.; Schroeder, C.; Nicke, A.; et al. AChBP-targeted alpha-conotoxin correlates distinct binding orientations with nAChR subtype selectivity. EMBO J. 2007, 26, 3858–3867. [Google Scholar] [CrossRef] [PubMed]
- Tsetlin, V.; Utkin, Y.; Kasheverov, I. Polypeptide and peptide toxins, magnifying lenses for binding sites in nicotinic acetylcholine receptors. Biochem. Pharmacol. 2009, 78, 720–731. [Google Scholar] [CrossRef] [PubMed]
- Utkin, Y.N. Three-finger toxins, a deadly weapon of elapid venom--milestones of discovery. Toxicon 2013, 62, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Shelukhina, I.V.; Kryukova, E.V.; Lips, K.S.; Tsetlin, V.I.; Kummer, W. Presence of alpha7 nicotinic acetylcholine receptors on dorsal root ganglion neurons proved using knockout mice and selective alpha-neurotoxins in histochemistry. J. Neurochem. 2009, 109, 1087–1095. [Google Scholar] [CrossRef] [PubMed]
- Frahm, S.; Slimak, M.A.; Ferrarese, L.; Santos-Torres, J.; Antolin-Fontes, B.; Auer, S.; Filkin, S.; Pons, S.; Fontaine, J.-F.; Tsetlin, V.; et al. Aversion to nicotine is regulated by the balanced activity of β4 and α5 nicotinic receptor subunits in the medial habenula. Neuron 2011, 70, 522–535. [Google Scholar] [CrossRef] [PubMed]
- Olivera, B.M. Conus peptides: Biodiversity-Based discovery and exogenomics. J. Biol. Chem. 2006, 281, 31173–31177. [Google Scholar] [CrossRef] [PubMed]
- Kasheverov, I.E.; Utkin, Y.N.; Tsetlin, V.I. Naturally occurring and synthetic peptides acting on nicotinic acetylcholine receptors. Curr. Pharm. Des. 2009, 15, 2430–2452. [Google Scholar] [CrossRef] [PubMed]
- Prashanth, J.R.; Lewis, R.J.; Dutertre, S. Towards an integrated venomics approach for accelerated conopeptide discovery. Toxicon 2012, 60, 470–477. [Google Scholar] [CrossRef] [PubMed]
- Otero, A.; Chapela, M.-J.; Atanassova, M.; Vieites, J.M.; Cabado, A.G. Cyclic Imines: Chemistry and Mechanism of Action: A Review. Chem. Res. Toxicol. 2011, 24, 1817–1829. [Google Scholar] [CrossRef] [PubMed]
- Bourne, Y.; Radic, Z.; Aráoz, R.; Talley, T.T.; Benoit, E.; Servent, D.; Taylor, P.; Molgó, J.; Marchot, P. Structural determinants in phycotoxins and AChBP conferring high affinity binding and nicotinic AChR antagonism. Proc. Natl. Acad. Sci. USA 2010, 107, 6076–6081. [Google Scholar] [CrossRef] [PubMed]
- Wandscheer, C.B.; Vilariño, N.; Espiña, B.; Louzao, M.C.; Botana, L.M. Human muscarinic acetylcholine receptors are a target of the marine toxin 13-desmethyl C spirolide. Chem. Res. Toxicol. 2010, 23, 1753–1761. [Google Scholar] [CrossRef] [PubMed]
- Munday, R.; Quilliam, M.A.; LeBlanc, P.; Lewis, N.; Gallant, P.; Sperker, S.A.; Ewart, H.S.; MacKinnon, S.L. Investigations into the toxicology of spirolides, a group of marine phycotoxins. Toxins (Basel) 2012, 4, 1–14. [Google Scholar] [CrossRef]
- Kudryavtsev, D.; Makarieva, T.; Utkina, N.; Santalova, E.; Kryukova, E.; Methfessel, C.; Tsetlin, V.; Stonik, V.; Kasheverov, I. Marine natural products acting on the acetylcholine-binding protein and nicotinic receptors: From computer modeling to binding studies and electrophysiology. Mar. Drugs 2014, 12, 1859–1875. [Google Scholar] [CrossRef] [PubMed]
- Molgó, J.; Aráoz, R.; Benoit, E.; Iorga, B.I. Physical and virtual screening methods for marine toxins and drug discovery targeting nicotinic acetylcholine receptors. Expert Opin. Drug Discov. 2013, 8, 1203–1223. [Google Scholar] [CrossRef] [PubMed]
- Philpot, R.M. Potential Use of Nicotinic Receptor Agonists for the Treatment of Chemotherapy-Induced Cognitive Deficits. Neurochem. Res. 2015. [Google Scholar] [CrossRef]
- Barbier, A.J.; Hilhorst, M.; van Vliet, A.; Snyder, P.; Palfreyman, M.G.; Gawryl, M.; Dgetluck, N.; Massaro, M.; Tiessen, R.; Timmerman, W.; et al. Pharmacodynamics, Pharmacokinetics, Safety, and Tolerability of Encenicline, a Selective α7 Nicotinic Receptor Partial Agonist, in Single Ascending-dose and Bioavailability Studies. Clin. Ther. 2015, 37, 311–324. [Google Scholar] [PubMed]
- Dutta, S.; Hosmane, B.S.; Awni, W.M. Population analyses of efficacy and safety of ABT-594 in subjects with diabetic peripheral neuropathic pain. AAPS J. 2012, 14, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Kasheverov, I.; Shelukhina, I.; Kudryavtsev, D.; Makarieva, T.; Spirova, E.; Guzii, A.; Stonik, V.; Tsetlin, V. 6-Bromohypaphorine from Marine Nudibranch Mollusk Hermissenda crassicornis is an Agonist of Human α7 Nicotinic Acetylcholine Receptor. Mar. Drugs 2015, 13, 1255–1266. [Google Scholar] [CrossRef] [PubMed]
- Muttenthaler, M.; Akondi, K.B.; Alewood, P.F. Structure-activity studies on alpha-conotoxins. Curr. Pharm. Des. 2011, 17, 4226–4241. [Google Scholar] [CrossRef] [PubMed]
- Lebbe, E.K.M.; Peigneur, S.; Wijesekara, I.; Tytgat, J. Conotoxins targeting nicotinic acetylcholine receptors: An overview. Mar. Drugs 2014, 12, 2970–3004. [Google Scholar] [CrossRef] [PubMed]
- Prashanth, J.R.; Brust, A.; Jin, A.-H.; Alewood, P.F.; Dutertre, S.; Lewis, R.J. Cone snail venomics: from novel biology to novel therapeutics. Future Med. Chem. 2014, 6, 1659–1675. [Google Scholar] [CrossRef] [PubMed]
- Del Bufalo, A.; Cesario, A.; Salinaro, G.; Fini, M.; Russo, P. Alpha9 alpha10 nicotinic acetylcholine receptors as target for the treatment of chronic pain. Curr. Pharm. Des. 2014, 20, 6042–6047. [Google Scholar] [CrossRef]
- Kauferstein, S.; Porth, C.; Kendel, Y.; Wunder, C.; Nicke, A.; Kordis, D.; Favreau, P.; Koua, D.; Stöcklin, R.; Mebs, D. Venomic study on cone snails (Conus spp.) from South Africa. Toxicon 2011, 57, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Peigneur, S.; van Der Haegen, A.; Möller, C.; Waelkens, E.; Diego-García, E.; Marí, F.; Naudé, R.; Tytgat, J. Unraveling the peptidome of the South African cone snails Conus pictus and Conus natalis. Peptides 2013, 41, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Ye, M.; Wang, Y.; Shao, X.; Yuan, D.; Liu, J.; Hawrot, E.; Wang, C.; Chi, C. A new subfamily of conotoxins belonging to the A-superfamily. Peptides 2010, 31, 2009–2016. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Zhangsun, D.; Wu, Y.; Zhu, X.; Hu, Y.; McIntyre, M.; Christensen, S.; Akcan, M.; Craik, D.J.; McIntosh, J.M. Characterization of a novel α-conotoxin from conus textile that selectively targets α6/α3β2β3 nicotinic acetylcholine receptors. J. Biol. Chem. 2013, 288, 894–902. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Li, H.; Liu, N.; Wu, C.; Jiang, J.; Yue, J.; Jing, Y.; Dai, Q. Diversity and evolution of conotoxins in Conus virgo, Conus eburneus, Conus imperialis and Conus marmoreus from the South China Sea. Toxicon 2012, 60, 982–989. [Google Scholar] [CrossRef]
- Inserra, M.C.; Kompella, S.N.; Vetter, I.; Brust, A.; Daly, N.L.; Cuny, H.; Craik, D.J.; Alewood, P.F.; Adams, D.J.; Lewis, R.J. Isolation and characterization of α-conotoxin LsIA with potent activity at nicotinic acetylcholine receptors. Biochem. Pharmacol. 2013, 86, 791–799. [Google Scholar] [CrossRef] [PubMed]
- Lebbe, E.K.M.; Peigneur, S.; Maiti, M.; Mille, B.G.; Devi, P.; Ravichandran, S.; Lescrinier, E.; Waelkens, E.; D’Souza, L.; Herdewijn, P.; et al. Discovery of a new subclass of α-conotoxins in the venom of Conus australis. Toxicon 2014, 91, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, B.; le Caer, J.-P.; Aráoz, R.; Thai, R.; Lamthanh, H.; Benoit, E.; Molgó, J. Isolation, purification and functional characterization of alpha-BnIA from Conus bandanus venom. Toxicon 2014, 91, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Morales-González, D.; Flores-Martínez, E.; Zamora-Bustillos, R.; Rivera-Reyes, R.; Michel-Morfín, J.E.; Landa-Jaime, V.; Falcón, A.; Aguilar, M.B. Diversity of A-conotoxins of three worm-hunting cone snails (Conus brunneus, Conus nux, and Conus princeps) from the Mexican Pacific coast. Peptides 2015. [Google Scholar] [CrossRef]
- Whiteaker, P.; Christensen, S.; Yoshikami, D.; Dowell, C.; Watkins, M.; Gulyas, J.; Rivier, J.; Olivera, B.M.; McIntosh, J.M. Discovery, synthesis, and structure activity of a highly selective alpha7 nicotinic acetylcholine receptor antagonist. Biochemistry 2007, 46, 6628–6638. [Google Scholar] [CrossRef]
- Armishaw, C.J.; Singh, N.; Medina-Franco, J.L.; Clark, R.J.; Scott, K.C.M.; Houghten, R.A.; Jensen, A.A. A synthetic combinatorial strategy for developing alpha-conotoxin analogs as potent alpha7 nicotinic acetylcholine receptor antagonists. J. Biol. Chem. 2010, 285, 1809–1821. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.-P.; Banerjee, J.; Dowell, C.; Wu, J.; Gyanda, R.; Houghten, R.A.; Toll, L.; McIntosh, J.M.; Armishaw, C.J. Discovery of a potent and selective α3β4 nicotinic acetylcholine receptor antagonist from an α-conotoxin synthetic combinatorial library. J. Med. Chem. 2014, 57, 3511–3521. [Google Scholar] [CrossRef] [PubMed]
- Azam, L.; Yoshikami, D.; McIntosh, J.M. Amino acid residues that confer high selectivity of the alpha6 nicotinic acetylcholine receptor subunit to alpha-conotoxin MII[S4A,E11A,L15A]. J. Biol. Chem. 2008, 283, 11625–11632. [Google Scholar] [CrossRef] [PubMed]
- Hone, A.J.; Scadden, M.; Gajewiak, J.; Christensen, S.; Lindstrom, J.; McIntosh, J.M. α-Conotoxin PeIA[S9H,V10A,E14N] potently and selectively blocks α6β2β3 versus α6β4 nicotinic acetylcholine receptors. Mol. Pharmacol. 2012, 82, 972–982. [Google Scholar] [CrossRef] [PubMed]
- Olivera, B.M.; Quik, M.; Vincler, M.; McIntosh, J.M. Subtype-selective conopeptides targeted to nicotinic receptors: Concerted discovery and biomedical applications. Channels (Austin) 2008, 2, 143–152. [Google Scholar]
- Yang, K.; Jin, G.; Wu, J. Mysterious alpha6-containing nAChRs: Function, pharmacology, and pathophysiology. Acta Pharmacol. Sin. 2009, 30, 740–751. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Alvarez, A.; Hernández-Vivanco, A.; McIntosh, J.M.; Albillos, A. Native α6β4* nicotinic receptors control exocytosis in human chromaffin cells of the adrenal gland. FASEB J. 2012, 26, 346–354. [Google Scholar] [CrossRef] [PubMed]
- Surin, A.M.; Kriukova, E.V; Strukov, A.S.; Zhmak, M.N.; Talka, R.; Tuominen, R.; Salminen, O.; Khiroug, L.; Kasheverov, I.E.; Tsetlin, V.I. Effect of alpha-conotoxin MII and its N-terminal derivatives on Ca2+ and Na+ signals induced by nicotine in neuroblastoma cell line SH-SY5Y. Bioorg. Khim. 2012, 38, 214–222. [Google Scholar] [PubMed]
- Hone, A.J.; Whiteaker, P.; Mohn, J.L.; Jacob, M.H.; McIntosh, J.M. Alexa Fluor 546-ArIB[V11L;V16A] is a potent ligand for selectively labeling alpha 7 nicotinic acetylcholine receptors. J. Neurochem. 2010, 114, 994–1006. [Google Scholar] [PubMed]
- Kasheverov, I.E.; Zhmak, M.N.; Khruschov, A.Y.; Tsetlin, V.I. Design of new α-conotoxins: From computer modeling to synthesis of potent cholinergic compounds. Mar. Drugs 2011, 9, 1698–1714. [Google Scholar] [CrossRef] [PubMed]
- Koromyslova, A.D.; Chugunov, A.O.; Efremov, R.G. Deciphering fine molecular details of proteins’ structure and function with a Protein Surface Topography (PST) method. J. Chem. Inf. Model. 2014, 54, 1189–1199. [Google Scholar] [CrossRef] [PubMed]
- Kasheverov, I.E.; Kudryavtsev, D.S.; Ivanov, I.A.; Zhmak, M.N.; Chugunov, A.O.; Tabakmakher, V.M.; Zelepuga, E.A.; Efremov, R.G.; Tsetlin, V.I. Rational design of new ligands for nicotinic receptors on the basis of α-conotoxin PnIA. Dokl. Biochem. Biophys. 2015, 461, 106–109. [Google Scholar] [CrossRef] [PubMed]
- Weinstein, S.A.; Schmidt, J.J.; Bernheimer, A.W.; Smith, L.A. Characterization and amino acid sequences of two lethal peptides isolated from venom of Wagler’s pit viper, Trimeresurus wagleri. Toxicon 1991, 29, 227–236. [Google Scholar] [CrossRef]
- Utkin, Y.N.; Weise, C.; Kasheverov, I.E.; Andreeva, T.V.; Kryukova, E.V.; Zhmak, M.N.; Starkov, V.G.; Hoang, N.A.; Bertrand, D.; Ramerstorfer, J.; et al. Azemiopsin from Azemiops feae viper venom, a novel polypeptide ligand of nicotinic acetylcholine receptor. J. Biol. Chem. 2012, 287, 27079–27086. [Google Scholar] [CrossRef] [PubMed]
- Brust, A.; Sunagar, K.; Undheim, E.A.; Vetter, I.; Yang, D.C.; Yang, D.C.; Casewell, N.R.; Jackson, T.N.W.; Koludarov, I.; Alewood, P.F.; et al. Differential evolution and neofunctionalization of snake venom metalloprotease domains. Mol. Cell. Proteomics 2013, 12, 651–663. [Google Scholar] [CrossRef] [PubMed]
- Thévenet, P.; Shen, Y.; Maupetit, J.; Guyon, F.; Derreumaux, P.; Tufféry, P. PEP-FOLD: An updated de novo structure prediction server for both linear and disulfide bonded cyclic peptides. Nucleic Acids Res. 2012, 40, 288–293. [Google Scholar] [CrossRef]
- Osipov, A.V; Rucktooa, P.; Kasheverov, I.E.; Filkin, S.Y.; Starkov, V.G.; Andreeva, T.V; Sixma, T.K.; Bertrand, D.; Utkin, Y.N.; Tsetlin, V.I. Dimeric α-cobratoxin X-ray structure: Localization of intermolecular disulfides and possible mode of binding to nicotinic acetylcholine receptors. J. Biol. Chem. 2012, 287, 6725–6734. [Google Scholar] [CrossRef] [PubMed]
- Lyukmanova, E.N.; Shenkarev, Z.O.; Shulepko, M.A.; Mineev, K.S.; D’Hoedt, D.; Kasheverov, I.E.; Filkin, S.Y.; Krivolapova, A.P.; Janickova, H.; Dolezal, V.; et al. NMR structure and action on nicotinic acetylcholine receptors of water-soluble domain of human LYNX1. J. Biol. Chem. 2011, 286, 10618–10627. [Google Scholar] [CrossRef]
- Guex, N.; Peitsch, M.C.; Schwede, T. Automated comparative protein structure modeling with SWISS-MODEL and Swiss-PdbViewer: A historical perspective. Electrophoresis 2009, 30, 162–173. [Google Scholar] [CrossRef]
- Tsetlin, V.I. Three-finger snake neurotoxins and Ly6 proteins targeting nicotinic acetylcholine receptors: Pharmacological tools and endogenous modulators. Trends Pharmacol. Sci. 2015, 36, 109–123. [Google Scholar] [CrossRef] [PubMed]
- Kini, R.M.; Doley, R. Structure, function and evolution of three-finger toxins: Mini proteins with multiple targets. Toxicon 2010, 56, 855–867. [Google Scholar] [CrossRef] [PubMed]
- Barber, C.M.; Isbister, G.K.; Hodgson, W.C. Alpha neurotoxins. Toxicon 2013, 66, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.I.; Reeks, T.; Vetter, I.; Vergara, I.; Kovtun, O.; Lewis, R.J.; Alewood, P.F.; Durek, T. Isolation and Structural and Pharmacological Characterization of α-Elapitoxin-Dpp2d, an Amidated Three Finger Toxin from Black Mamba Venom. Biochemistry 2014, 53, 3758–3766. [Google Scholar] [CrossRef]
- Blacklow, B.; Kornhauser, R.; Hains, P.G.; Loiacono, R.; Escoubas, P.; Graudins, A.; Nicholson, G.M. α-Elapitoxin-Aa2a, a long-chain snake α-neurotoxin with potent actions on muscle (α1)(2)βγδ nicotinic receptors, lacks the classical high affinity for neuronal α7 nicotinic receptors. Biochem. Pharmacol. 2011, 81, 314–325. [Google Scholar] [CrossRef] [PubMed]
- Marcon, F.; Leblanc, M.; Vetter, I.; Lewis, R.J.; Escoubas, P.; Nicholson, G.M. Pharmacological characterization of α-elapitoxin-Al2a from the venom of the Australian pygmy copperhead (Austrelaps. labialis): An atypical long-chain α-neurotoxin with only weak affinity for α7 nicotinic receptors. Biochem. Pharmacol. 2012, 84, 851–863. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, Y.; Kano, M.; Tamiya, N.; Shimada, Y. Acetylcholine receptors of human skeletal muscle: A species difference detected by snake neurotoxins. Brain Res. 1985, 346, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Pawlak, J.; Mackessy, S.P.; Fry, B.G.; Bhatia, M.; Mourier, G.; Fruchart-Gaillard, C.; Servent, D.; Ménez, R.; Stura, E.; Ménez, A.; et al. Denmotoxin, a three-finger toxin from the colubrid snake Boiga dendrophila (Mangrove Catsnake) with bird-specific activity. J. Biol. Chem. 2006, 281, 29030–29041. [Google Scholar] [CrossRef] [PubMed]
- Heyborne, W.H.; Mackessy, S.P. Identification and characterization of a taxon-specific three-finger toxin from the venom of the Green Vinesnake (Oxybelis fulgidus; Family Colubridae). Biochimie 2013, 95, 1923–1932. [Google Scholar] [CrossRef] [PubMed]
- Osipov, A.V.; Kasheverov, I.E.; Makarova, Y.V.; Starkov, V.G.; Vorontsova, O.V.; Ziganshin, R.K.; Andreeva, T.V.; Serebryakova, M.V.; Benoit, A.; Hogg, R.C.; et al. Naturally occurring disulfide-bound dimers of three-fingered toxins: A paradigm for biological activity diversification. J. Biol. Chem. 2008, 283, 14571–14580. [Google Scholar] [CrossRef] [PubMed]
- Pawlak, J.; Mackessy, S.P.; Sixberry, N.M.; Stura, E.A.; Le Du, M.H.; Menez, R.; Foo, C.S.; Menez, A.; Nirthanan, S.; Kini, R.M. Irditoxin, a novel covalently linked heterodimeric three finger toxin with high taxon-specific neurotoxicity. FASEB J. 2009, 23, 534–545. [Google Scholar] [CrossRef] [PubMed]
- McCann, C.M.; Bracamontes, J.; Steinbach, J.H.; Sanes, J.R. The cholinergic antagonist alpha-bungarotoxin also binds and blocks a subset of GABA receptors. Proc. Natl. Acad. Sci. USA 2006, 103, 5149–5154. [Google Scholar] [CrossRef] [PubMed]
- Hannan, S.; Mortensen, M.; Smart, T.G. Snake neurotoxin α-bungarotoxin is an antagonist at native GABAA receptors. Neuropharmacology 2015, 93C, 28–40. [Google Scholar] [CrossRef] [PubMed]
- Miwa, J.M.; Freedman, R.; Lester, H.A. Neural systems governed by nicotinic acetylcholine receptors: Emerging hypotheses. Neuron 2011, 70, 20–33. [Google Scholar] [CrossRef] [PubMed]
- Ibañez-Tallon, I.; Nitabach, M.N. Tethering toxins and peptide ligands for modulation of neuronal function. Curr. Opin. Neurobiol. 2012, 22, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Miwa, J.M.; Lester, H.A.; Walz, A. Optimizing cholinergic tone through lynx modulators of nicotinic receptors: Implications for plasticity and nicotine addiction. Physiology (Bethesda) 2012, 27, 187–199. [Google Scholar] [CrossRef]
- Lyukmanova, E.N.; Shulepko, M.A.; Buldakova, S.L.; Kasheverov, I.E.; Shenkarev, Z.O.; Reshetnikov, R.V.; Filkin, S.Y.; Kudryavtsev, D.S.; Ojomoko, L.O.; Kryukova, E.V.; et al. Water-soluble LYNX1 residues important for interaction with muscle-type and/or neuronal nicotinic receptors. J. Biol. Chem. 2013, 288, 15888–15899. [Google Scholar] [CrossRef] [PubMed]
- Nichols, W.A.; Henderson, B.J.; Yu, C.; Parker, R.L.; Richards, C.I.; Lester, H.A.; Miwa, J.M. Lynx1 shifts α4β2 nicotinic receptor subunit stoichiometry by affecting assembly in the endoplasmic reticulum. J. Biol. Chem. 2014, 289, 31423–31432. [Google Scholar] [CrossRef] [PubMed]
- Puddifoot, C.A.; Wu, M.; Sung, R.-J.; Joiner, W.J. Ly6h regulates trafficking of alpha7 nicotinic acetylcholine receptors and nicotine-induced potentiation of glutamatergic signaling. J. Neurosci. 2015, 35, 3420–3430. [Google Scholar] [CrossRef] [PubMed]
- Nirthanan, S.; Gopalakrishnakone, P.; Gwee, M.C.E.; Khoo, H.E.; Kini, R.M. Non-conventional toxins from Elapid venoms. Toxicon 2003, 41, 397–407. [Google Scholar] [CrossRef] [PubMed]
- Mordvintsev, D.Y.; Polyak, Y.L.; Rodionov, D.I.; Jakubik, J.; Dolezal, V.; Karlsson, E.; Tsetlin, V.I.; Utkin, Y.N. Weak toxin WTX from Naja kaouthia cobra venom interacts with both nicotinic and muscarinic acetylcholine receptors. FEBS J. 2009, 276, 5065–5075. [Google Scholar] [CrossRef] [PubMed]
- Utkin, Y.N.; Kukhtina, V.V; Kryukova, E.V; Chiodini, F.; Bertrand, D.; Methfessel, C.; Tsetlin, V.I. “Weak toxin” from Naja kaouthia is a nontoxic antagonist of alpha 7 and muscle-type nicotinic acetylcholine receptors. J. Biol. Chem. 2001, 276, 15810–15815. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Robinson, J.E.; Joiner, W.J. SLEEPLESS is a bifunctional regulator of excitability and cholinergic synaptic transmission. Curr. Biol. 2014, 24, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Vulfius, C.A.; Gorbacheva, E.V.; Starkov, V.G.; Osipov, A.V.; Kasheverov, I.E.; Andreeva, T.V.; Astashev, M.E.; Tsetlin, V.I.; Utkin, Y.N. An unusual phospholipase A2 from puff adder Bitis arietans venom—A novel blocker of nicotinic acetylcholine receptors. Toxicon 2011, 57, 787–793. [Google Scholar] [CrossRef]
- Vulfius, C.A.; Kasheverov, I.E.; Starkov, V.G.; Osipov, A.V.; Andreeva, T.V.; Filkin, S.Y.; Gorbacheva, E.V.; Astashev, M.E.; Tsetlin, V.I.; Utkin, Y.N. Inhibition of nicotinic acetylcholine receptors, a novel facet in the pleiotropic activities of snake venom phospholipases A2. PLoS ONE 2014, 9, e115428. [Google Scholar] [CrossRef] [PubMed]
- Vulfius, C.A.; Starkov, V.G.; Andreeva, T.V.; Tsetlin, V.I.; Utkin, Yu.N. Novel antagonists of nicotinic acethylcholine receptors—Proteins from venoms of viperidae snakes. Dokl. Biochem. Biophys. 2015, 461, 604–607. [Google Scholar] [CrossRef]
- Bixel, M.G.; Weise, C.; Bolognesi, M.L.; Rosini, M.; Brierly, M.J.; Mellor, I.R.; Usherwood, P.N.R.; Melchiorre, C.; Hucho, F. Location of the polyamine binding site in the vestibule of the nicotinic acetylcholine receptor ion channel. J. Biol. Chem. 2000, 276, 6151–6160. [Google Scholar] [CrossRef] [PubMed]
- Pandhare, A.; Hamouda, A.K.; Staggs, B.; Aggarwal, S.; Duddempudi, P.K.; Lever, J.R.; Lapinsky, D.J.; Jansen, M.; Cohen, J.B.; Blanton, M.P. Bupropion binds to two sites in the Torpedo nicotinic acetylcholine receptor transmembrane domain: A photoaffinity labeling study with the bupropion analogue [(125)I]-SADU-3–72. Biochemistry 2012, 51, 2425–2435. [Google Scholar] [CrossRef] [PubMed]
- Collins, T.; Young, G.; Millar, N. Competitive binding at a nicotinic receptor transmembrane site of two α7-selective positive allosteric modulators with differing effects on agonist-evoked. Neuropharmacology 2011, 61, 1306–1313. [Google Scholar] [CrossRef] [PubMed]
- Pandya, A.A.; Yakel, J.L. Effects of neuronal nicotinic acetylcholine receptor allosteric modulators in animal behavior studies. Biochem. Pharmacol. 2013, 86, 1054–1062. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, Y.; Li, S.-T. The effect of local anesthetics on the inhibition of adult muscle-type nicotinic acetylcholine receptors by nondepolarizing muscle relaxants. Eur. J. Pharmacol. 2010, 630, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Alberola-Die, A.; Reboreda, A.; Lamas, J.A.; Morales, A. Lidocaine effects on acetylcholine-elicited currents from mouse superior cervical ganglion neurons. Neurosci. Res. 2013, 75, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Zuber, B.; Unwin, N. Structure and superorganization of acetylcholine receptor-rapsyn complexes. Proc. Natl. Acad. Sci. USA 2013, 110, 10622–10627. [Google Scholar] [CrossRef] [PubMed]
- Punga, A.R.; Maj, M.; Lin, S.; Meinen, S.; Rüegg, M.A. MuSK levels differ between adult skeletal muscles and influence postsynaptic plasticity. Eur. J. Neurosci. 2011, 33, 890–898. [Google Scholar] [CrossRef] [PubMed]
- Stiegler, A.L.; Burden, S.J.; Hubbard, S.R. Crystal structure of the frizzled-like cysteine-rich domain of the receptor tyrosine kinase MuSK. J. Mol. Biol. 2009, 393, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Akdemir, A.; Edink, E.; Thompson, A.J.; Lummis, S.C.R.; Kooistra, A.J.; de Graaf, C.; de Esch, I.J.P. Identification of novel α7 nicotinic receptor ligands by in silico screening against the crystal structure of a chimeric α7 receptor ligand binding domain. Bioorg. Med. Chem. 2012, 20, 5992–6002. [Google Scholar] [CrossRef] [PubMed]
- Hilf, R.J.C.; Dutzler, R. X-ray structure of a prokaryotic pentameric ligand-gated ion channel. Nature 2008, 452, 375–379. [Google Scholar] [CrossRef] [PubMed]
- Bocquet, N.; Nury, H.; Baaden, M.; le Poupon, C.; Changeux, J.-P.; Delarue, M.; Corringer, P.-J. X-ray structure of a pentameric ligand-gated ion channel in an apparently open conformation. Nature 2009, 457, 111–114. [Google Scholar] [CrossRef] [PubMed]
- Nury, H.; van Renterghem, C.; Weng, Y.; Tran, A.; Baaden, M.; Dufresne, V.; Changeux, J.-P.; Sonner, J.M.; Delarue, M.; Corringer, P.-J. X-ray structures of general anaesthetics bound to a pentameric ligand-gated ion channel. Nature 2011, 469, 428–431. [Google Scholar] [CrossRef] [PubMed]
- Hibbs, R.E.; Gouaux, E. Principles of activation and permeation in an anion-selective Cys-loop receptor. Nature 2011, 474, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Hassaine, G.; Deluz, C.; Grasso, L.; Wyss, R.; Tol, M.B.; Hovius, R.; Graff, A.; Stahlberg, H.; Tomizaki, T.; Desmyter, A.; et al. X-ray structure of the mouse serotonin 5-HT3 receptor. Nature 2014, 512, 276–281. [Google Scholar] [CrossRef] [PubMed]
- Miller, P.S.; Aricescu, A.R. Crystal structure of a human GABAA receptor. Nature 2014, 512, 270–275. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kudryavtsev, D.; Shelukhina, I.; Vulfius, C.; Makarieva, T.; Stonik, V.; Zhmak, M.; Ivanov, I.; Kasheverov, I.; Utkin, Y.; Tsetlin, V. Natural Compounds Interacting with Nicotinic Acetylcholine Receptors: From Low-Molecular Weight Ones to Peptides and Proteins. Toxins 2015, 7, 1683-1701. https://doi.org/10.3390/toxins7051683
Kudryavtsev D, Shelukhina I, Vulfius C, Makarieva T, Stonik V, Zhmak M, Ivanov I, Kasheverov I, Utkin Y, Tsetlin V. Natural Compounds Interacting with Nicotinic Acetylcholine Receptors: From Low-Molecular Weight Ones to Peptides and Proteins. Toxins. 2015; 7(5):1683-1701. https://doi.org/10.3390/toxins7051683
Chicago/Turabian StyleKudryavtsev, Denis, Irina Shelukhina, Catherine Vulfius, Tatyana Makarieva, Valentin Stonik, Maxim Zhmak, Igor Ivanov, Igor Kasheverov, Yuri Utkin, and Victor Tsetlin. 2015. "Natural Compounds Interacting with Nicotinic Acetylcholine Receptors: From Low-Molecular Weight Ones to Peptides and Proteins" Toxins 7, no. 5: 1683-1701. https://doi.org/10.3390/toxins7051683
APA StyleKudryavtsev, D., Shelukhina, I., Vulfius, C., Makarieva, T., Stonik, V., Zhmak, M., Ivanov, I., Kasheverov, I., Utkin, Y., & Tsetlin, V. (2015). Natural Compounds Interacting with Nicotinic Acetylcholine Receptors: From Low-Molecular Weight Ones to Peptides and Proteins. Toxins, 7(5), 1683-1701. https://doi.org/10.3390/toxins7051683