Transcriptome Analysis of Dimethyl Fumarate Inhibiting the Growth of Aspergillus carbonarius

 ,
, 
Abstract
1. Introduction
2. Results
2.1. Inhibitory Effect of Dimethyl Fumarate on Mycelial Growth
2.2. SEM Analysis of Mycelial Morphology
2.3. Transcriptomic Analysis
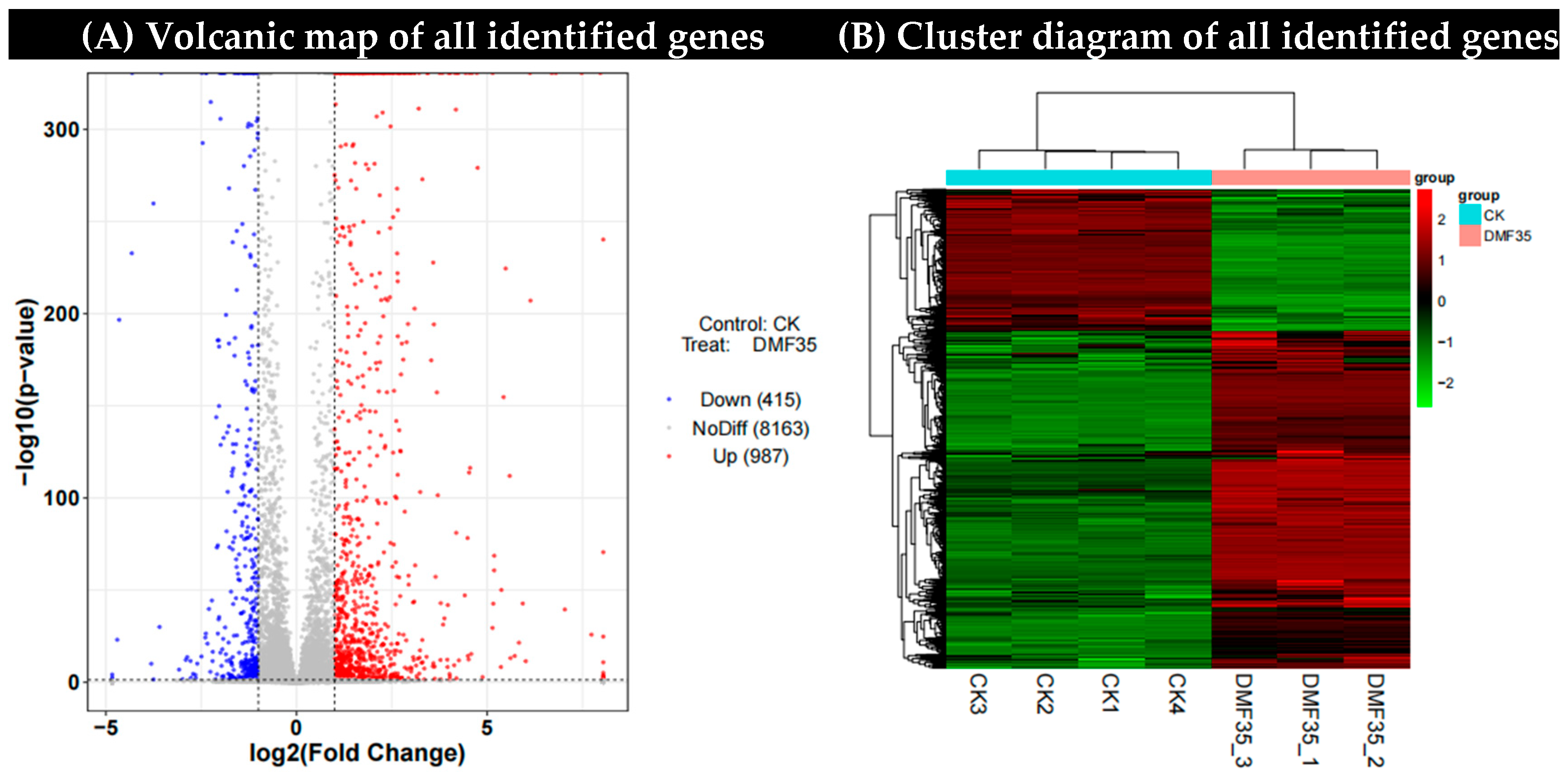
2.3.1. Identification and Screening of DEGs Under Dimethyl Fumarate Treatment
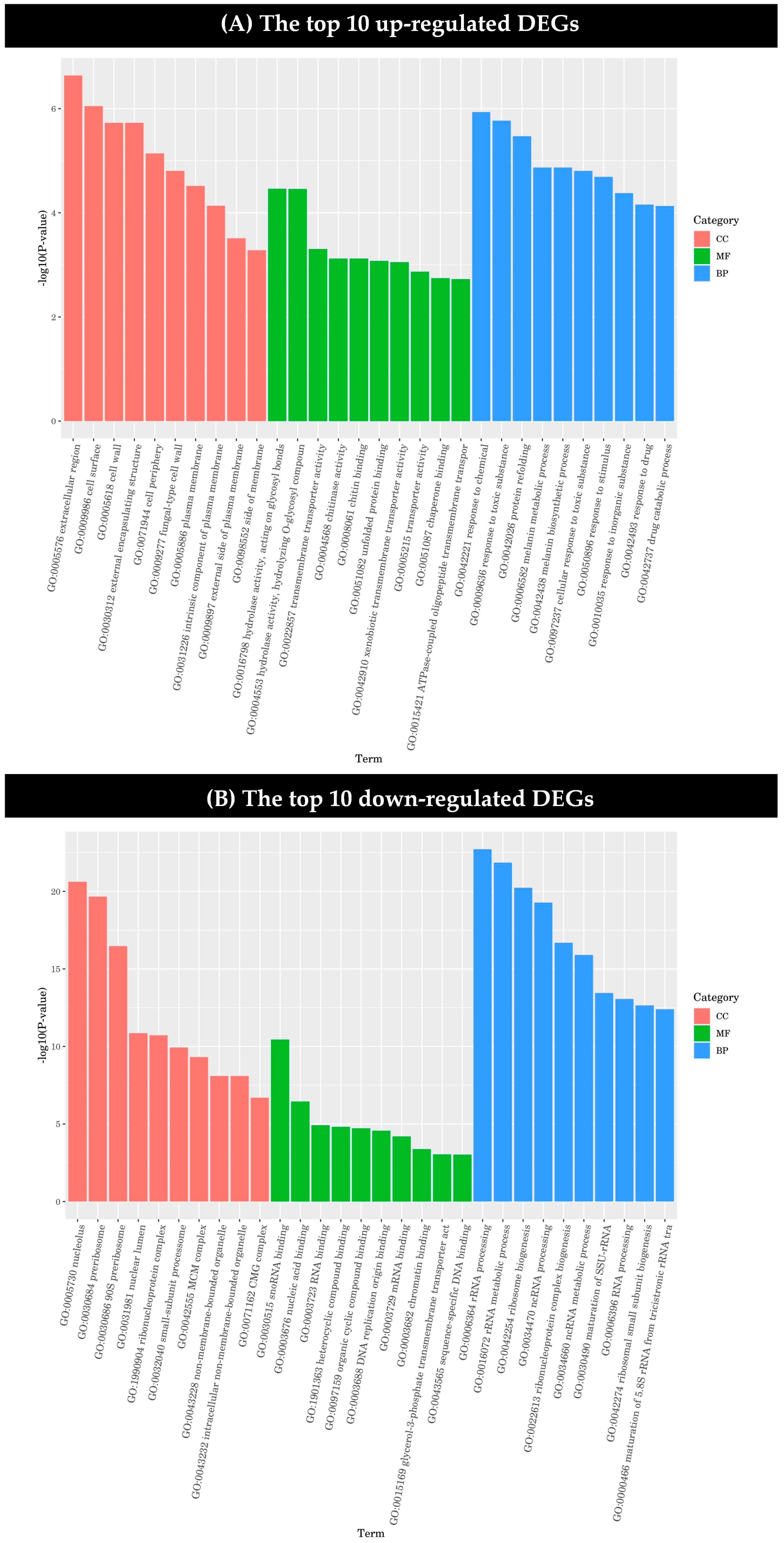
2.3.2. Functional Classification of DEGs and Pathway Analysis
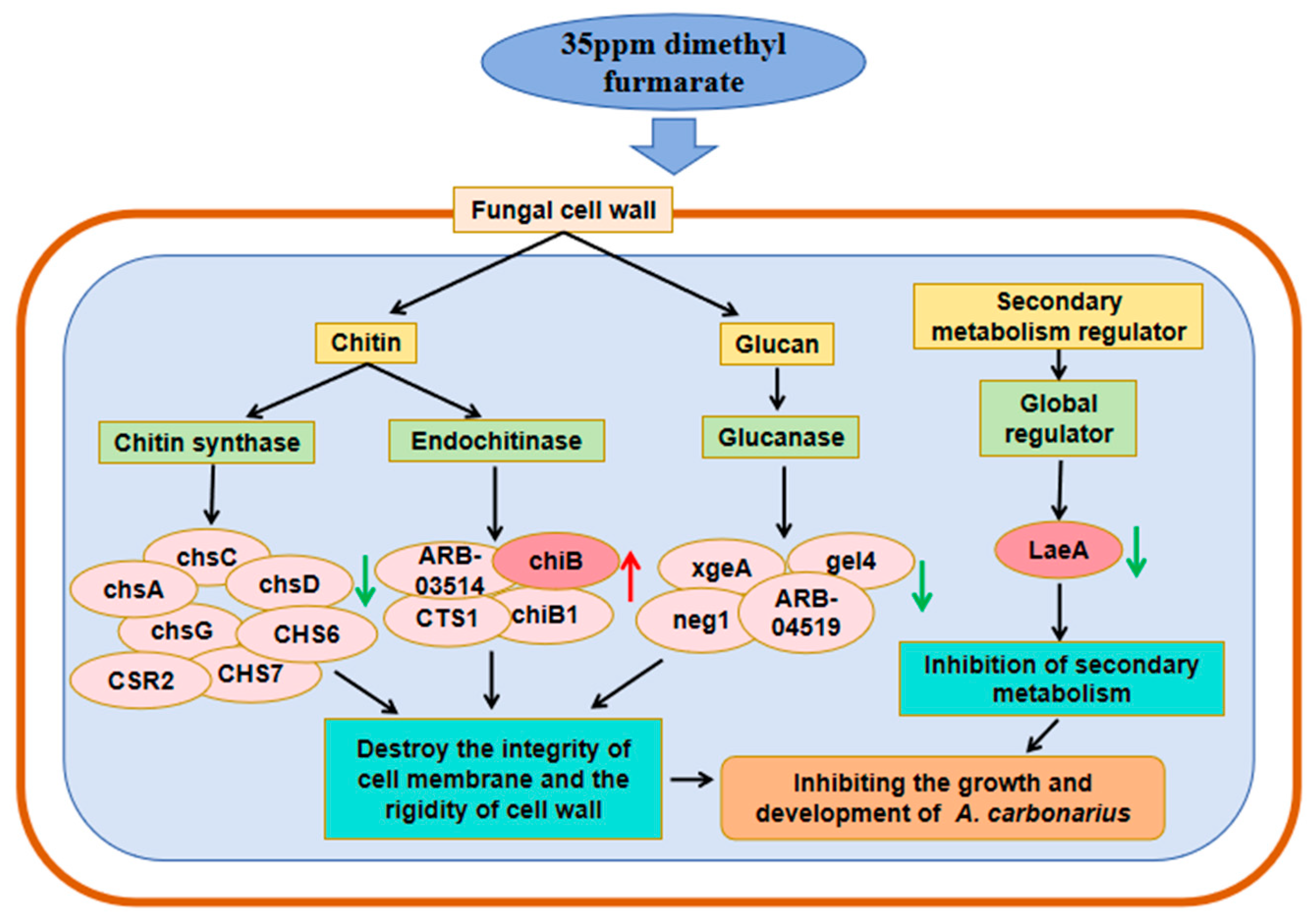
2.3.3. Functional Analysis of DEGs
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Chemicals, Fungal Strain, and Growth Conditions
5.2. Antifungal Activity of Dimethyl Fumarate
5.3. OTA Detection
5.4. Scanning Electron Microscopy (SEM) Analysis
5.5. Total RNA Isolation, cDNA Library Construction, and Sequencing
5.6. Database Search and Analysis
5.7. Differential Expression Analysis
5.8. Functional Enrichment Analysis of Differentially Expressed Genes
5.9. Gene Expression Analysis Performed with RT-qPCR
5.10. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| DEGs | Differentially expressed genes |
| OTA | Ochratoxin A |
| HPLC | High-performance liquid chromatography |
References
- Huo, X.; Tian, X.; Liu, Z.; Wang, L.; Kong, Q.; Wang, D.; Ren, X. Combination of LED blue light with peppermint essential oil emulsion for the postharvest storage of Shine Muscat grape to control Aspergillus carbonarius. Postharvest Biol. Technol. 2024, 218, 113175. [Google Scholar] [CrossRef]
- Hu, G.; Chen, M.; Wang, Q.; Guo, D. Effect of prochloraz on production activity of ochratoxin on grape surface. Agrochemical 2023, 62, 923–928. [Google Scholar]
- Shi, M.; Zhang, Z.; Gao, C.; Li, F.; Dong, R.; Qiao, D.; Xue, F.; Wei, D.; Wu, Y.; Huang, W.; et al. Effects of melatonin treatment on resistance to Aspergillus carbonarius in postharvest ‘Cabernet Sauvignon’ grape. Plant Physiol. J. 2024, 60, 461–470. [Google Scholar]
- Peng, Q.; Yang, B.; Wang, L.; Yang, S.; Zhu, X. Effects of Three Fungicidal Treatments on Growth of Aspergillus carbonarius and Production of Ochratoxin A. Sci. Technol. Food Ind. 2019, 40, 152–157. [Google Scholar]
- Zhao, Z.; Cai, W.; Song, L.; Mu, X.; Hu, Y. Comprehensive Property Investigation of Mold Inhibitor Treated Raw Cotton and Ramie Fabric. Materials 2020, 13, 1105. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Liu, H.; Zhang, J.; Ci, H. Study on the Determination of Dimethyl Fumarate in Food. Modern. Food 2018, 2, 85–87. [Google Scholar]
- Oriella, G.; Domenico, M.; Antonio, S.; Stefania, A.; Lucia, G. Determination of dimethyl fumarate in desiccant and antimould sachets by reversed-phase liquid chromatography. Biomed. Chromatogr. BMC 2011, 25, 1315–1318. [Google Scholar]
- Lategan, W.T.; Wang, L.; Sprague, N.T.; Rousseau, F.S. Pharmacokinetics and Bioavailability of Monomethyl Fumarate Following a Single Oral Dose of Bafiertam™ (Monomethyl Fumarate) or Tecfidera ® (Dimethyl Fumarate). CNS Drugs 2021, 35, 567–574. [Google Scholar] [CrossRef]
- Salvatore, G.; Grazia, A.D.; Grazia, M.; Claudio, B.; Luca, R.G.; Settimio, R.; Chiara, M.E.; Elisabetta, P.; Velia, D. Drug-Repurposing Strategy for Dimethyl Fumarate. Pharmaceuticals 2023, 16, 974. [Google Scholar] [CrossRef]
- Ning, M.; Yi, W.; Fei, X.; Du, K.; Wang, Y.; Shi, L.; Ji, L.; Liu, T.; Ma, X. Dimethyl fumarate reduces the risk of mycotoxins via improving intestinal barrier and microbiota. Oncotarget 2017, 8, 44625–44638. [Google Scholar]
- Wang, L.; Jin, J.; Liu, X.; Wang, Y.; Liu, Y.; Zhao, Y.; Xing, F. Effect of cinnamaldehyde on morphological alterations of Aspergillus ochraceus and expression of key genes involved in ochratoxin A biosynthesis. Toxins 2018, 10, 340. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Jin, J.; Zheng, M.; Yang, Q.; Xing, F. Ethanol inhibits aflatoxin B1 biosynthesis in Aspergillus flavus by up-regulating oxidative stress-related genes. Front. Microbiol. 2020, 10, 2946. [Google Scholar] [CrossRef]
- Xing, F.; Wang, L.; Liu, X.; Selvaraj, J.N.; Wang, Y.; Zhao, Y.; Liu, Y. Aflatoxin B1 inhibition in Aspergillus flavus by Aspergillus niger through down-regulating expression of major biosynthetic genes and AFB1 degradation by atoxigenic A. flavus. Int. J. Food Microbiol. 2017, 256, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.; Wang, L.; Jiang, D.; Wang, M.; Liu, H.; Yu, H.; Yao, W. Transcriptomic analysis of inhibition by eugenol of ochratoxin A biosynthesis and growth of Aspergillus carbonarius. Food Control 2022, 135, 108788. [Google Scholar] [CrossRef]
- Zhang, J. The Synthesis and Application of Dimethyl Fumarate as Fungicide on Stored Tobacco Mildew; Jiangnan University: Wuxi, China, 2008. [Google Scholar]
- Bi, L.; Zheng, W.; Li, W. Synthesis of Dimethyl Fumarate and Its Application as a New Type Mildew Inhibitor Preservative. Yunnan Chem. Technol. 1999, 1, 58–60. [Google Scholar]
- Bok, J.W.; Keller, N.P. LaeA, a Regulator of Secondary Metabolism in Aspergillus spp. Eukaryot Cell 2004, 3, 527–535. [Google Scholar] [CrossRef]
- Maor, U.; Barda, O.; Sadhasivam, S.; Bi, Y.; Levin, E.; Zakin, V.; Prusky, D.; Sionov, E. Functional roles of LaeA, polyketide synthase, and glucose oxidase in the regulation of ochratoxin A biosynthesis and virulence in Aspergillus carbonarius. Mol. Plant Pathol. 2020, 22, 117–129. [Google Scholar] [CrossRef]
- Yuta, T.; Jun, T.; Shihori, A.; Takahiro, M.; Takehito, N.; Kenji, W. Genomic Mushroom Hunting Decrypts Coprinoferrin, A Siderophore Secondary Metabolite Vital to Fungal Cell Development. Org. Lett. 2019, 21, 7582–7586. [Google Scholar]
- Wang, Y.; Chen, H.; Guo, Z.; Sun, L.; Fu, Y.; Li, T.; Lin, W.; Lin, X. Quantitative proteomic analysis of iron-regulated outer membrane proteins in Aeromonas hydrophila as potential vaccine candidates. Fish Shellfish Immunol. 2017, 68, 1–9. [Google Scholar] [CrossRef]
- Matthew, J.; Yang, H.; Hong, N.; Munro, P.; Will, S.; Junyong, K.; Neil, G.; Wenjun, Z.; Rachel, O.; Yi, T. Elucidating the molecular programming of a nonlinear non-ribosomal peptide synthetase responsible for fungal siderophore biosynthesis. Nat. Commun. 2023, 14, 2832. [Google Scholar]
- Kaur, R.; Kalia, A.; Lore, J.S.; Kaur, A.; Yadav, I.; Sharma, P.; Sandhu, J.S. Trichoderma sp. endochitinase and β-1,3-glucanase impede Rhizoctonia solani growth independently, and their combined use does not enhance impediment. Plant Pathol. 2021, 70, 1388–1396. [Google Scholar] [CrossRef]
- Brandão, M.R.; Ferreira, F.R.V.; Batista, R.L.; Alves, E.; Lira, N.A.; Bellete, B.S.; Scolforo, J.R.S.; Cardoso, M.G. Antifungal and antimycotoxigenic effect of the essential oil of Eremanthus erythropappus on three different Aspergillus species. Flavour Fragr. J. 2020, 35, 524–533. [Google Scholar] [CrossRef]
- Yan, W.; Yong, Z.; Aiyun, Z.; Jing, N.; Di, W.; Xiao, L.S.; Lou, X.; Kai, Y. Transcriptomic analysis reveals the inhibition mechanism of pulsed light on fungal growth and ochratoxin A biosynthesis in Aspergillus carbonarius. Food Res. Int. 2023, 165, 112501. [Google Scholar]
- Gómez, V.J.; Tarazona, A.; Mateo, F.; Jiménez, M.; Mateo, E.M. Potential impact of engineered silver nanoparticles in the control of aflatoxins, OTA and the main aflatoxigenic and ochratoxigenic species affecting foods. Food Control 2019, 101, 58–68. [Google Scholar] [CrossRef]
- Barrios-González, J.; Martínez, C.; Aguilera, A.; Raimbault, M. Germination of concentrated suspensions of spores from Aspergillus niger. Biotechnol. Lett. 1989, 11, 551–554. [Google Scholar] [CrossRef]
- Yu, X.; Kong, Q.; Ren, X. Study of three kinds of essential oil on antimicrobial activity of fruit pathogenic bacteria. J. Food Sci. Biotechnol. 2017, 36, 714–719. [Google Scholar]
- Moussaoui, E.A.; Jawhari, Z.F.; Almehdi, M.A.; Elmsellem, H.; Benbrahim, K.F.; Bousta, D.; Bari, A. Antibacterial, antifungal and antioxidant activity of total polyphenols of Withania frutescens L. Bioorganic Chem. 2019, 93, 103337. [Google Scholar] [CrossRef]
- Jiang, W.; Liang, X.; Li, H.; Mo, L.; Chen, W.; Wang, T.; Wang, H.; Xing, Y.; Liao, J. Inhibitory effect of tannic acid on the growth of Apiospora arundinis and 3-Nitropropionic acid production. J. Appl. Microbiol. 2023, 134, 264. [Google Scholar] [CrossRef]
- Huang, J.; Guan, H.; Huang, Y.; Lai, K.; Chen, H.; Xue, H.; Zhang, B. Evaluating the effects of microparticle addition on mycelial morphology, natural yellow pigments productivity and key genes regulation in submerged fermentation of Monascus purpureus. Biotechnol. Bioeng. 2021, 118, 2503–2513. [Google Scholar] [CrossRef]
- Shi, X.; Qiu, H.; Wang, J.; Zhang, Z.; Wang, Y.; Sun, G. A handy method to remove bacterial contamination from fungal cultures. PLoS ONE 2019, 14, e0224635. [Google Scholar] [CrossRef]
- Kenneth, J.; Thomas, D.S. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Swissprot Name | log2FoldChange | p-Value | Annotated Gene Function | |
|---|---|---|---|---|---|
| Cell integrity | ASPCADRAFT_203630 | chiB | 2.137 | 9.288 × 10−185 | Endochitinase |
| ASPCADRAFT_125551 | chiB1 | 1.657 | 1.139 × 10−13 | ||
| ASPCADRAFT_208876 | ARB_03514 | 1.561 | 1.837 × 10−17 | ||
| ASPCADRAFT_205337 | CTS1 | 0.955 | 4.009 × 10−22 | ||
| ASPCADRAFT_210467 | xgeA | −1.438 | 1.383 × 10−106 | Glucanase | |
| ASPCADRAFT_208658 | gel4 | −0.717 | 1.000× 10−3 | ||
| ASPCADRAFT_212042 | neg1 | −0.456 | 3.283 × 10−6 | ||
| ASPCADRAFT_206132 | ARB_04519 | −0.517 | 2.478 × 10−55 | ||
| ASPCADRAFT_206390 | fcyB | 1.489 | 1.020 × 10−53 | Purine-cytosine permease | |
| ASPCADRAFT_135110 | PCH_Pc12g07500 | 1.468 | 4.070 × 10−26 | Mutanase | |
| ASPCADRAFT_503657 | mok13 | 1.053 | 2.498 × 10−49 | Cell wall alpha-1,3-glucan synthase | |
| ASPCADRAFT_203143 | nvfG | −1.923 | 3.034 × 10−24 | Ketoreductase | |
| ASPCADRAFT_211936 | chsD | −0.867 | 1.417 × 10−168 | Chitin synthase | |
| ASPCADRAFT_208758 | chsG | −0.419 | 8.630 × 10−57 | ||
| ASPCADRAFT_209480 | CSR2 | −0.519 | 5.100 × 10−32 | ||
| ASPCADRAFT_203230 | CHS6 | −0.255 | 2.991 × 10−22 | ||
| ASPCADRAFT_208304 | chsC | −0.422 | 2.749 × 10−14 | ||
| ASPCADRAFT_211141 | chsA | −0.384 | 7.000× 10−3 | ||
| ASPCADRAFT_159307 | chs7 | −0.273 | 0.844 | ||
| Development | ASPCADRAFT_208824 | acyN | 2.697 | 0.000 | Nonribosomal peptide synthetase |
| ASPCADRAFT_5941 | LaeA | −1.119 | 2.691 × 10−10 | Secondary metabolism regulator | |
| ASPCADRAFT_212444 | brlA | 2.279 | 3.074 × 10−199 | regulator of conidiophore development | |
| ASPCADRAFT_169479 | wetA | 1.091 | 2.241 × 10−6 | Developmental regulatory protein | |
| Stress response | ASPCADRAFT_207672 | LUC7 | 2.683 | 0 | Glutathione S-transferase-like protein |
| ASPCADRAFT_174114 | gst3 | 1.837 | 4.817 × 10−38 | Glutathione S-transferase 3 | |
| ASPCADRAFT_131005 | gst3 | 1.345 | 6.182 × 10−5 | Glutathione S-transferase 3 |
| Gene ID | Swissprot Name | Upstream Primer | Downstream Primer |
|---|---|---|---|
| ASPCADRAFT_5941 | laeA | taatgctcgcatcctcgacc | tgagatcccaggaatcctctc |
| ASPCADRAFT_203630 | chiB | ctgatccaacatcgacacgc | caccatgatctcctacgacactg |
| ASPCADRAFT_208824 | acyN | cattctcttccgccatcatcg | cccattggtttctgtcggtagg |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Tai, B.; Yu, X.; Li, E.; Wang, G.; Jin, J.; Xing, F. Transcriptome Analysis of Dimethyl Fumarate Inhibiting the Growth of Aspergillus carbonarius. Toxins 2025, 17, 339. https://doi.org/10.3390/toxins17070339
Wang S, Tai B, Yu X, Li E, Wang G, Jin J, Xing F. Transcriptome Analysis of Dimethyl Fumarate Inhibiting the Growth of Aspergillus carbonarius. Toxins. 2025; 17(7):339. https://doi.org/10.3390/toxins17070339
Chicago/Turabian StyleWang, Siruo, Bowen Tai, Xifan Yu, Erfeng Li, Gang Wang, Jing Jin, and Fuguo Xing. 2025. "Transcriptome Analysis of Dimethyl Fumarate Inhibiting the Growth of Aspergillus carbonarius" Toxins 17, no. 7: 339. https://doi.org/10.3390/toxins17070339
APA StyleWang, S., Tai, B., Yu, X., Li, E., Wang, G., Jin, J., & Xing, F. (2025). Transcriptome Analysis of Dimethyl Fumarate Inhibiting the Growth of Aspergillus carbonarius. Toxins, 17(7), 339. https://doi.org/10.3390/toxins17070339