Short Toxin-like Proteins Attack the Defense Line of Innate Immunity



Abstract
:1. Introduction
2. Results and Discussion
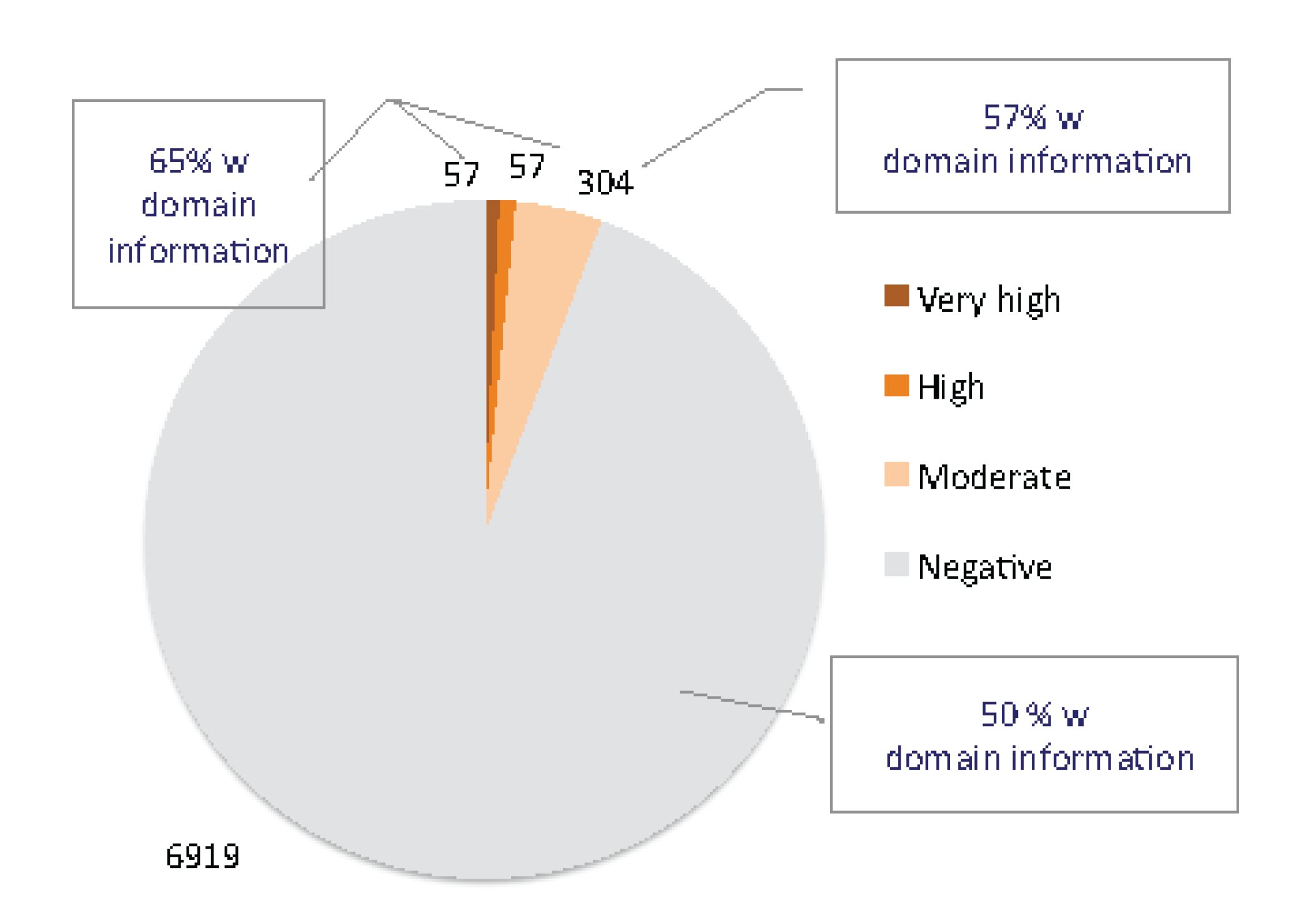
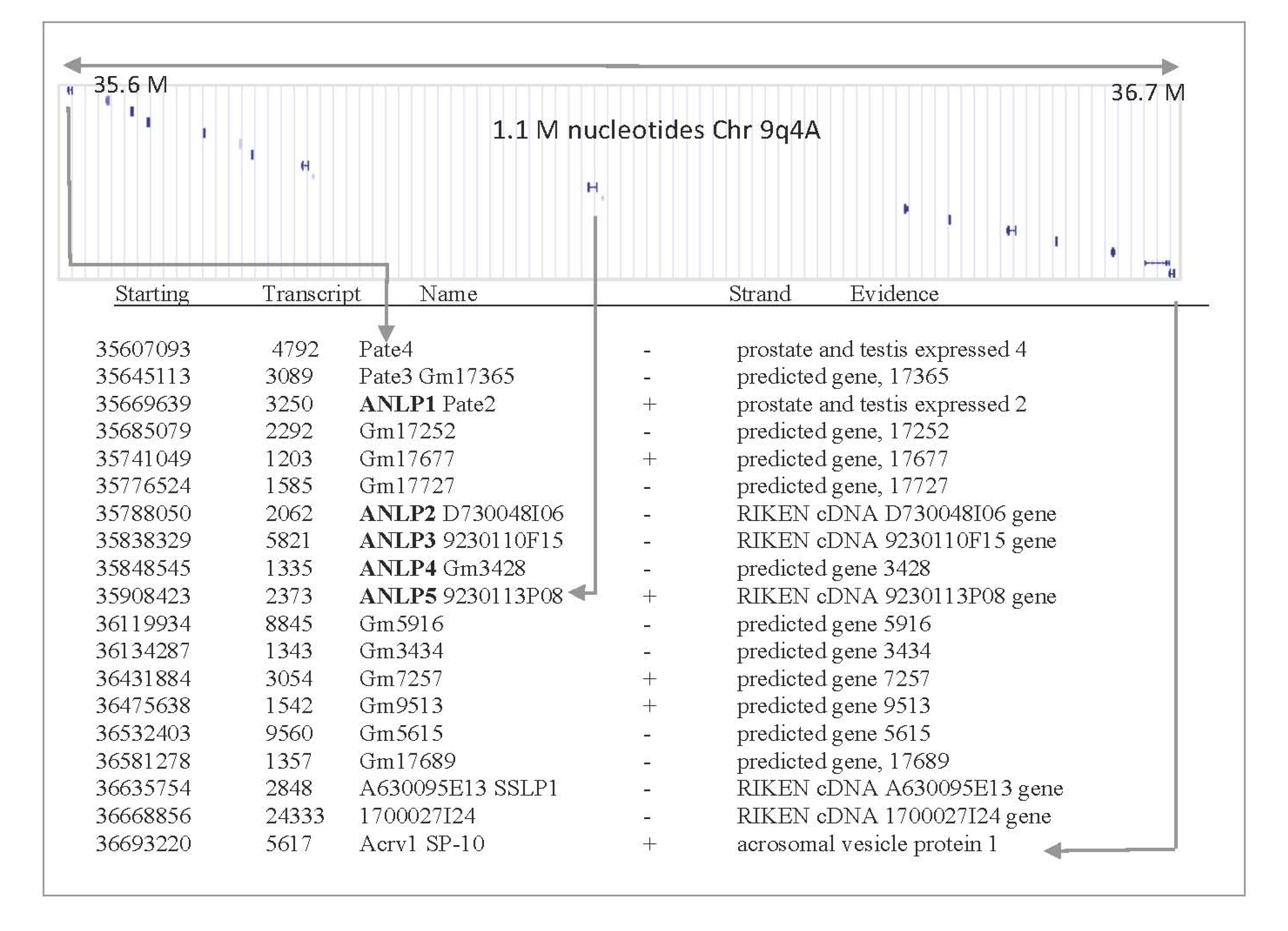
2.1. Discovering Natural Endogenous α-Neurotoxins in Mouse
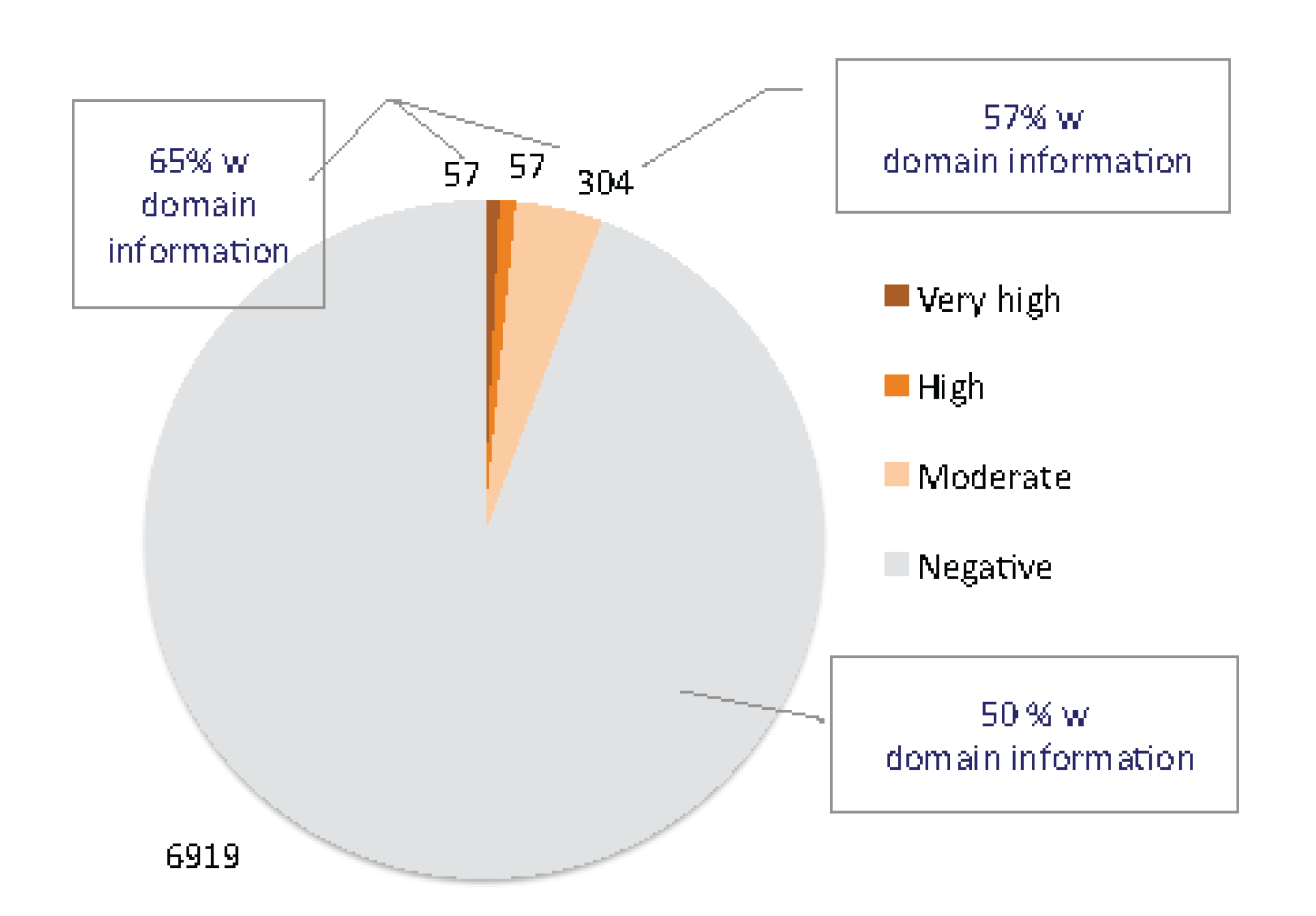
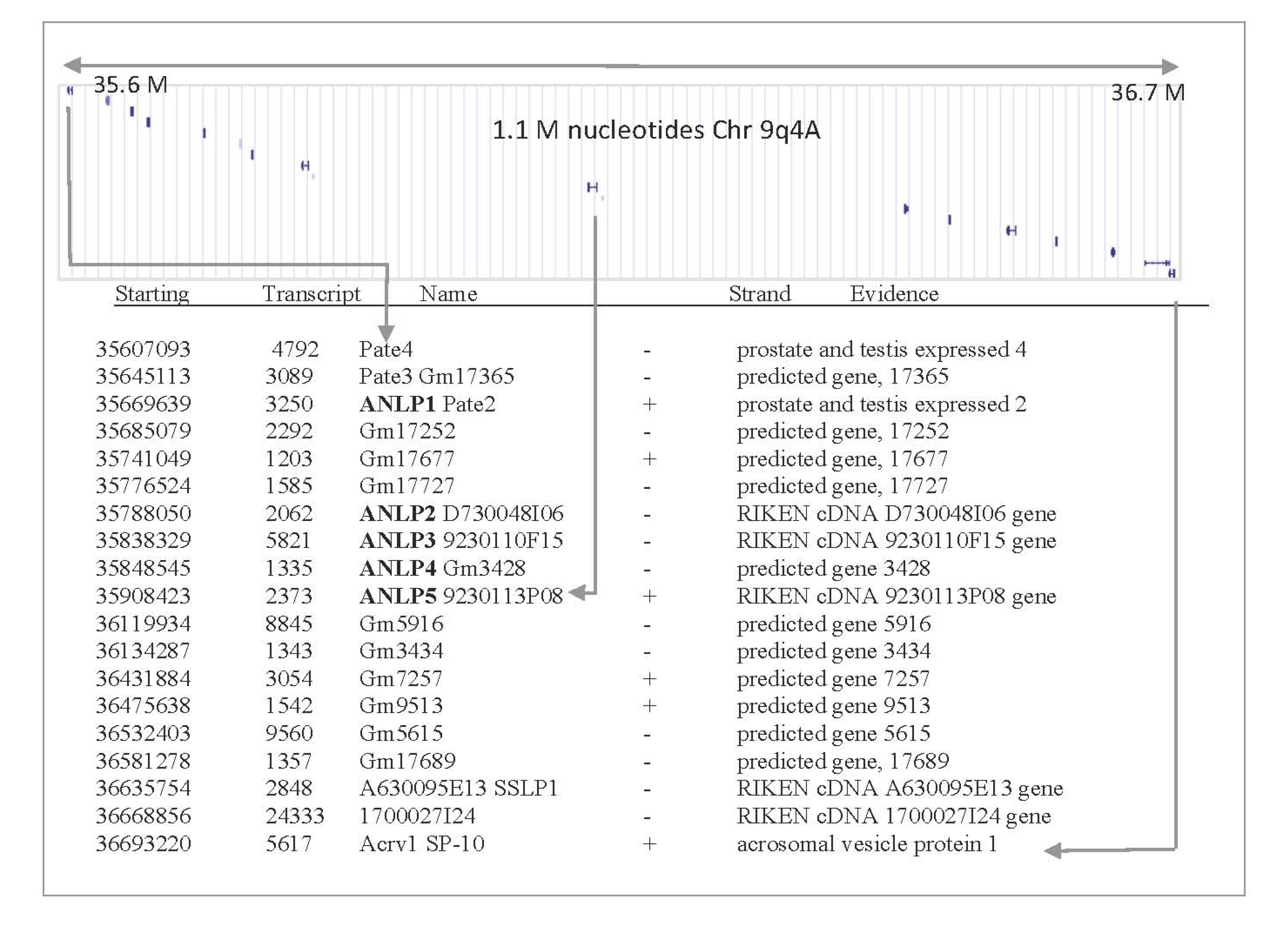
2.2. ANLPs Belong to an Expanded Gene Family with a Single Ly6/uPAR Domain
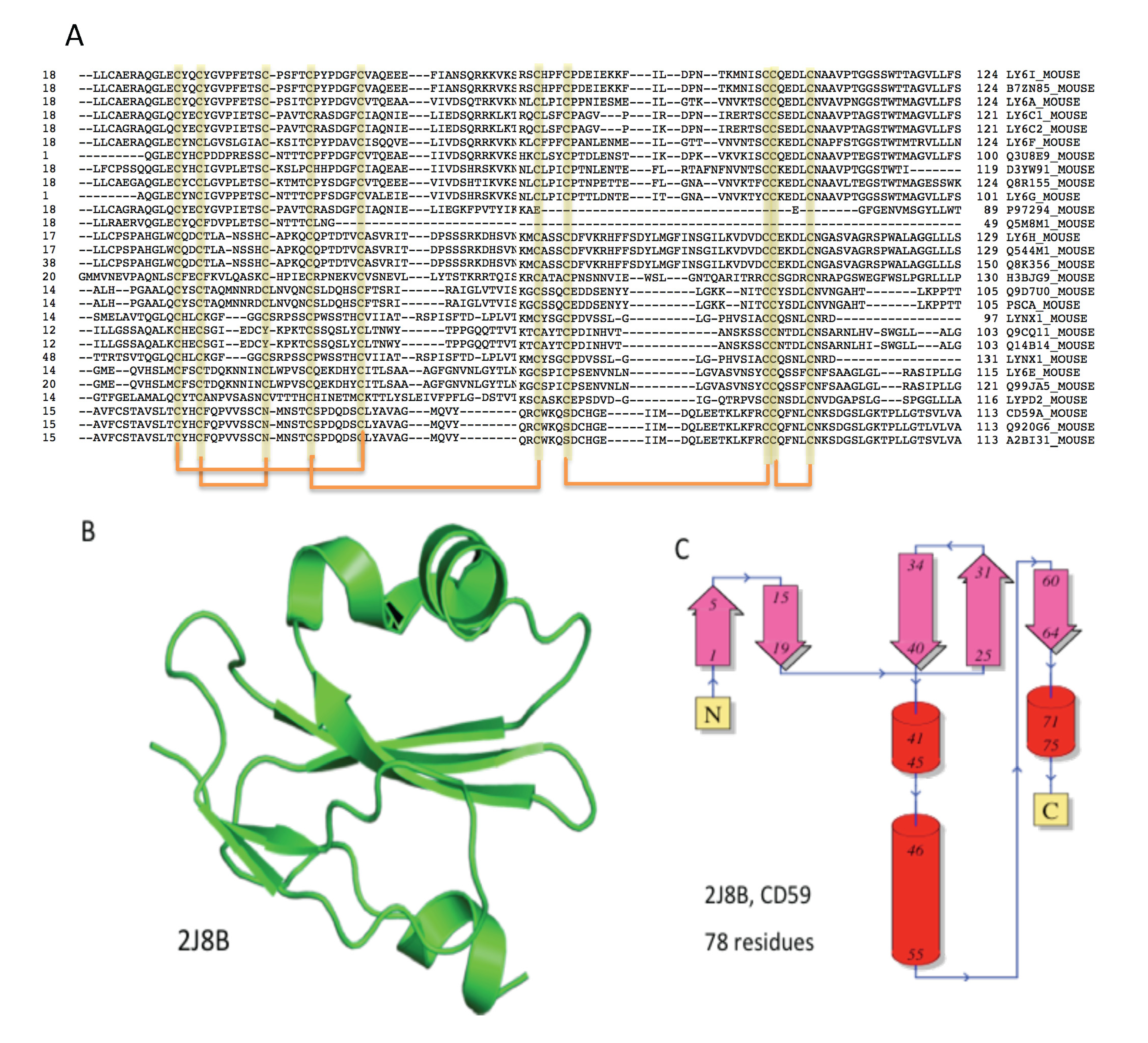
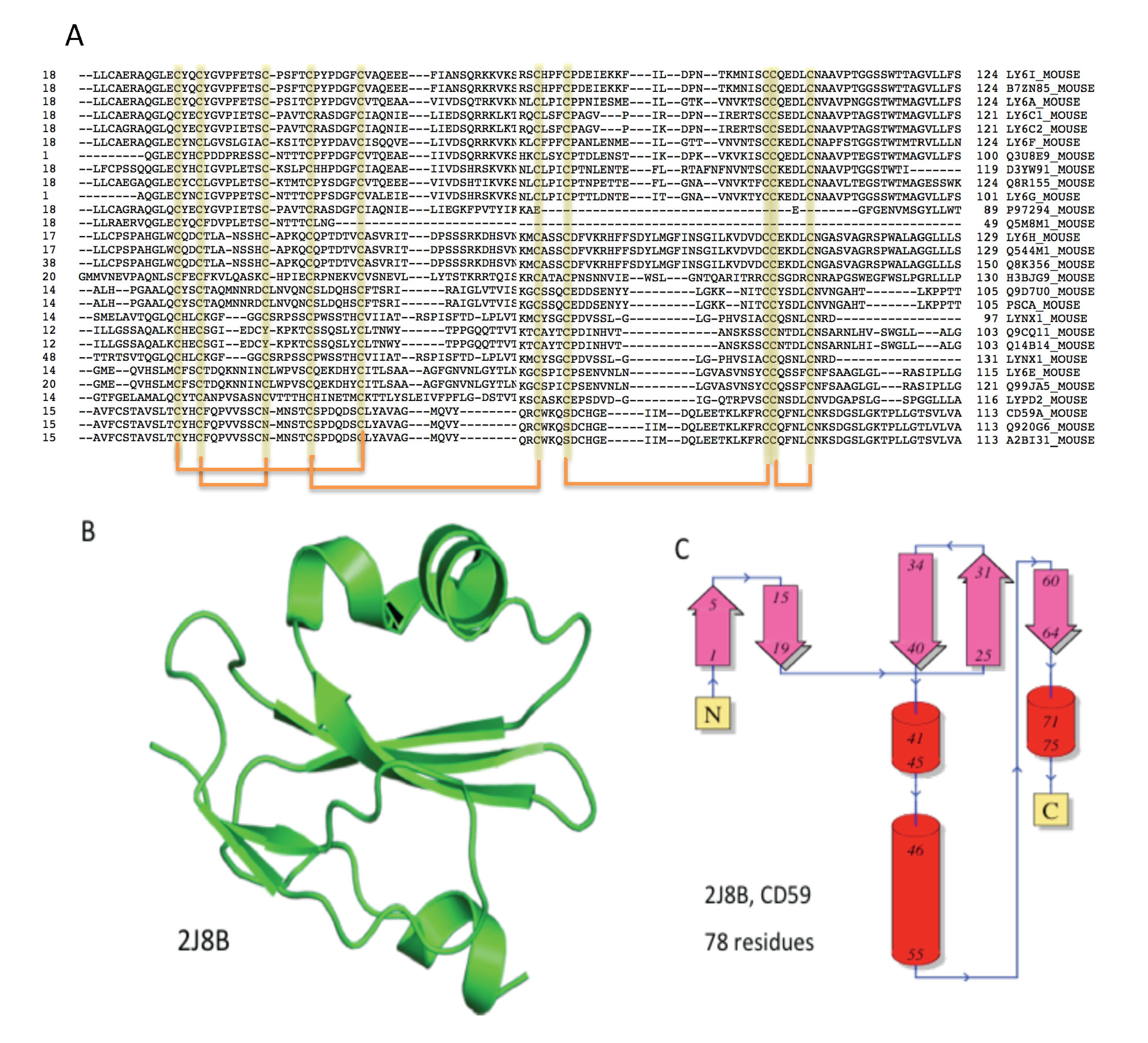
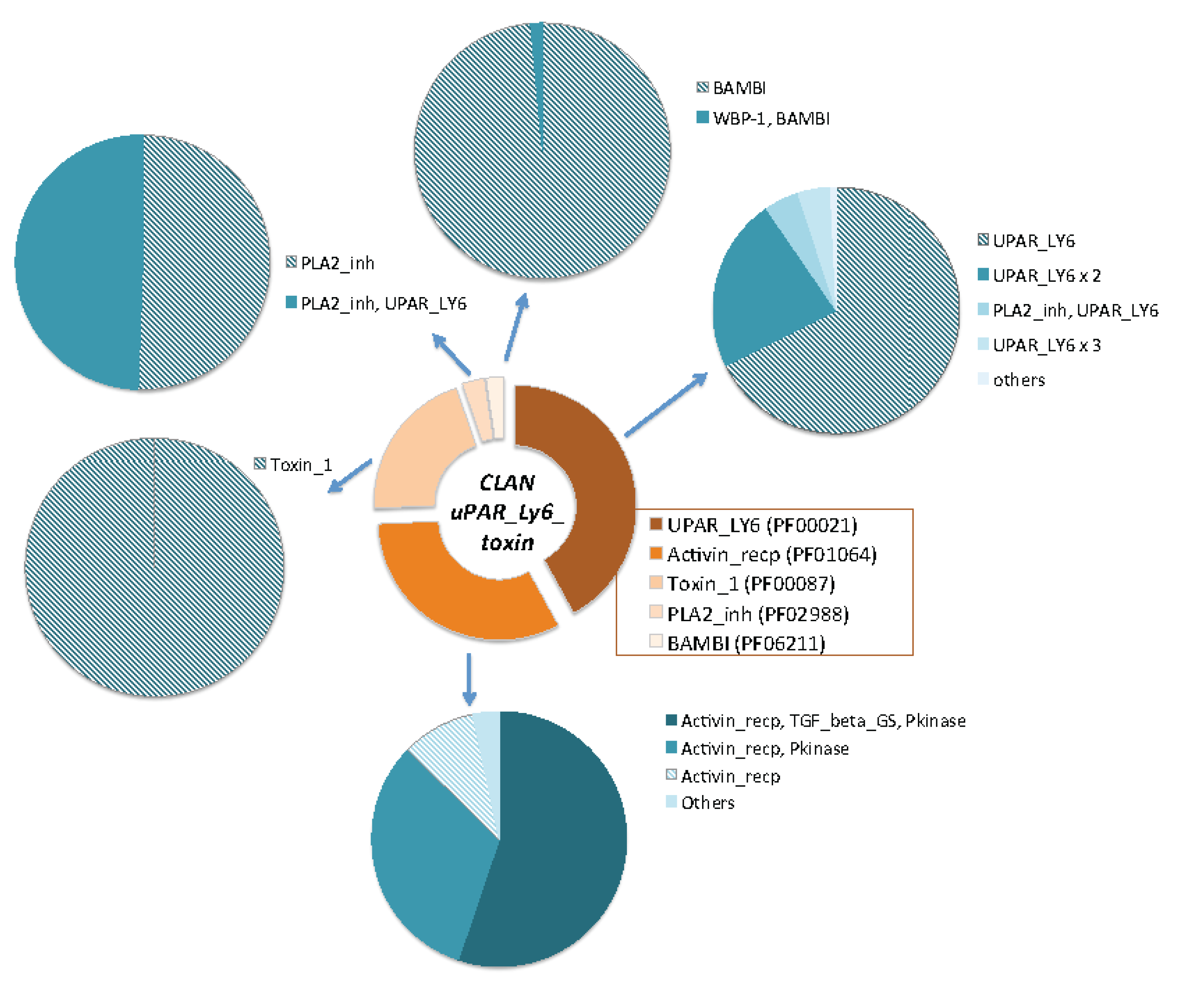
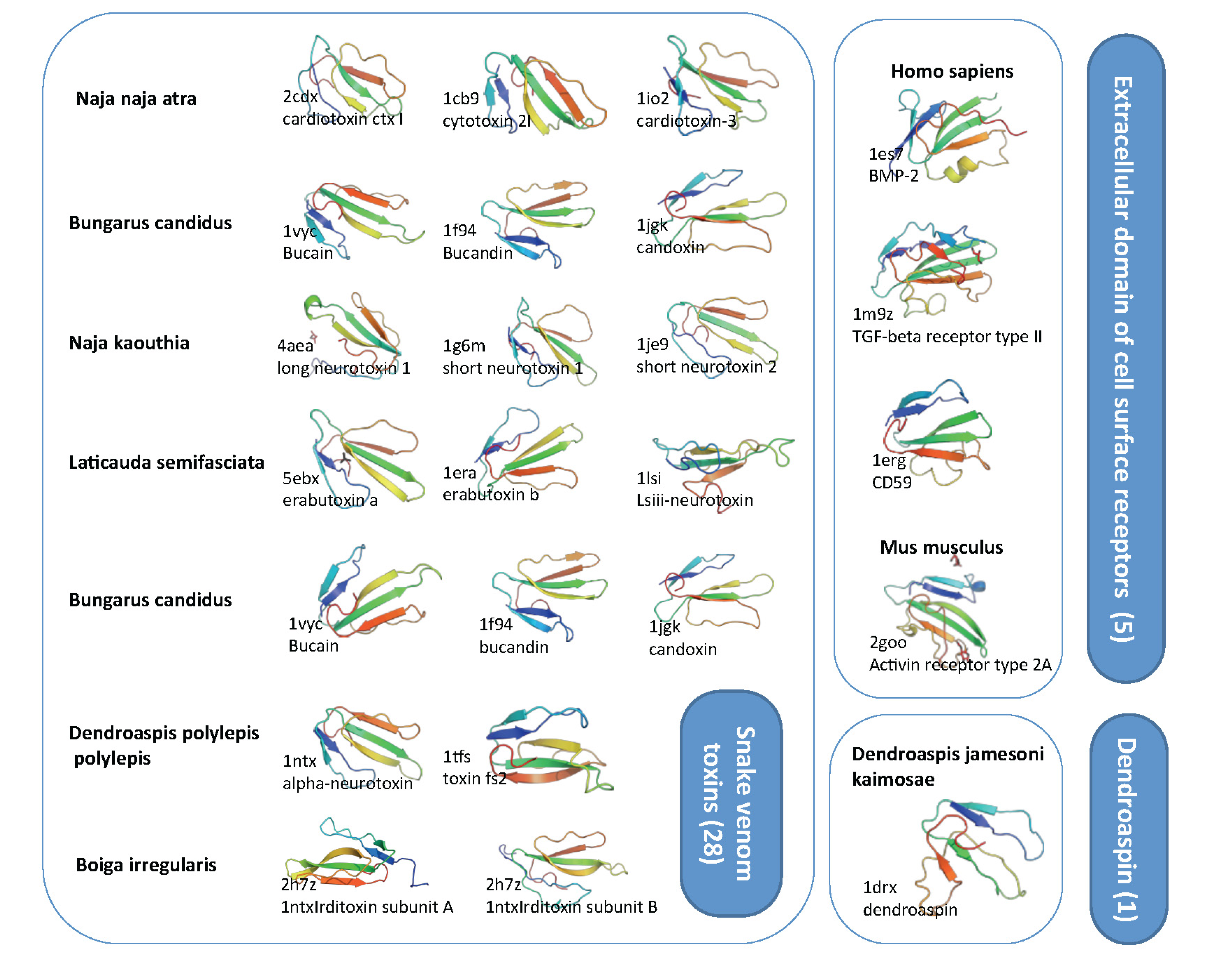
2.3. Domain Composition and Structural View on Snake Toxins and TOLIPs
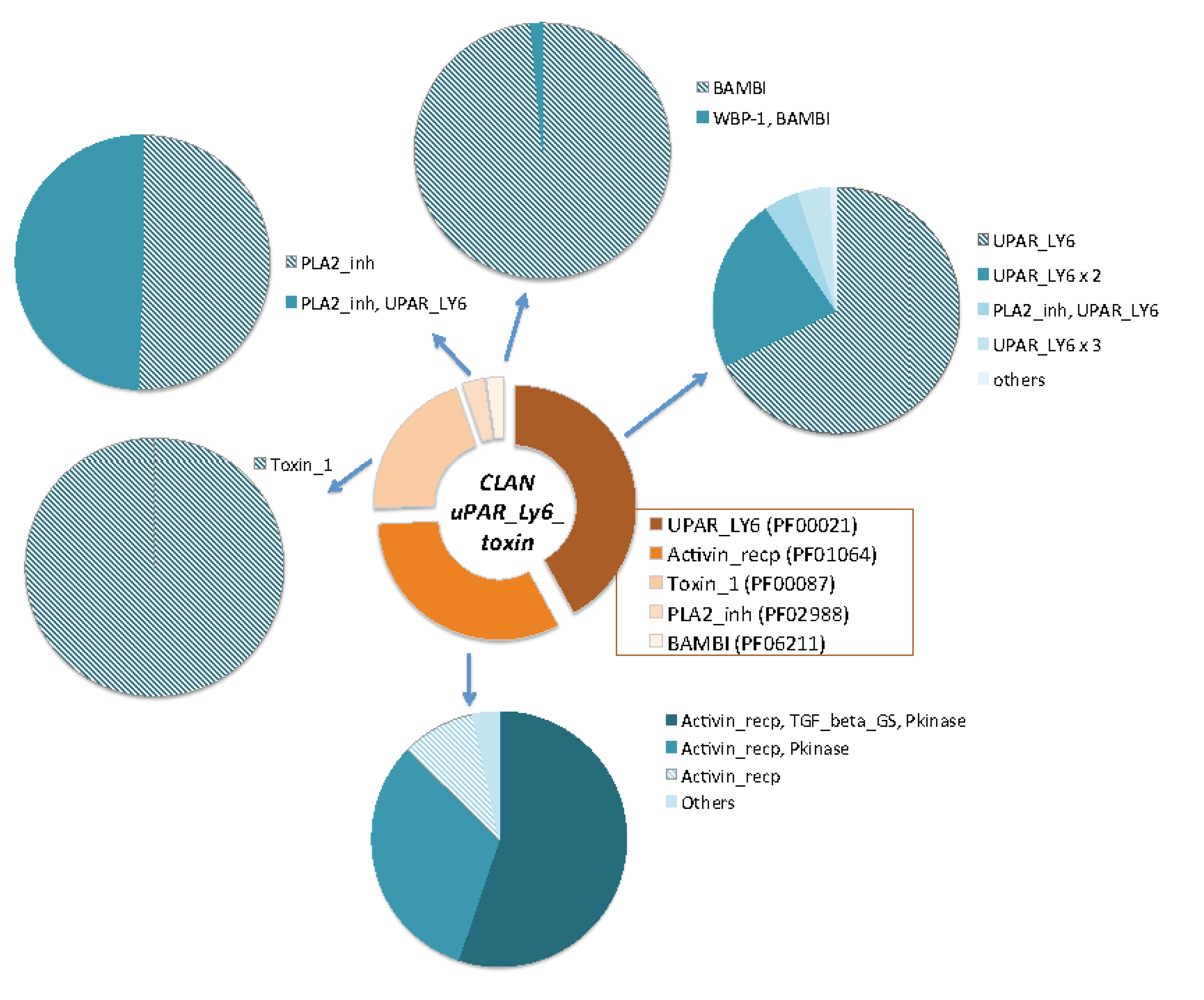

2.4. Ample Binding Specificities for the Snake-Toxin-like Fold

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene symbol | Biological process | Reference |
|---|---|---|
| CD177 | Neutrophil proliferation, inflammatory settings | [34] |
| CD59 | Regulating the action of the complement MAC | [35] |
| GPI-HBP | Lipid uptake from high-density lipoprotein (HDL) particles | [36] |
| Intectin | Intestinal epithelial apoptosis | [37] |
| Ly-6A | Mediating cell-cell adhesion | [38] |
| Ly-6C | Regulating adhesion and homing of T-cells | [39] |
| Lynx-1 | Neuronal survival and Apoptosis | [40] |
| Lynx2 | Potentate axon outgrowth and guidance | [41] |
| PSCA | Neuronal development and survival | [42] |
| SAMP-14 | Sperm-egg interactions | [43] |
| SLURP-1 | Implicated in a skin disorder, Mal de Meleda | [44] |
| SLURP-2 | Induced in psoriasis vulgaris | [19] |
2.5. TOLIPs Alter the Signaling Pathways of the Immune System
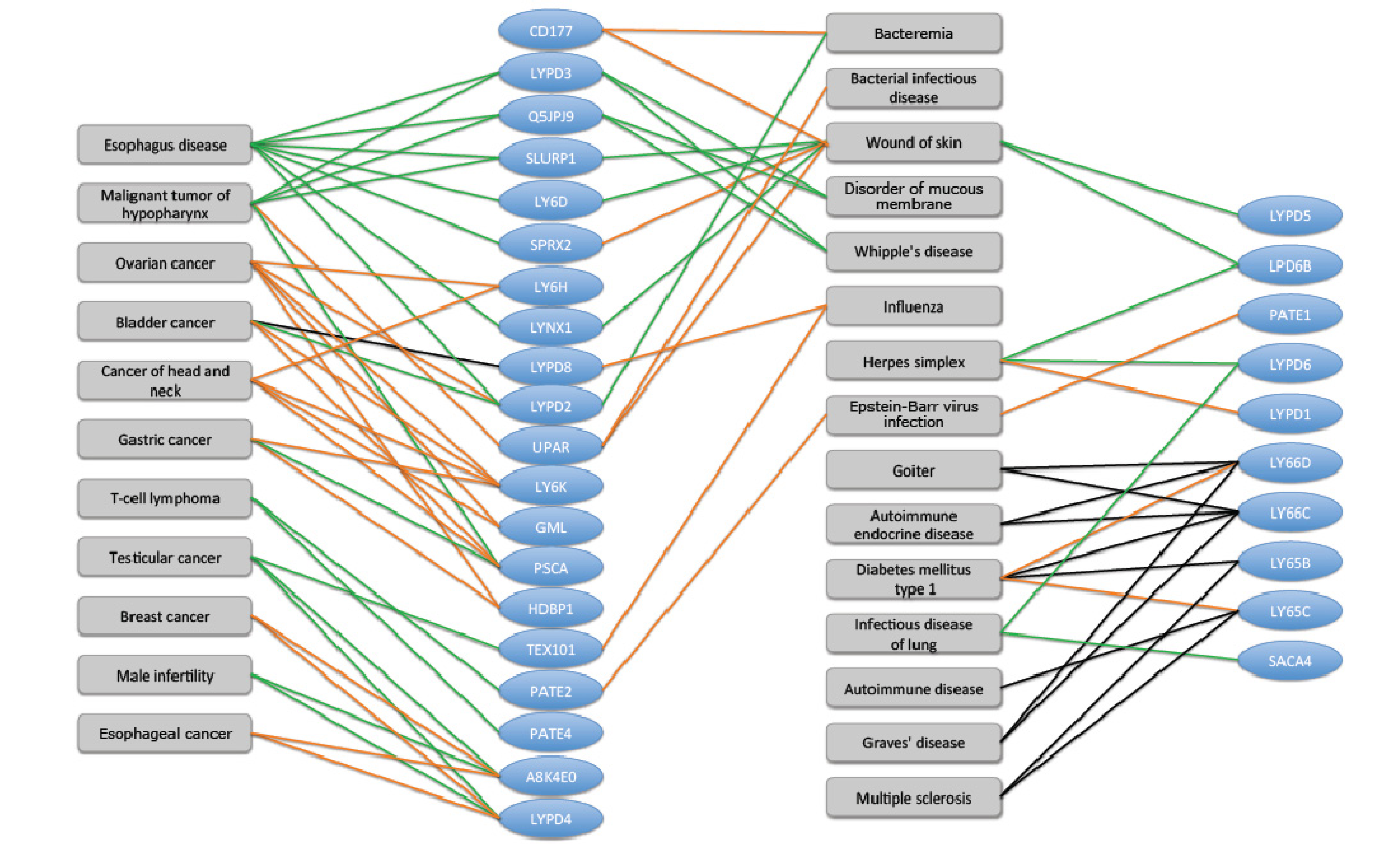
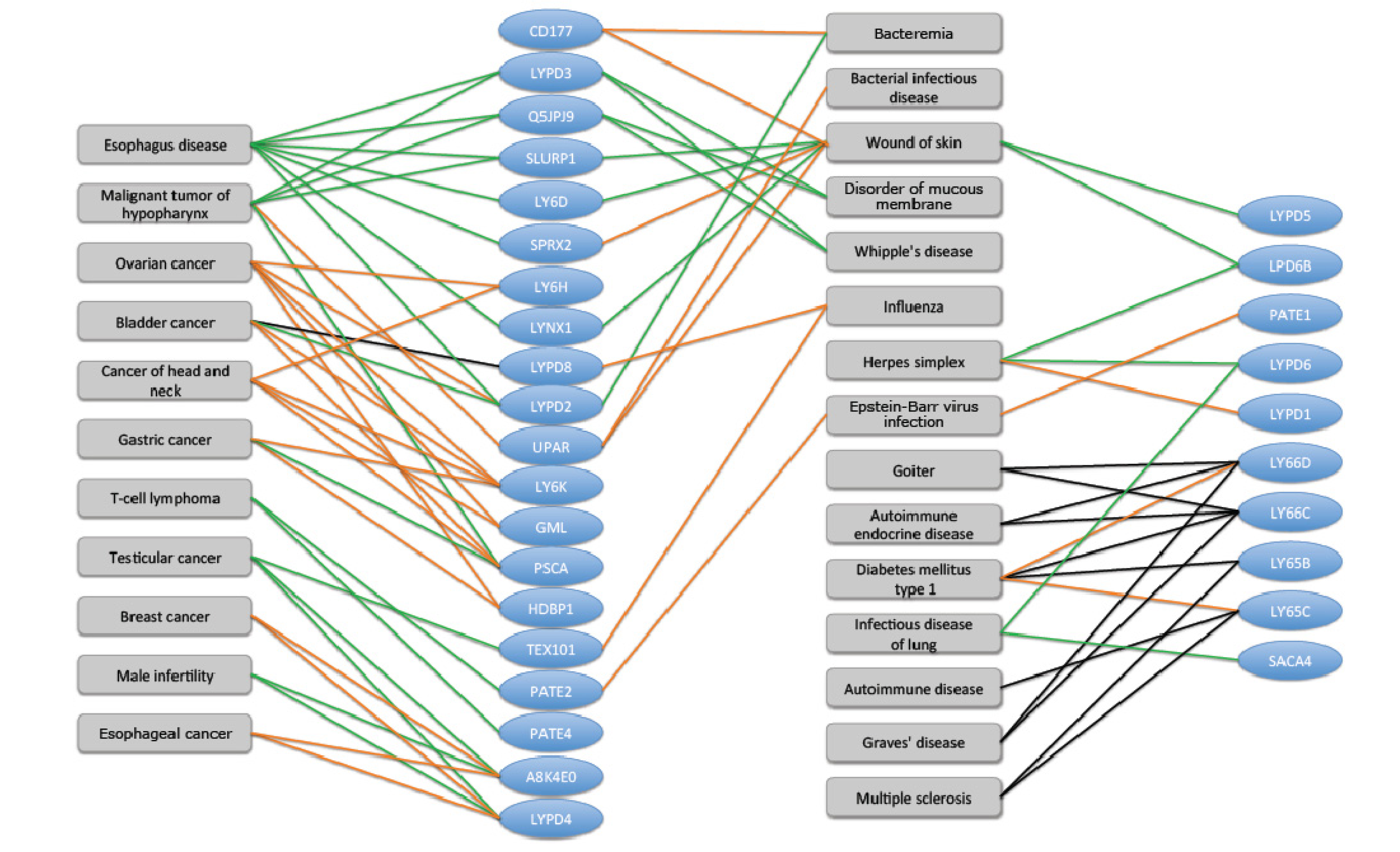
2.6. TOLIPs Linkage to Pathologies and Diseases
3. Experimental Section
3.1. Data Collection
3.2. ClanTox Prediction
3.3. Bioinformatics Analysis Tools
4. Conclusions
Abbreviations
| ADHD | attention deficit hyperactivity disorder |
| ANLP | α-neurotoxin-like protein |
| ClanTox | classifier of animal toxins |
| GPI | glycosylphosphatidylinositol |
| GWAS | genome-wide association studies |
| LPS | lipopolysaccharide |
| MAC | membrane attack complex |
| MS | mass spectrometry |
| nAChR | nicotinic acetylcholine receptor |
| OCLP | omega conotoxin-like protein |
| PSCA | prostate stem cell antigen |
| SP | signal peptide |
| TFP | three-finger proteins |
| TNF | tumor necrosis factor |
| TOLIP | toxin-like proteins |
| uPAR | urokinase-type plasminogen activator |
Acknowledgements
Conflict of Interest
References
- Loewenstein, Y.; Raimondo, D.; Redfern, O.C.; Watson, J.; Frishman, D.; Linial, M.; Orengo, C.; Thornton, J.; Tramontano, A. Protein function annotation by homology-based inference. Genome Biol. 2009, 10, 207. [Google Scholar] [CrossRef]
- Frith, M.C.; Forrest, A.R.; Nourbakhsh, E.; Pang, K.C.; Kai, C.; Kawai, J.; Carninci, P.; Hayashizaki, Y.; Bailey, T.L.; Grimmond, S.M. The abundance of short proteins in the mammalian proteome. PLoS Genet. 2006, 2, e52. [Google Scholar] [CrossRef]
- Clark, M.B.; Johnston, R.L.; Inostroza-Ponta, M.; Fox, A.H.; Fortini, E.; Moscato, P.; Dinger, M.E.; Mattick, J.S. Genome-wide analysis of long noncoding RNA stability. Genome Res. 2012, 22, 885–898. [Google Scholar] [CrossRef]
- Fry, B.G.; Roelants, K.; Champagne, D.E.; Scheib, H.; Tyndall, J.D.; King, G.F.; Nevalainen, T.J.; Norman, J.A.; Lewis, R.J.; Norton, R.S.; Renjifo, C.; de la Vega, R.C. The toxicogenomic multiverse: Convergent recruitment of proteins into animal venoms. Annu. Rev. Genomics Hum. Genet. 2009, 10, 483–511. [Google Scholar] [CrossRef]
- Naamati, G.; Askenazi, M.; Linial, M. A predictor for toxin-like proteins exposes cell modulator candidates within viral genomes. Bioinformatics 2010, 26, i482–i488. [Google Scholar] [CrossRef]
- Kaplan, N.; Morpurgo, N.; Linial, M. Novel families of toxin-like peptides in insects and mammals: a computational approach. J. Mol. Biol. 2007, 369, 553–566. [Google Scholar] [CrossRef]
- Whittington, C.M.; Papenfuss, A.T.; Bansal, P.; Torres, A.M.; Wong, E.S.; Deakin, J.E.; Graves, T.; Alsop, A.; Schatzkamer, K.; Kremitzki, C.; et al. Defensins and the convergent evolution of platypus and reptile venom genes. Genome Res. 2008, 18, 986–994. [Google Scholar] [CrossRef]
- Bachar-Wikstrom, E.; Wikstrom, J.D.; Ariav, Y.; Tirosh, B.; Kaiser, N.; Cerasi, E.; Leibowitz, G. Stimulation of autophagy improves endoplasmic reticulum stress-induced diabetes. Diabetes 2013, 62, 1227–1237. [Google Scholar] [CrossRef]
- Tsetlin, V. Snake venom alpha-neurotoxins and other “three-finger” proteins. Eur. J. Biochem. 1999, 264, 281–286. [Google Scholar] [CrossRef]
- Walkinshaw, M.D.; Saenger, W.; Maelicke, A. Three-dimensional structure of the “long” neurotoxin from cobra venom. Proc. Natl. Acad. Sci. USA 1980, 77, 2400–2404. [Google Scholar] [CrossRef]
- Tsernoglou, D.; Petsko, G.A.; Hudson, R.A. Structure and function of snake venom curarimimetic neurotoxins. Mol. Pharmacol. 1978, 14, 710–716. [Google Scholar]
- Kini, R.M. Molecular moulds with multiple missions: functional sites in three-finger toxins. Clin. Exp. Pharmacol. Physiol. 2002, 29, 815–822. [Google Scholar] [CrossRef]
- Koua, D.; Brauer, A.; Laht, S.; Kaplinski, L.; Favreau, P.; Remm, M.; Lisacek, F.; Stocklin, R. ConoDictor: A tool for prediction of conopeptide superfamilies. Nucleic Acids Res. 2012, 40, W238–W241. [Google Scholar]
- Lenffer, J.; Lai, P.; El Mejaber, W.; Khan, A.M.; Koh, J.L.; Tan, P.T.; Seah, S.H.; Brusic, V. CysView: Protein classification based on cysteine pairing patterns. Nucleic Acids Res. 2004, 32, W350–W355. [Google Scholar] [CrossRef]
- Kaas, Q.; Westermann, J.C.; Halai, R.; Wang, C.K.; Craik, D.J. ConoServer, a database for conopeptide sequences and structures. Bioinformatics 2008, 24, 445–446. [Google Scholar] [CrossRef]
- Fry, B.G. From genome to “venome”: Molecular origin and evolution of the snake venom proteome inferred from phylogenetic analysis of toxin sequences and related body proteins. Genome Res. 2005, 15, 403–420. [Google Scholar] [CrossRef]
- Fry, B.G.; Vidal, N.; Norman, J.A.; Vonk, F.J.; Scheib, H.; Ryan Ramjan, S.F.; Kuruppu, S.; Fung, K.; Blair Hedges, S.; Richardson, M.K.; et al. Early evolution of the venom system in lizards and snakes. Nature 2006, 439, 584–588. [Google Scholar] [CrossRef]
- Miwa, J.M.; Ibanez-Tallon, I.; Crabtree, G.W.; Sanchez, R.; Sali, A.; Role, L.W.; Heintz, N. lynx1, an endogenous toxin-like modulator of nicotinic acetylcholine receptors in the mammalian CNS. Neuron 1999, 23, 105–114. [Google Scholar] [CrossRef]
- Tjiu, J.W.; Lin, P.J.; Wu, W.H.; Cheng, Y.P.; Chiu, H.C.; Thong, H.Y.; Chiang, B.L.; Yang, W.S.; Jee, S.H. SLURP1 mutation-impaired T-cell activation in a family with Mal de Meleda. Br. J. Dermatol. 2011, 164, 47–53. [Google Scholar] [CrossRef]
- Tirosh, Y.; Morpurgo, N.; Cohen, M.; Linial, M.; Bloch, G. Raalin, a transcript enriched in the honey bee brain, is a remnant of genomic rearrangement in Hymenoptera. Insect. Mol. Biol. 2012, 21, 305–318. [Google Scholar] [CrossRef]
- Wright, R.M.; John, E.; Klotz, K.; Flickinger, C.J.; Herr, J.C. Cloning and sequencing of cDNAs coding for the human intra-acrosomal antigen SP-10. Biol. Reprod. 1990, 42, 693–701. [Google Scholar] [CrossRef]
- Anderson, D.J.; Johnson, P.M.; Alexander, N.J.; Jones, W.R.; Griffin, P.D. Monoclonal antibodies to human trophoblast and sperm antigens: Report of two WHO-sponsored workshops, June 30, 1986—Toronto, Canada. J. Reprod. Immunol. 1987, 10, 231–257. [Google Scholar] [CrossRef]
- Naamati, G.; Askenazi, M.; Linial, M. ClanTox: A classifier of short animal toxins. Nucleic Acids Res. 2009, 37, W363–W368. [Google Scholar] [CrossRef]
- Buczek, O.; Bulaj, G.; Olivera, B.M. Conotoxins and the posttranslational modification of secreted gene products. Cell. Mol. Life Sci. 2005, 62, 3067–3079. [Google Scholar] [CrossRef]
- Mulder, N.; Apweiler, R. InterPro and InterProScan: Tools for protein sequence classification and comparison. Methods Mol. Biol. 2007, 396, 59–70. [Google Scholar] [CrossRef]
- Bono, H.; Kasukawa, T.; Furuno, M.; Hayashizaki, Y.; Okazaki, Y. FANTOM DB: Database of Functional Annotation of RIKEN Mouse cDNA Clones. Nucleic Acids Res. 2002, 30, 116–118. [Google Scholar] [CrossRef]
- Finn, R.D.; Tate, J.; Mistry, J.; Coggill, P.C.; Sammut, S.J.; Hotz, H.R.; Ceric, G.; Forslund, K.; Eddy, S.R.; Sonnhammer, E.L.; Bateman, A. The Pfam protein families database. Nucleic Acids Res. 2008, 36, D281–D288. [Google Scholar] [CrossRef]
- Fortes-Dias, C.L.; Jannotti, M.L.; Franco, F.J.; Magalhaes, A.; Diniz, C.R. Studies on the specificity of CNF, a phospholipase A2 inhibitor isolated from the blood plasma of the South American rattlesnake (Crotalus durissus terrificus). I. Interaction with PLA2 from Lachesis muta muta snake venom. Toxicon 1999, 37, 1747–1759. [Google Scholar] [CrossRef]
- Csaba, G.; Birzele, F.; Zimmer, R. Systematic comparison of SCOP and CATH: A new gold standard for protein structure analysis. BMC Struct. Biol. 2009, 9, 23. [Google Scholar] [CrossRef]
- Adermann, K.; Wattler, F.; Wattler, S.; Heine, G.; Meyer, M.; Forssmann, W.G.; Nehls, M. Structural and phylogenetic characterization of human SLURP-1, the first secreted mammalian member of the Ly-6/uPAR protein superfamily. Protein Sci. 1999, 8, 810–819. [Google Scholar]
- Ploug, M.; Ellis, V. Structure-function relationships in the receptor for urokinase-type plasminogen activator. Comparison to other members of the Ly-6 family and snake venom alpha-neurotoxins. FEBS Lett. 1994, 349, 163–168. [Google Scholar] [CrossRef]
- Woody, J.N. Ly-6 is a T-cell differentiation antigen. Nature 1977, 269, 61–63. [Google Scholar] [CrossRef]
- Spindler, K.R.; Welton, A.R.; Lim, E.S.; Duvvuru, S.; Althaus, I.W.; Imperiale, J.E.; Daoud, A.I.; Chesler, E.J. The major locus for mouse adenovirus susceptibility maps to genes of the hematopoietic cell surface-expressed LY6 family. J. Immunol. 2010, 184, 3055–3062. [Google Scholar] [CrossRef]
- Sachs, U.J.; Andrei-Selmer, C.L.; Maniar, A.; Weiss, T.; Paddock, C.; Orlova, V.V.; Choi, E.Y.; Newman, P.J.; Preissner, K.T.; Chavakis, T.; Santoso, S. The neutrophil-specific antigen CD177 is a counter-receptor for platelet endothelial cell adhesion molecule-1 (CD31). J. Biol. Chem. 2007, 282, 23603–23612. [Google Scholar] [CrossRef]
- Davies, A.; Simmons, D.L.; Hale, G.; Harrison, R.A.; Tighe, H.; Lachmann, P.J.; Waldmann, H. CD59, an LY-6-like protein expressed in human lymphoid cells, regulates the action of the complement membrane attack complex on homologous cells. J. Exp. Med. 1989, 170, 637–654. [Google Scholar] [CrossRef]
- Ioka, R.X.; Kang, M.J.; Kamiyama, S.; Kim, D.H.; Magoori, K.; Kamataki, A.; Ito, Y.; Takei, Y.A.; Sasaki, M.; Suzuki, T.; et al. Expression cloning and characterization of a novel glycosylphosphatidylinositol-anchored high density lipoprotein-binding protein, GPI-HBP1. J. Biol. Chem. 2003, 278, 7344–7349. [Google Scholar] [CrossRef]
- Kitazawa, H.; Nishihara, T.; Nambu, T.; Nishizawa, H.; Iwaki, M.; Fukuhara, A.; Kitamura, T.; Matsuda, M.; Shimomura, I. Intectin, a novel small intestine-specific glycosylphosphatidylinositol-anchored protein, accelerates apoptosis of intestinal epithelial cells. J. Biol. Chem. 2004, 279, 42867–42874. [Google Scholar] [CrossRef]
- Bamezai, A.; Rock, K.L. Overexpressed Ly-6A.2 mediates cell-cell adhesion by binding a ligand expressed on lymphoid cells. Proc. Natl. Acad. Sci. USA 1995, 92, 4294–4298. [Google Scholar] [CrossRef]
- Jutila, M.A.; Kroese, F.G.; Jutila, K.L.; Stall, A.M.; Fiering, S.; Herzenberg, L.A.; Berg, E.L.; Butcher, E.C. Ly-6C is a monocyte/macrophage and endothelial cell differentiation antigen regulated by interferon-gamma. Eur. J. Immunol. 1988, 18, 1819–1826. [Google Scholar] [CrossRef]
- Miwa, J.M.; Stevens, T.R.; King, S.L.; Caldarone, B.J.; Ibanez-Tallon, I.; Xiao, C.; Fitzsimonds, R.M.; Pavlides, C.; Lester, H.A.; Picciotto, M.R.; Heintz, N. The prototoxin lynx1 acts on nicotinic acetylcholine receptors to balance neuronal activity and survival in vivo. Neuron 2006, 51, 587–600. [Google Scholar] [CrossRef]
- Dessaud, E.; Salaun, D.; Gayet, O.; Chabbert, M.; deLapeyriere, O. Identification of lynx2, a novel member of the ly-6/neurotoxin superfamily, expressed in neuronal subpopulations during mouse development. Mol. Cell. Neurosci. 2006, 31, 232–242. [Google Scholar] [CrossRef]
- Hruska, M.; Keefe, J.; Wert, D.; Tekinay, A.B.; Hulce, J.J.; Ibanez-Tallon, I.; Nishi, R. Prostate stem cell antigen is an endogenous lynx1-like prototoxin that antagonizes alpha7-containing nicotinic receptors and prevents programmed cell death of parasympathetic neurons. J. Neurosci. 2009, 29, 14847–14854. [Google Scholar] [CrossRef]
- Shetty, J.; Wolkowicz, M.J.; Digilio, L.C.; Klotz, K.L.; Jayes, F.L.; Diekman, A.B.; Westbrook, V.A.; Farris, E.M.; Hao, Z.; Coonrod, S.A.; Flickinger, C.J.; Herr, J.C. SAMP14, a novel, acrosomal membrane-associated, glycosylphosphatidylinositol-anchored member of the Ly-6/urokinase-type plasminogen activator receptor superfamily with a role in sperm-egg interaction. J. Biol. Chem. 2003, 278, 30506–30515. [Google Scholar] [CrossRef]
- Arredondo, J.; Chernyavsky, A.I.; Webber, R.J.; Grando, S.A. Biological effects of SLURP-1 on human keratinocytes. J. Invest. Dermatol. 2005, 125, 1236–1241. [Google Scholar] [CrossRef]
- Koh, K.; Joiner, W.J.; Wu, M.N.; Yue, Z.; Smith, C.J.; Sehgal, A. Identification of SLEEPLESS, a sleep-promoting factor. Science 2008, 321, 372–376. [Google Scholar] [CrossRef]
- Wu, M.N.; Joiner, W.J.; Dean, T.; Yue, Z.; Smith, C.J.; Chen, D.; Hoshi, T.; Sehgal, A.; Koh, K. SLEEPLESS, a Ly-6/neurotoxin family member, regulates the levels, localization and activity of Shaker. Nat. Neurosci. 2010, 13, 69–75. [Google Scholar] [CrossRef]
- Mallya, M.; Campbell, R.D.; Aguado, B. Characterization of the five novel Ly-6 superfamily members encoded in the MHC, and detection of cells expressing their potential ligands. Protein Sci. 2006, 15, 2244–2256. [Google Scholar] [CrossRef]
- Tsuji, H.; Okamoto, K.; Matsuzaka, Y.; Iizuka, H.; Tamiya, G.; Inoko, H. SLURP-2, a novel member of the human Ly-6 superfamily that is up-regulated in psoriasis vulgaris. Genomics 2003, 81, 26–33. [Google Scholar] [CrossRef]
- Hickey, M.J. Has Ly6G finally found a job? Blood 2012, 120, 1352–1353. [Google Scholar] [CrossRef]
- Huang, Y.; Qiao, F.; Abagyan, R.; Hazard, S.; Tomlinson, S. Defining the CD59-C9 binding interaction. J. Biol. Chem. 2006, 281, 27398–27404. [Google Scholar] [CrossRef]
- Monleon, I.; Martinez-Lorenzo, M.J.; Anel, A.; Lasierra, P.; Larrad, L.; Pineiro, A.; Naval, J.; Alava, M.A. CD59 cross-linking induces secretion of APO2 ligand in overactivated human T cells. Eur. J. Immunol. 2000, 30, 1078–1087. [Google Scholar] [CrossRef]
- Craik, D.J.; Daly, N.L.; Waine, C. The cystine knot motif in toxins and implications for drug design. Toxicon 2001, 39, 43–60. [Google Scholar] [CrossRef]
- Saeki, N.; Gu, J.; Yoshida, T.; Wu, X. Prostate stem cell antigen: a Jekyll and Hyde molecule? Clin. Cancer Res. 2010, 16, 3533–3538. [Google Scholar]
- Omine, M.; Kinoshita, T.; Nakakuma, H.; Maciejewski, J.P.; Parker, C.J.; Socie, G. Paroxysmal nocturnal hemoglobinuria. Int. J. Hematol. 2005, 82, 417–421. [Google Scholar] [CrossRef]
- Nevo, Y.; Ben-Zeev, B.; Tabib, A.; Straussberg, R.; Anikster, Y.; Shorer, Z.; Fattal-Valevski, A.; Ta-Shma, A.; Aharoni, S.; Rabie, M.; et al. CD59 deficiency is associated with chronic hemolysis and childhood relapsing immune-mediated polyneuropathy. Blood 2013, 121, 129–135. [Google Scholar] [CrossRef]
- Boutet, E.; Lieberherr, D.; Tognolli, M.; Schneider, M.; Bairoch, A. UniProtKB/Swiss-Prot. Methods Mol. Biol. 2007, 406, 89–112. [Google Scholar]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef]
- Soding, J.; Biegert, A.; Lupas, A.N. The HHpred interactive server for protein homology detection and structure prediction. Nucleic Acids Res. 2005, 33, W244–W248. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Tirosh, Y.; Ofer, D.; Eliyahu, T.; Linial, M. Short Toxin-like Proteins Attack the Defense Line of Innate Immunity. Toxins 2013, 5, 1314-1331. https://doi.org/10.3390/toxins5071314
Tirosh Y, Ofer D, Eliyahu T, Linial M. Short Toxin-like Proteins Attack the Defense Line of Innate Immunity. Toxins. 2013; 5(7):1314-1331. https://doi.org/10.3390/toxins5071314
Chicago/Turabian StyleTirosh, Yitshak, Dan Ofer, Tsiona Eliyahu, and Michal Linial. 2013. "Short Toxin-like Proteins Attack the Defense Line of Innate Immunity" Toxins 5, no. 7: 1314-1331. https://doi.org/10.3390/toxins5071314
APA StyleTirosh, Y., Ofer, D., Eliyahu, T., & Linial, M. (2013). Short Toxin-like Proteins Attack the Defense Line of Innate Immunity. Toxins, 5(7), 1314-1331. https://doi.org/10.3390/toxins5071314