Procoagulant Adaptation of a Blood Coagulation Prothrombinase-like Enzyme Complex in Australian Elapid Venom

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Identification of the Venom Prothrombinase-like Complex
3. Structural Characteristics of the Venom FXa-like Catalytic Subunit
3.1. Blood Coagulation Factor Xa
3.2. The Venom-Derived Factor Xa-like Catalytic Subunit

3.2.1. The Activation Peptide of Venom FX
3.2.2. The Light Chain of Venom FXa
3.2.3. The Heavy Chain of Venom FXa
4. Structural Characteristics of the Venom FVa-like Cofactor Subunit
4.1. Blood Coagulation Factor V
4.2. The Factor Va-like Cofactor Subunit
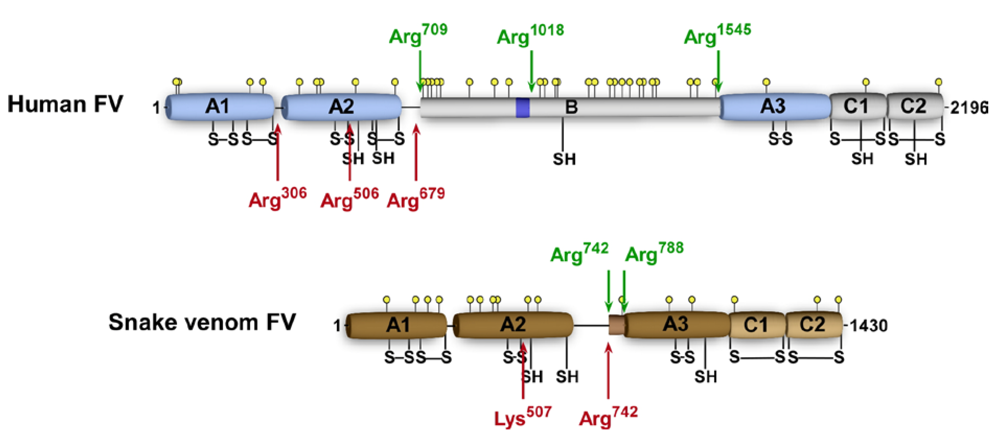
4.2.1. The B Domain of Venom FV
4.2.2. The Disulfide Bond Connecting the Heavy and Light Chains of Venom FV
4.2.3. Activated Protein C Cleavage Sites in Venom FV
4.2.4. Interdomain Connections in Venom FV
4.2.5. Posttranslational Modifications of Venom FV
5. Evolutionary Adaptation of Prothrombinase to a Powerful Hemostatic Toxin
5.1. Transcriptional Regulation and Protein Expression of the Venom Prothrombinase-like Complex
5.2. Gain-of-Function Adaptations in the P. textilis Venom-Derived FVa-like Subunit
6. Final Remarks
Acknowledgements
Conflicts of Interest
References
- Broad, A.J.; Sutherland, S.K.; Coulter, A.R. The lethality in mice of dangerous Australian and other snake venom. Toxicon 1979, 17, 661–664. [Google Scholar]
- Speijer, H.; Govers-Riemslag, J.W.; Zwaal, R.F.; Rosing, J. Prothrombin activation by an activator from the venom of Oxyuranus scutellatus (Taipan snake). J. Biol. Chem. 1986, 261, 13258–13267. [Google Scholar] [PubMed]
- Masci, P.P.; Whitaker, A.N.; de Jersey, J. Purification and characterization of a prothrombin activator from the venom of the Australian brown snake, Pseudonaja textilis textilis. Biochem. Int. 1988, 17, 825–835. [Google Scholar] [PubMed]
- Rao, V.S.; Kini, R.M. Pseutarin C, a prothrombin activator from Pseudonaja textilis venom: its structural and functional similarity to mammalian coagulation factor Xa-Va complex. Thromb. Haemost. 2002, 88, 611–619. [Google Scholar] [PubMed]
- Lavin, M.F.; Masci, P.P. Prothrombinase complexes with different physiological roles. Thromb. Haemost. 2009, 102, 421–423. [Google Scholar]
- Mann, K.G.; Nesheim, M.E.; Church, W.R.; Haley, P.E.; Krishnaswamy, S. Surface dependent reactions of the vitamin K-dependent enzyme complexes. Blood 1990, 76, 1–16. [Google Scholar]
- Chester, A.; Crawford, G.P. In vitro coagulant properties of venoms from Australian snakes. Toxicon 1982, 20, 501–504. [Google Scholar] [CrossRef] [PubMed]
- Rosing, J.; Tans, G. Structural and functional properties of snake venom prothrombin activators. Toxicon 1992, 30, 1515–1527. [Google Scholar]
- Kini, R.M. The intriguing world of prothrombin activators from snake venom. Toxicon 2005, 45, 1133–1145. [Google Scholar]
- Denson, K.W. Coagulant and anticoagulant action of snake venoms. Toxicon 1969, 7, 5–11. [Google Scholar]
- Owen, W.G.; Jackson, C.M. Activation of prothrombin with Oxyuranus scutellatus scutellatus (taipan snake) venom. Thromb. Res. 1973, 3, 705–714. [Google Scholar] [CrossRef]
- Walker, F.J.; Owen, W.G.; Esmon, C.T. Characterization of the prothrombin activator from the venom of Oxyuranus scutellatus scutellatus (taipan venom). Biochemistry 1980, 19, 1020–1023. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Rezaie, A.R. Proexosite-1-dependent recognition and activation of prothrombin by taipan venom. J. Biol. Chem. 2004, 279, 17869–17874. [Google Scholar]
- Rao, V.S.; Swarup, S.; Kini, R.M. The nonenzymatic subunit of pseutarin C, a prothrombin activator from eastern brown snake (Pseudonaja textilis) venom, shows structural similarity to mammalian coagulation factor V. Blood 2003, 102, 1347–1354. [Google Scholar] [CrossRef] [PubMed]
- Rao, V.S.; Swarup, S.; Kini, R.M. The catalytic subunit of pseutarin C, a group C prothrombin activator from the venom of Pseudonaja textilis, is structrually similar to mammalian blood coagulation factor Xa. Thromb. Haemost. 2004, 92, 509–521. [Google Scholar] [PubMed]
- Welton, R.E.; Burnell, J.N. Full length nucleotide sequence of a factor V-like subunit of oscutarin from Oxyuranus scutellatus scutellatus (coastal Taipan). Toxicon 2005, 46, 328–336. [Google Scholar]
- St. Pierre, L.; Masci, P.P.; Filippovich, I.; Sorokina, N.; Marsh, N.; Miller, D.J.; Lavin, M.F. Comparative analysis of prothrombin activators from the venom of Australian elapids. Mol. Biol. Evol. 2005, 22, 1853–1864. [Google Scholar]
- Filippovich, I.; Sorokina, N.; St Pierre, L.; Flight, S.; de Jersey, J.; Perry, N.; Masci, P.P.; Lavin, M.F. Cloning and functional expression of venom prothrombin activator protease from Pseudonaja textilis with whole blood procoagulant activity. Br. J. Haematol. 2005, 131, 237–246. [Google Scholar]
- Minh Le, T.N.; Reza, M.A.; Swarup, S.; Kini, R.M. Gene duplication of coagulation factor V and origin of venom prothrombin activator in Pseudonaja textilis snake. Thromb. Haemost. 2005, 93, 420–429. [Google Scholar] [PubMed]
- Furie, B.; Bouchard, B.A.; Furie, B.C. Vitamin K-dependent biosynthesis of γ-carboxyglutamic acid. Blood 1999, 93, 1798–1808. [Google Scholar]
- Sunnerhagen, M.; Forsén, S.; Hoffrén, A.-M.; Drakenberg, T.; Teleman, O.; Stenflo, J. Structure of the Ca2+-free Gla domain sheds light on membrane binding of blood coagulation proteins. Nat. Struct. Biol. 1995, 2, 504–509. [Google Scholar]
- Bode, W.; Mayr, I.; Bauman, Y.; Huber, R.; Stone, S.R.; Hofsteenge, J. The refined 1.9 Å crystal structure of human α-thrombin: Interaction with D-Phe-Pro-Arg chloromethylketone and significance of the Tyr-Pro-Trp insertion segment. EMBO J. 1989, 8, 3467–3475. [Google Scholar] [PubMed]
- Huber, R.; Bode, W. Structural basis of the activation and action of trypsin. Acc. Chem. Res. 1978, 11, 114–122. [Google Scholar]
- Reza, M.A.; Minh Le, T.N.; Swarup, S.; Manjunatha, K.R. Molecular evolution caught in action: gene duplication and evolution of molecular isoforms of prothrombin activators in Pseudonaja textilis (brown snake). J. Thromb. Haemost. 2006, 4, 1346–1353. [Google Scholar]
- Rudolph, A.E.; Mullane, M.P.; Porche-Sorbet, R.; Daust, H.A.; Miletich, J.P. The role of the factor X activation peptide: a deletion mutagenesis approach. Thromb. Haemost. 2002, 88, 756–762. [Google Scholar]
- Yang, L.; Manithody, C.; Rezaie, A.R. Functional role of O-linked and N-linked glycosylation sites present on the activation peptide of factor X. J. Thromb. Haemost. 2009, 7, 1696–1702. [Google Scholar]
- Bos, M.H.A.; Boltz, M.; St. Pierre, L.; Masci, P.P.; de Jersey, J.; Lavin, M.F.; Camire, R.M. Venom factor V from the common brown snake escapes hemostatic regulation through procoagulant adaptations. Blood 2009, 114, 686–692. [Google Scholar]
- Toso, R.; Zhu, H.; Camire, R.M. The conformational switch from the factor X zymogen to protease state mediates exosite expression and prothrombinase assembly. J. Biol. Chem. 2008, 283, 18627–18635. [Google Scholar]
- Mann, K.G.; Kalafatis, M. Factor V: A combination of Dr. Jekyll and Mr. Hyde. Blood 2002, 101, 20–30. [Google Scholar] [PubMed]
- Camire, R.M.; Bos, M.H.A. The molecular basis of factor V and VIII procofactor activation. J. Thromb. Haemost. 2009, 7, 1951–1961. [Google Scholar]
- Toso, R.; Camire, R.M. Removal of B-domain sequences from factor V rather than specific proteolysis underlies the mechanism by which cofactor function is realized. J. Biol. Chem. 2004, 279, 21643–21650. [Google Scholar]
- Zhu, H.; Toso, R.; Camire, R.M. Inhibitory sequences within the B-domain stabilize circulating factor V in an inactive state. J. Biol. Chem. 2007, 282, 15033–15039. [Google Scholar]
- Adams, T.E.; Hockin, M.F.; Mann, K.G.; Everse, S.J. The crystal structure of activated protein C-inactivated bovine factor Va: Implications for cofactor function. Proc. Natl. Acad. Sci. USA 2004, 101, 8918–8923. [Google Scholar]
- Birrell, G.W.; Earl, S.; Masci, P.P.; de Jersey, J.; Wallis, T.P.; Gorman, J.J.; Lavin, M.F. Molecular diversity in venom from the Australian Brown snake, Pseudonaja textilis. Mol. Cell. Proteomics 2006, 5, 379–389. [Google Scholar]
- Kalafatis, M. Identification and partial characterization of factor Va heavy-chain kinase from human platelets. J. Biol. Chem. 1998, 273, 8459–8466. [Google Scholar]
- Pittman, D.D.; Tomkinson, K.N.; Michnick, D.; Seligsohn, U.; Kaufman, R.J. Posttranslational sulfation of factor V is required for efficient thrombin cleavage and activation and for full procoagulant activity. Biochemistry 1994, 33, 6952–6959. [Google Scholar]
- Masci, P.P.; Rowe, E.A.; Whitaker, A.N.; de Jersey, J. Fibrinolysis as a feature of disseminated intravascular coagulation (DIC) after Pseudonaja textilis textilis envenomation. Thromb. Res. 1990, 59, 859–870. [Google Scholar] [CrossRef] [PubMed]
- Lalloo, D.G.; Trevett, A.J.; Korinhona, A.; Nwokolo, N.; Laurenson, I.F.; Paul, M.; Black, J.; Naraqi, S.; Mavo, B.; Saweri, A.; et al. Snake bites by the Papuan taipan (Oxyuranus scutellatus canni): paralysis, hemostatic and electrocardiographic abnormalities, and effects of antivenom. Am. J. Trop. Med. Hyg. 1995, 52, 525–531. [Google Scholar] [PubMed]
- Barrett, R.; Little, M. Five years of snake envenoming in far north Queensland. Emerg. Med. (Fremantle) 2003, 15, 500–510. [Google Scholar] [CrossRef] [PubMed]
- Currie, B.J. Snakebite in tropical Australia: a prospective study in the "Top End" of the Northern Territory. Med. J. Aust. 2004, 181, 693–697. [Google Scholar]
- Reza, M.A.; Swarup, S.; Kini, R.M. Structure of two genes encoding parallel prothrombin activators in Tropidechis carinatus snake: gene duplication and recruitment of factor X gene to the venom gland. J. Thromb. Haemost. 2007, 5, 117–126. [Google Scholar]
- Kwong, S.; Woods, A.E.; Mirtschin, P.J.; Ge, R.; Kini, R.M. The recruitment of blood coagulation factor X into snake venom gland as a toxin: the role of promoter cis-elements in its expression. Thromb. Haemost. 2009, 102, 469–478. [Google Scholar]
- Mirtschin, P.J.; Dunstan, N.; Hough, B.; Hamilton, E.; Klein, S.; Lucas, J.; Millar, D.; Madaras, F.; Nias, T. Venom yields from Australian and some other species of snakes. Ecotoxicology 2006, 15, 531–538. [Google Scholar]
- Mann, K.G.; Brummel, K.; Butenas, S. What is all that throm bin for? J. Thromb. Haemost. 2003, 1, 1504–1514. [Google Scholar] [CrossRef] [PubMed]
- Krishnaswamy, S.; Jones, K.C.; Mann, K.G. Prothrombinase complex assembly: Kinetic mechanism of enzyme assembly on phospholipid vesicles. J. Biol. Chem. 1988, 263, 3823–3834. [Google Scholar]
- Majumder, R.; Quinn-Allen, M.A.; Kane, W.H.; Lentz, B.R. A phosphatidylserine binding site in factor Va C1 domain regulates both assembly and activity of the prothrombinase complex. Blood 2008, 112, 2795–2802. [Google Scholar]
- Qureshi, S.H.; Yang, L.; Manithody, C.; Rezaie, A.R. Membrane-dependent interaction of factor Xa and prothrombin with factor Va in the prothrombinase complex. Biochemistry 2009, 48, 5034–5041. [Google Scholar]
- Fohlman, J.; Lind, P.; Eaker, D. Taipoxin, an extremely potent presynaptic snake venom neurotoxin. Elucidation of the primary structure of the acidic carbohydrate-containing taipoxin-subunit, a prophospholipase homolog. FEBS Lett. 1977, 84, 367–371. [Google Scholar]
- Pearson, J.A.; Tyler, M.I.; Retson, K.V.; Howden, M.E. Studies on the subunit structure of textilotoxin, a potent presynaptic neurotoxin from the venom of the Australian common brown snake (Pseudonaja textilis). 3. The complete amino-acid sequences of all the subunits. Biochim. Biophys. Acta 1993, 1161, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Joseph, J.S.; Chung, M.C.; Mirtschin, P.J.; Kini, R.M. Effect of snake venom procoagulants on snake plasma: implications for the coagulation cascade of snakes. Toxicon 2002, 40, 175–183. [Google Scholar]
- Thwin, M.M.; Gopalakrishnakone, P. Snake envenomation and protective natural endogenous proteins: a mini review of the recent developments (1991–1997). Toxicon 1998, 36, 1471–1482. [Google Scholar]
- Takacs, Z.; Wilhelmsen, K.C.; Sorota, S. Cobra (Naja spp.) nicotinic acetylcholine receptor exhibits resistance to Erabu sea snake (Laticauda semifasciata) short-chain alpha-neurotoxin. J. Mol. Evol. 2004, 58, 516–526. [Google Scholar] [CrossRef] [PubMed]
© 2010 by the authors; licensee MDPI, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Bos, M.H.A.; Camire, R.M. Procoagulant Adaptation of a Blood Coagulation Prothrombinase-like Enzyme Complex in Australian Elapid Venom. Toxins 2010, 2, 1554-1567. https://doi.org/10.3390/toxins2061554
Bos MHA, Camire RM. Procoagulant Adaptation of a Blood Coagulation Prothrombinase-like Enzyme Complex in Australian Elapid Venom. Toxins. 2010; 2(6):1554-1567. https://doi.org/10.3390/toxins2061554
Chicago/Turabian StyleBos, Mettine H.A., and Rodney M. Camire. 2010. "Procoagulant Adaptation of a Blood Coagulation Prothrombinase-like Enzyme Complex in Australian Elapid Venom" Toxins 2, no. 6: 1554-1567. https://doi.org/10.3390/toxins2061554
APA StyleBos, M. H. A., & Camire, R. M. (2010). Procoagulant Adaptation of a Blood Coagulation Prothrombinase-like Enzyme Complex in Australian Elapid Venom. Toxins, 2(6), 1554-1567. https://doi.org/10.3390/toxins2061554