Bacillus thuringiensis Spores and Cry3A Toxins Act Synergistically to Expedite Colorado Potato Beetle Mortality

 ,
,  ,
, 
 and
and
Abstract
:1. Introduction
2. Results
2.1. Susceptibility of CPB Larvae to Bt Treatments
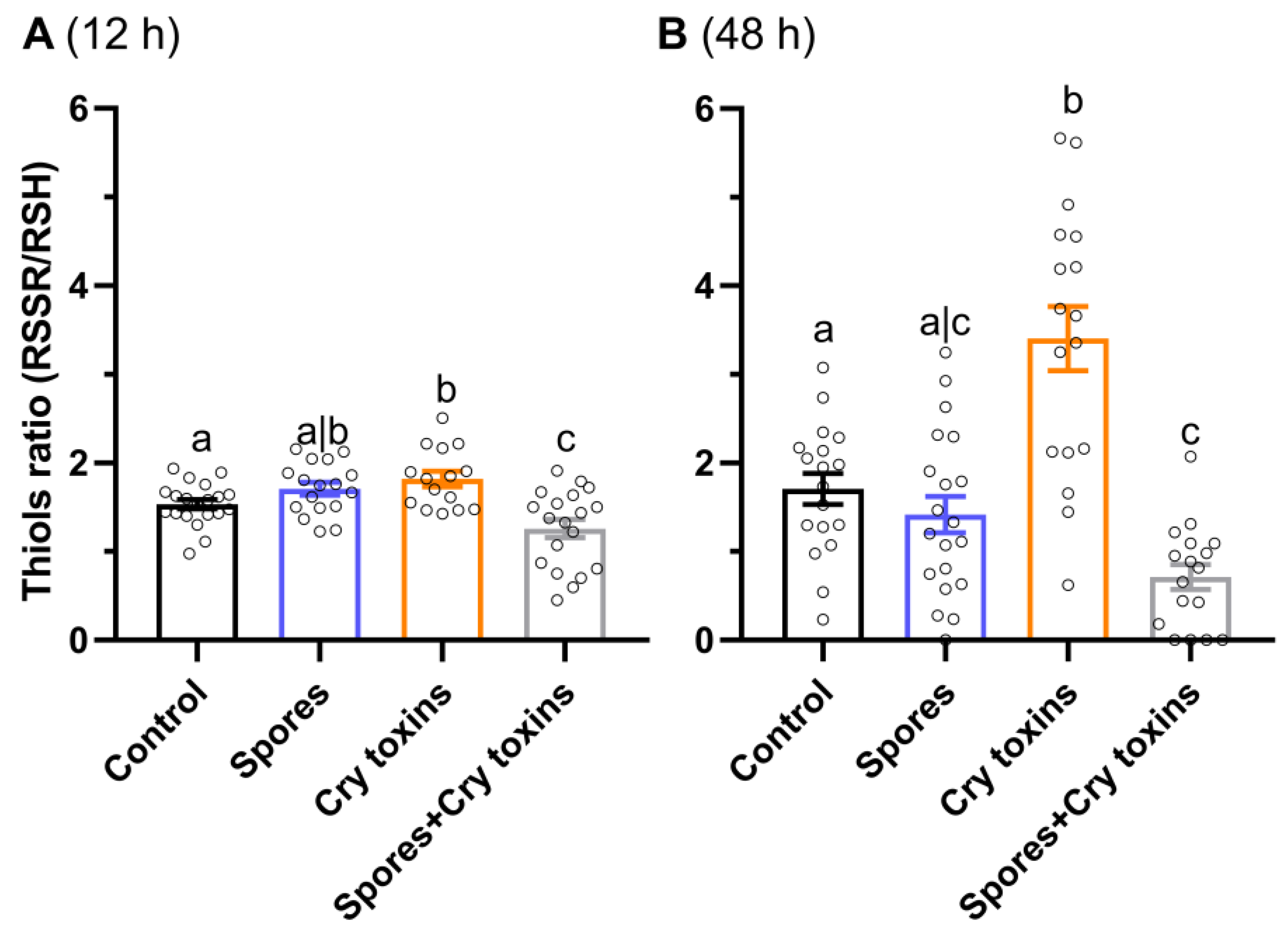
2.2. Redox Balance and Detoxification in CPB Larvae Post Bt Treatment
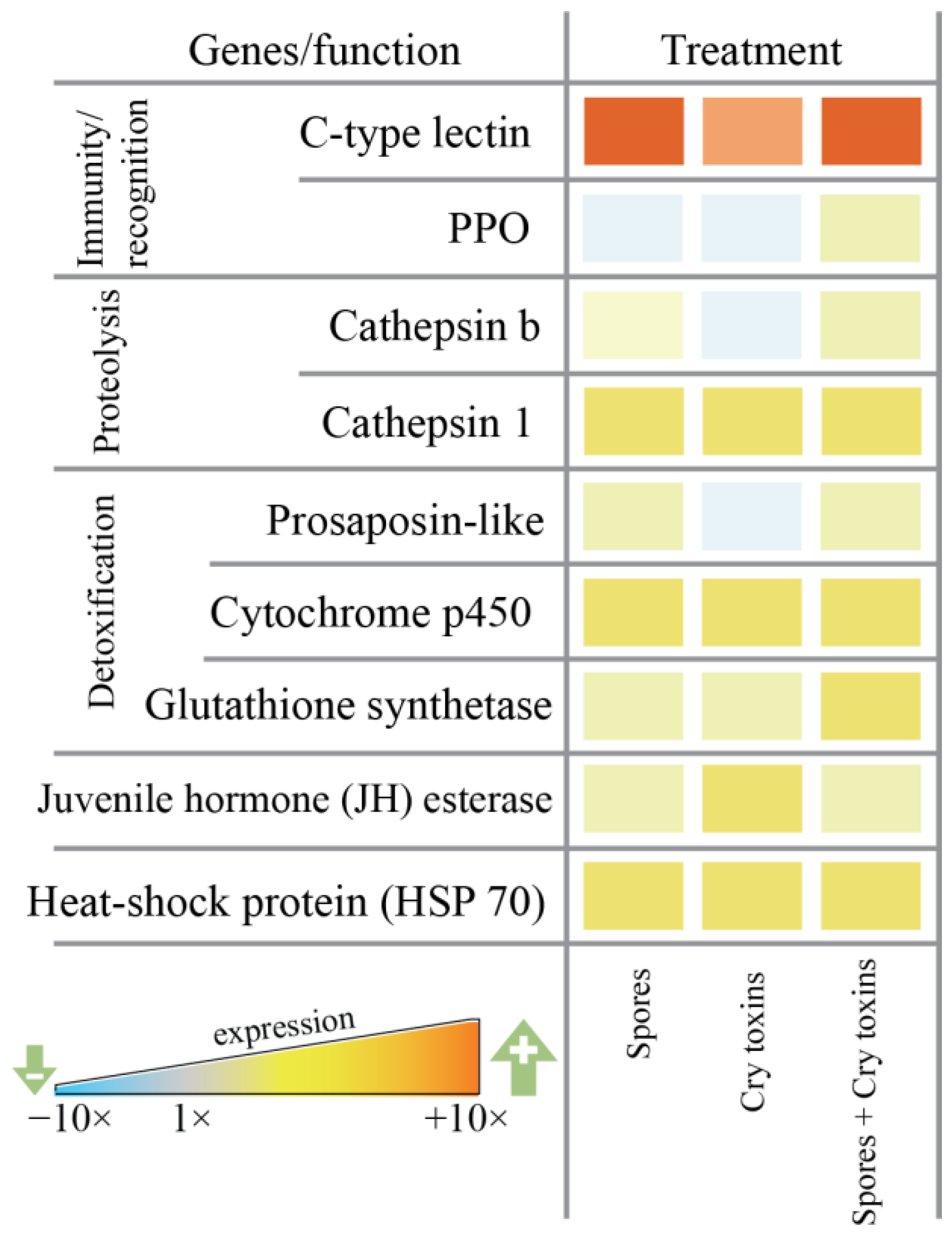
2.3. Expression of Immunity/Detoxification-Related Genes in the Midgut of CPB Larvae Post Bt Treatment
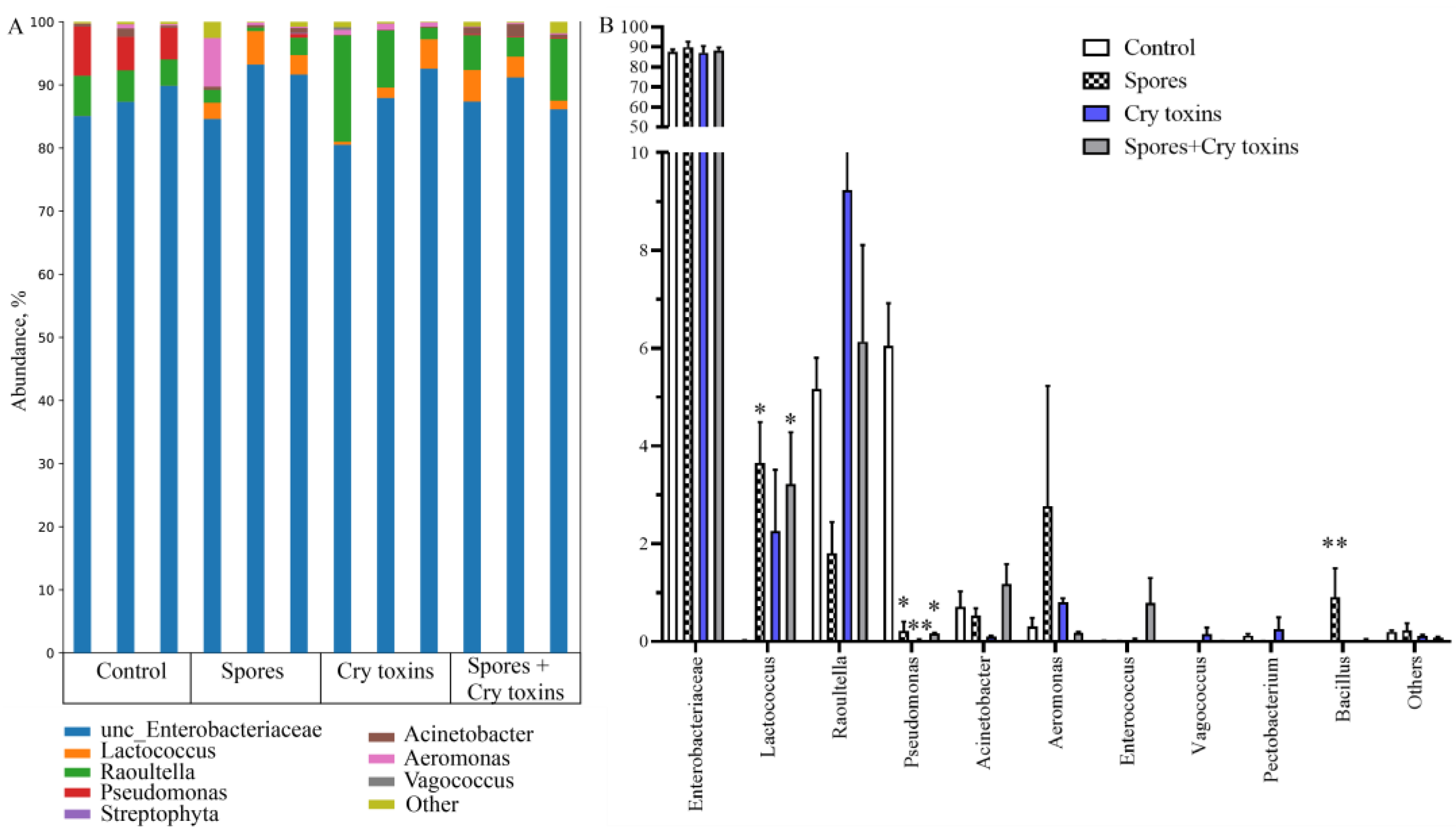
2.4. Microbiota of CPB Larvae Post Bt Treatments
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Insects and Bacteria
5.2. Enzymatic Activity and Redox Balance in Midgut of CPB Larvae
5.3. QRT–PCR Analysis of Insect Immunity and Stress-Related Gene Expression in the Midgut
5.4. 16S (V3/V4) rDNA Bacterial Diversity Analysis of Midgut and Cadaver Community of CPB
5.5. Data Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sanahuja, G.; Banakar, R.; Twyman, R.M.; Capell, T.; Christou, P. Bacillus thuringiensis: A century of research, development and commercial applications. Plant Biotechnol. J. 2011, 9, 283–300. [Google Scholar] [CrossRef] [Green Version]
- Bravo, A.; Gill, S.S.; Soberon, M. Bacillus thuringiensis: Mechanisms and use. In Comprehensive Molecular Insect Science; Gilbert, L.I., Kostas, I., Gill, S.S., Eds.; Elsevier: Amsterdam, The Netherlands, 2005; Volume 6, pp. 175–205. [Google Scholar]
- Soberon, M.; Gill, S.S.; Bravo, A. Signaling versus punching hole: How do Bacillus thuringiensis toxins kill insect midgut cells? Cell. Mol. Life Sci. 2009, 66, 1337–1349. [Google Scholar] [CrossRef]
- Zhang, X.B.; Candas, M.; Griko, N.B.; Taussig, R.; Bulla, L.A. A mechanism of cell death involving an adenylyl cyclase/PKA signaling pathway is induced by the Cry1Ab toxin of Bacillus thuringiensis. Proc. Natl. Acad. Sci. USA 2006, 103, 9897–9902. [Google Scholar] [CrossRef] [Green Version]
- Brivio, M.F.; Mastore, M. When appearance misleads: The role of the entomopathogen surface in the relationship with its host. Insects 2020, 11, 387. [Google Scholar] [CrossRef]
- Nielsen-LeRoux, C.; Gaudriault, S.; Ramarao, N.; Lereclus, D.; Givaudan, A. How the insect pathogen bacteria Bacillus thuringiensis and Xenorhabdus/Photorhabdus occupy their hosts. Curr. Opin. Microbiol. 2012, 15, 220–231. [Google Scholar] [CrossRef]
- Chakroun, M.; Banyuls, N.; Bel, Y.; Escriche, B.; Ferré, J. Bacterial Vegetative Insecticidal Proteins (Vip) from Entomopathogenic Bacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 329–350. [Google Scholar] [CrossRef] [Green Version]
- Donovan, W.P.; Engleman, J.T.; Donovan, J.C.; Baum, J.A.; Bunkers, G.J.; Chi, D.J.; Clinton, W.P.; English, L.; Heck, G.R.; Ilagan, O.M.; et al. Discovery and characterization of Sip1A: A novel secreted protein from Bacillus thuringiensis with activity against coleopteran larvae. Appl. Microbiol. Biotechnol. 2006, 72, 713–719. [Google Scholar] [CrossRef]
- Ehling-Schulz, M.; Lereclus, D.; Koehler, T.M. The Bacillus cereus Group: Bacillus Species with Pathogenic Potential. Microbiol. Spectr. 2019, 7, 7. [Google Scholar] [CrossRef]
- Malovichko, Y.V.; Nizhnikov, A.A.; Antonets, K.S. Repertoire of the bacillus thuringiensis virulence factors unrelated to major classes of protein toxins and its role in specificity of host-pathogen interactions. Toxins 2019, 11, 347. [Google Scholar] [CrossRef] [Green Version]
- Buisson, C.; Gohar, M.; Huillet, E.; Nielsen-LeRoux, C. Bacillus thuringiensis spores and vegetative bacteria: Infection capacity and role of the virulence regulon plcr following intrahaemocoel injection of galleria mellonella. Insects 2019, 10, 129. [Google Scholar] [CrossRef] [Green Version]
- Consentino, L.; Rejasse, A.; Crapart, N.; Bevilacqua, C.; Nielsen-LeRoux, C. Laser capture microdissection to study Bacillus cereus iron homeostasis gene expression during Galleria mellonella in vivo gut colonization. Virulence 2021, 12, 2104–2121. [Google Scholar] [CrossRef]
- Schoville, S.D.; Chen, Y.H.; Andersson, M.N.; Benoit, J.B.; Bhandari, A.; Bowsher, J.H.; Brevik, K.; Cappelle, K.; Chen, M.J.M.; Childers, A.K.; et al. A model species for agricultural pest genomics: The genome of the Colorado potato beetle, Leptinotarsa decemlineata (Coleoptera: Chrysomelidae). Sci. Rep. 2018, 8, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Weber, D.C. Colorado beetle: Pest on the move. Pestic. Outlook 2003, 14, 256–259. [Google Scholar] [CrossRef]
- Balaško, M.K.; Mikac, K.M.; Bažok, R.; Lemic, D. Modern techniques in colorado potato beetle (Leptinotarsa decemlineata say) control and resistance management: History review and future perspectives. Insects 2020, 11, 581. [Google Scholar] [CrossRef]
- Alyokhin, A.V.; Ferro, D.N. Relative fitness of Colorado potato beetle (Coleoptera: Chrysomelidae) resistant and susceptible to the Bacillus thuringiensis Cry3A toxin. J. Econ. Entomol. 1999, 92, 510–515. [Google Scholar] [CrossRef] [Green Version]
- Christou, P.; Capell, T.; Kohli, A.; Gatehouse, J.A.; Gatehouse, A.M.R. Recent developments and future prospects in insect pest control in transgenic crops. Trends Plant Sci. 2006, 11, 302–308. [Google Scholar] [CrossRef]
- Sexson, D.L.; Wyman, J.A. Effect of crop rotation distance on populations of Colorado potato beetle (Coleoptera: Chrysomelidae): Development of areawide Colorado potato beetle pest management strategies. J. Econ. Entomol. 2005, 98, 716–724. [Google Scholar] [CrossRef]
- Pinos, D.; Andrés-Garrido, A.; Ferré, J.; Hernández-Martínez, P. Response Mechanisms of Invertebrates to Bacillus thuringiensis and Its Pesticidal Proteins. Microbiol. Mol. Biol. Rev. 2021, 85, 85. [Google Scholar] [CrossRef]
- Jurat-Fuentes, J.L.; Heckel, D.G.; Ferré, J. Mechanisms of Resistance to Insecticidal Proteins from Bacillus thuringiensis. Annu. Rev. Entomol. 2021, 66, 121–140. [Google Scholar] [CrossRef]
- Dubovskiy, I.M.; Grizanova, E.V.; Whitten, M.M.A.; Mukherjee, K.; Greig, C.; Alikina, T.; Kabilov, M.; Vilcinskas, A.; Glupov, V.V.; Butt, T.M. Immuno-physiological adaptations confer wax moth Galleria mellonella resistance to Bacillus thuringiensis. Virulence 2016, 7, 860–870. [Google Scholar] [CrossRef] [Green Version]
- Dubovskiy, I.M.; Kryukova, N.A.; Glupov, V.V.; Ratcliffe, N.A. Encapsulation and nodulation in insects. Invertebr. Surviv. J. 2016, 13. [Google Scholar] [CrossRef]
- Saikhedkar, N.; Summanwar, A.; Joshi, R.; Giri, A. Cathepsins of lepidopteran insects: Aspects and prospects. Insect Biochem. Mol. Biol. 2015, 64, 51–59. [Google Scholar] [CrossRef]
- Grizanova, E.V.; Krytsyna, T.I.; Surcova, V.S.; Dubovskiy, I.M. The role of midgut nonspecific esterase in the susceptibility of Galleria mellonella larvae to Bacillus thuringiensis. J. Invertebr. Pathol. 2019, 166, 107208. [Google Scholar] [CrossRef]
- Ma, G.; Roberts, H.; Sarjan, M.; Featherstone, N.; Lahnstein, J.; Akhurst, R.; Schmidt, O. Is the mature endotoxin Cry1Ac from Bacillus thuringiensis inactivated by a coagulation reaction in the gut lumen of resistant Helicoverpa armigera larvae? Insect Biochem. Mol. Biol. 2005, 35, 729–739. [Google Scholar] [CrossRef]
- Castagnola, A.; Jurat-Fuentes, J.L. Intestinal regeneration as an insect resistance mechanism to entomopathogenic bacteria. Curr. Opin. Insect Sci. 2016, 15, 104–110. [Google Scholar] [CrossRef] [Green Version]
- Zhai, Z.; Boquete, J.P.; Lemaitre, B. Cell-Specific Imd-NF-κB Responses Enable Simultaneous Antibacterial Immunity and Intestinal Epithelial Cell Shedding upon Bacterial Infection. Immunity 2018, 48, 897–910.e7. [Google Scholar] [CrossRef] [Green Version]
- Buchon, N.; Broderick, N.A.; Lemaitre, B. Gut homeostasis in a microbial world: Insights from Drosophila melanogaster. Nat. Rev. Microbiol. 2013, 11, 615–626. [Google Scholar] [CrossRef] [Green Version]
- Felton, G.W.; Summers, C.B. Antioxidant systems in insects. Arch. Insect Biochem. Physiol. 1995, 29, 187–197. [Google Scholar] [CrossRef]
- Li, X.; Schuler, M.A.; Berenbaum, M.R. Molecular Mechanisms of Metabolic Resistance to Synthetic and Natural Xenobiotics. Annu. Rev. Entomol. 2007, 52, 231–253. [Google Scholar] [CrossRef]
- Hiraiwa, M.; Martin, B.M.; Kishimoto, Y.; Conner, G.E.; Tsuji, S.; O’Brien, J.S. Lysosomal proteolysis of prosaposin, the precursor of saposins (sphingolipid activator proteins): Its mechanism and inhibition by ganglioside. Arch. Biochem. Biophys. 1997, 341, 17–24. [Google Scholar] [CrossRef]
- Udupi, V.; Rice-evans, C. Thiol compounds as protective agents in erythrocytes under oxidative stress. Free Radic. Res. 1992, 16, 315–323. [Google Scholar] [CrossRef]
- Grizanova, E.V.; Semenova, A.D.; Komarov, D.A.; Chertkova, E.A.; Slepneva, I.A.; Dubovskiy, I.M. Maintenance of redox balance by antioxidants in hemolymph of the greater wax moth Galleria mellonella larvae during encapsulation response. Arch. Insect Biochem. Physiol. 2018, 98, e21460. [Google Scholar] [CrossRef]
- Li, S.; de Mandal, S.; Xu, X.; Jin, F. The tripartite interaction of host immunity-bacillus thuringiensis infection-gut microbiota. Toxins 2020, 12, 514. [Google Scholar] [CrossRef]
- Caccia, S.; Di Lelio, I.; La Storia, A.; Marinelli, A.; Varricchio, P.; Franzetti, E.; Banyuls, N.; Tettamanti, G.; Casartelli, M.; Giordana, B.; et al. Midgut microbiota and host immunocompetence underlie Bacillus thuringiensis killing mechanism. Proc. Natl. Acad. Sci. USA 2016, 113, 9486–9491. [Google Scholar] [CrossRef] [Green Version]
- Patil, C.D.; Borase, H.P.; Salunke, B.K.; Patil, S.V. Alteration in Bacillus thuringiensis toxicity by curing gut flora: Novel approach for mosquito resistance management. Parasitol. Res. 2013, 112, 3283–3288. [Google Scholar] [CrossRef] [PubMed]
- Li, R.S.; Jarrett, P.; Burges, H.D. Importance of spores, crystals, and δ-endotoxins in the pathogenicity of different varieties of Bacillus thuringiensis in Galleria mellonella and Pieris brassicae. J. Invertebr. Pathol. 1987, 50, 277–284. [Google Scholar] [CrossRef]
- Fedhila, S.; Buisson, C.; Dussurget, O.; Serror, P.; Glomski, I.J.; Liehl, P.; Lereclus, D.; Nielsen-LeRoux, C. Comparative analysis of the virulence of invertebrate and mammalian pathogenic bacteria in the oral insect infection model Galleria mellonella. J. Invertebr. Pathol. 2010, 103, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Miyasono, M.; Inagaki, S.; Yamamoto, M.; Ohba, K.; Ishiguro, T.; Takeda, R.; Hayashi, Y. Enhancement of δ-Endotoxin Activity by Toxin-Free Spore of Bacillus thuringiensis against the Diamondback Moth, Plutella xylostella. J. Invertebr. Pathol. 1994, 63, 111–112. [Google Scholar] [CrossRef]
- Johnson, D.E.; McGaughey, W.H. Contribution of Bacillus thuringiensis spores to toxicity of purified cry proteins towards indianmeal moth larvae. Curr. Microbiol. 1996, 33, 54–59. [Google Scholar] [CrossRef]
- Kalmykova, G.; Burtseva, L.; Milne, R.; Van Frankenhuyzen, K. Activity of spores and extracellular proteins from six Cry+ strains and a Cry- strain of Bacillus thuringiensis subsp. kurstaki against the western spruce budworm, Choristoneura occidentalis (Lepidoptera: Tortricidae). Can. J. Microbiol. 2009, 55, 536–543. [Google Scholar] [CrossRef]
- Liu, Y.B.; Tabashnik, B.E.; Moar, W.J.; Smith, R.A. Synergism between Bacillus thuringiensis spores and toxins against resistant and susceptible diamondback moths (Plutella xylostella). Appl. Environ. Microbiol. 1998, 64, 385–1389. [Google Scholar] [CrossRef] [Green Version]
- Tetreau, G. Interaction between insects, toxins, and bacteria: Have we been wrong so far? Toxins 2018, 10, 281. [Google Scholar] [CrossRef] [Green Version]
- Du, C.; Chan, W.C.; McKeithan, T.W.; Nickerson, K.W. Surface display of recombinant proteins on Bacillus thuringiensis spores. Appl. Environ. Microbiol. 2005, 71, 3337–3341. [Google Scholar] [CrossRef] [Green Version]
- Charlton, S.; Moir, A.J.G.; Baillie, L.; Moir, A. Characterization of the exosporium of Bacillus cereus. J. Appl. Microbiol. 1999, 87, 241–245. [Google Scholar] [CrossRef] [PubMed]
- Fedhila, S.; Nel, P.; Lereclus, D. The InhA2 metalloprotease of Bacillus thuringiensis strain 407 is required for pathogenicity in insects infected via the oral route. J. Bacteriol. 2002, 184, 3296–3304. [Google Scholar] [CrossRef] [Green Version]
- Dalhammar, G.; Steiner, H. Characterization of inhibitor A, a protease from Bacillus thuringiensis which degrades attacins and cecropins, two classes of antibacterial proteins in insects. Eur. J. Biochem. 1984, 139, 247–252. [Google Scholar] [CrossRef]
- Miyoshi, S.I.; Shinoda, S. Microbial metalloproteases and pathogenesis. Microbes Infect. 2000, 2, 91–98. [Google Scholar] [CrossRef]
- Geng, J.; Jiang, J.; Shu, C.; Wang, Z.; Song, F.; Geng, L.; Duan, J.; Zhang, J. Bacillus thuringiensis vip1 functions as a receptor of vip2 toxin for binary insecticidal activity against holotrichia parallela. Toxins 2019, 11, 440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, J.; Tong, Z.; Zhan, Y.; Buisson, C.; Song, F.; He, K.; Nielsen-LeRoux, C.; Guo, S. A Bacillus thuringiensis Chitin-Binding Protein is Involved in Insect Peritrophic Matrix Adhesion and Takes Part in the Infection Process. Toxins 2020, 12, 252. [Google Scholar] [CrossRef]
- Capo, F.; Wilson, A.; Di Cara, F. The intestine of Drosophila melanogaster: An emerging versatile model system to study intestinal epithelial homeostasis and host-microbial interactions in humans. Microorganisms 2019, 7, 336. [Google Scholar] [CrossRef] [Green Version]
- Dubovskiy, I.M.; Martemyanov, V.V.; Vorontsova, Y.L.; Rantala, M.J.; Gryzanova, E.V.; Glupov, V.V. Effect of bacterial infection on antioxidant activity and lipid peroxidation in the midgut of Galleria mellonella L. larvae (Lepidoptera, Pyralidae). Comp. Biochem. Physiol.-C Toxicol. Pharmacol. 2008, 48, 1–5. [Google Scholar] [CrossRef]
- Sajjadian, S.M.; Kim, Y. Dual Oxidase-Derived Reactive Oxygen Species Against Bacillus thuringiensis and Its Suppression by Eicosanoid Biosynthesis Inhibitors. Front. Microbiol. 2020, 11, 528. [Google Scholar] [CrossRef]
- Yaroslavtseva, O.N.; Dubovskiy, I.M.; Khodyrev, V.P.; Duisembekov, B.A.; Kryukov, V.Y.; Glupov, V.V. Immunological mechanisms of synergy between fungus Metarhizium robertsii and bacteria Bacillus thuringiensis ssp. morrisoni on Colorado potato beetle larvae. J. Insect Physiol. 2017, 96, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Gunning, R.V.; Dang, H.T.; Kemp, F.C.; Nicholson, I.C.; Moores, G.D. New resistance mechanism in Helicoverpa armigera threatens transgenic crops expressing Bacillus thuringiensis Cry1Ac toxin. Appl. Environ. Microbiol. 2005, 71, 2558–2563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chertkova, E.A.; Grizanova, E.V.; Dubovskiy, I.M. Bacterial and fungal infections induce bursts of dopamine in the haemolymph of the Colorado potato beetle Leptinotarsa decemlineata and greater wax moth Galleria mellonella. J. Invertebr. Pathol. 2018, 153, 203–206. [Google Scholar] [CrossRef] [PubMed]
- García-Robles, I.; De Loma, J.; Capilla, M.; Roger, I.; Boix-Montesinos, P.; Carrión, P.; Vicente, M.; López-Galiano, M.J.; Real, M.D.; Rausell, C. Proteomic insights into the immune response of the Colorado potato beetle larvae challenged with Bacillus thuringiensis. Dev. Comp. Immunol. 2020, 104, 103525. [Google Scholar] [CrossRef]
- Grizanova, E.V.; Dubovskiy, I.M.; Whitten, M.M.A.; Glupov, V.V. Contributions of cellular and humoral immunity of Galleria mellonella larvae in defence against oral infection by bacillus thuringiensis. J. Invertebr. Pathol. 2014, 119, 40–46. [Google Scholar] [CrossRef]
- Zwick, R.K.; Ohlstein, B.; Klein, O.D. Intestinal renewal across the animal kingdom: Comparing stem cell activity in mouse and Drosophila. Am. J. Physiol. Liver Physiol. 2019, 316, G313–G322. [Google Scholar] [CrossRef]
- Wu, K.; Yang, B.; Huang, W.; Dobens, L.; Song, H.; Ling, E. Gut immunity in Lepidopteran insects. Dev. Comp. Immunol. 2016, 64, 65–74. [Google Scholar] [CrossRef]
- Terra, W.R.; Dias, R.O.; Oliveira, P.L.; Ferreira, C.; Venancio, T.M. Transcriptomic analyses uncover emerging roles of mucins, lysosome/secretory addressing and detoxification pathways in insect midguts. Curr. Opin. Insect Sci. 2018, 29, 34–40. [Google Scholar] [CrossRef]
- Griffitts, J.S.; Aroian, R. V Many roads to resistance: How invertebrates adapt to Bt toxins. Bioessays 2005, 27, 614–624. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; You, M.; Rao, X.J.; Yu, X.Q. Insect C-type lectins in innate immunity. Dev. Comp. Immunol. 2018, 83, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.X.; Zhu, B.J.; Tang, L.; Sun, Y.; Chen, C.; Nadeem Abbas, M.; Wang, L.; Qian, C.; Wei, G.Q.; Liu, C. liang Cathepsin O is involved in the innate immune response and metamorphosis of Antheraea pernyi. J. Invertebr. Pathol. 2017, 150, 6–14. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Cattaneo, A.; Gobert, F.X.; Müller, M.; Toscano, F.; Flores, M.; Lescure, A.; Del Nery, E.; Benaroch, P. Cleavage of Toll-like receptor 3 by cathepsins B and H is essential for signaling. Proc. Natl. Acad. Sci. USA 2012, 109, 9053–9058. [Google Scholar] [CrossRef] [Green Version]
- Nunes, C.; Sucena, É.; Koyama, T. Endocrine regulation of immunity in insects. FEBS J. 2020, 288, 3928–3947. [Google Scholar] [CrossRef]
- Rantala, M.J.; Dubovskiy, I.M.; Pölkki, M.; Krama, T.; Contreras-Garduño, J.; Krams, I.A. Effect of juvenile hormone on resistance against entomopathogenic fungus metharizium robertsii differs between sexes. J. Fungi 2020, 6, 298. [Google Scholar] [CrossRef] [PubMed]
- van Frankenhuyzen, K.; Liu, Y.H.; Tonon, A. Interactions between Bacillus thuringiensis subsp kurstaki HD-1 and midgut bacteria in larvae of gypsy moth and spruce budworm. J. Invertebr. Pathol. 2010, 103, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Polenogova, O.V.; Noskov, Y.A.; Yaroslavtseva, O.N.; Kryukova, N.A.; Alikina, T.; Klementeva, T.N.; Andrejeva, J.; Khodyrev, V.P.; Kabilov, M.R.; Kryukov, V.Y.; et al. Influence of Bacillus thuringiensis and avermectins on gut physiology and microbiota in Colorado potato beetle: Impact of enterobacteria on susceptibility to insecticides. PLoS ONE 2021, 16, e0248704. [Google Scholar] [CrossRef]
- Sarvari, M.; Mikani, A.; Mehrabadi, M. The innate immune gene Relish and Caudal jointly contribute to the gut immune homeostasis by regulating antimicrobial peptides in Galleria mellonella. Dev. Comp. Immunol. 2020, 110, 103732. [Google Scholar] [CrossRef]
- Broderick, N.A.; Robinson, C.J.; McMahon, M.D.; Holt, J.; Handelsman, J.; Raffa, K.F. Contributions of gut bacteria to Bacillus thuringiensis-induced mortality vary across a range of Lepidoptera. BMC Biol. 2009, 7, 11. [Google Scholar] [CrossRef] [Green Version]
- Thomas, W.E.; Ellar, D.J. Bacillus thuringiensis var israelensis crystal δ-endotoxin: Effects on insect and mammalian cells in vitro and in vivo. J. Cell Sci. 1983, 60, 181–197. [Google Scholar] [CrossRef] [PubMed]
- Prabhakaran, S.K.; Kamble, S.T. Activity and electrophoretic characterization of esterases in insecticide-resistant and susceptible strains of German cockroach (Dictyoptera: Blattellidae). J. Econ. Entomol. 1993, 86, 1009–1013. [Google Scholar] [CrossRef] [Green Version]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S transferases. The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef]
- Butt, T.M.; Greenfield, B.P.J.; Greig, C.; Maffeis, T.G.G.; Taylor, J.W.D.; Piasecka, J.; Dudley, E.; Abdulla, A.; Dubovskiy, I.M.; Garrido-Jurado, I.; et al. Metarhizium anisopliae pathogenesis of mosquito larvae: A verdict of accidental death. PLoS ONE 2013, 8, e81686. [Google Scholar] [CrossRef] [Green Version]
- Rael, L.T.; Thomas, G.W.; Craun, M.L.; Curtis, C.G.; Bar-Or, R.; Bar-Or, D. Lipid peroxidation and the thiobarbituric acid assay: Standardization of the assay when using saturated and unsaturated fatty acids. J. Biochem. Mol. Biol. 2004, 37, 749–752. [Google Scholar] [CrossRef]
- Bar-Or, D.; Rael, L.T.; Lau, E.P.; Rao, N.K.R.; Thomas, G.W.; Winkler, J.V.; Yukl, R.L.; Kingston, R.G.; Curtis, C.G. An analog of the human albumin N-terminus (Asp-Ala-His-Lys) prevents formation of copper-induced reactive oxygen species. Biochem. Biophys. Res. Commun. 2001, 284, 856–862. [Google Scholar] [CrossRef]
- Khramtsov, V.V.; Yelinova (Popova), V.I.; Weiner, L.M.; Berezina, T.A.; Martin, V.V.; Volodarsky, L.B. Quantitative determination of SH groups in low- and high-molecular-weight compounds by an electron spin resonance method. Anal. Biochem. 1989, 182, 58–63. [Google Scholar] [CrossRef]
- Butt, T.M.; Coates, C.J.; Dubovskiy, I.M.; Ratcliffe, N.A. Entomopathogenic Fungi: New Insights into Host-Pathogen Interactions. Adv. Genet. 2016, 94, 307–364. [Google Scholar] [CrossRef]
- Rozen, S.; Skaletsky, H. Primer3 on the WWW for general users and for biologist programmers. Methods Mol. Biol. 2000, 132, 365–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuker, M. Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acids Res. 2003, 31, 3406–3415. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Matyugina, E.; Belkova, N.; Borzenko, S.; Lukyanov, P.; Kabilov, M.; Baturina, O.; Van Kley, A.M.; Nalian, A.; Ptitsyn, A. Structure and diversity dynamics of microbial communities at day and night: Investigation of meromictic Lake Doroninskoe, Transbaikalia, Russia. J. Oceanol. Limnol. 2018, 36, 1978–1992. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Edgar, R. UNOISE2: Improved error-correction for Illumina 16S and ITS amplicon sequencing. bioRxiv 2016. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R. SINTAX: A simple non-Bayesian taxonomy classifier for 16S and ITS sequences. bioRxiv 2016. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robertson, J.L.; Preisler, H.K. Pesticide Bioassays with Arthropods; CRC Press: Boca Raton, FL, USA, 1992. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spores | Cry3A Toxins | Spores + Cry3A Toxins | |
|---|---|---|---|
| Control | X2 (1) = 0.988, p = 0.3201 | X2 (1) = 11.64, p = 0.0006 HR = 3.9 (95% CI, 1.8–8.8) | X2 (1) = 46.75, p < 0.0001 HR = 6.3 (95% CI, 3.7–10.6) |
| Spores | - | X2 (1) = 6.224, p = 0.0126 HR = 2.6 (95% CI, 1.2–5.5) | X2 (1) = 36.05, p < 0.0001 HR = 4.9 (95% CI, 2.9–8.2) |
| Cry3A toxins | - | - | X2 (1) = 15.92, p < 0.0001 HR = 2.6 (95% CI, 1.7–4.1) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dubovskiy, I.M.; Grizanova, E.V.; Tereshchenko, D.; Krytsyna, T.I.; Alikina, T.; Kalmykova, G.; Kabilov, M.; Coates, C.J. Bacillus thuringiensis Spores and Cry3A Toxins Act Synergistically to Expedite Colorado Potato Beetle Mortality. Toxins 2021, 13, 746. https://doi.org/10.3390/toxins13110746
Dubovskiy IM, Grizanova EV, Tereshchenko D, Krytsyna TI, Alikina T, Kalmykova G, Kabilov M, Coates CJ. Bacillus thuringiensis Spores and Cry3A Toxins Act Synergistically to Expedite Colorado Potato Beetle Mortality. Toxins. 2021; 13(11):746. https://doi.org/10.3390/toxins13110746
Chicago/Turabian StyleDubovskiy, Ivan M., Ekaterina V. Grizanova, Daria Tereshchenko, Tatiana I. Krytsyna, Tatyana Alikina, Galina Kalmykova, Marsel Kabilov, and Christopher J. Coates. 2021. "Bacillus thuringiensis Spores and Cry3A Toxins Act Synergistically to Expedite Colorado Potato Beetle Mortality" Toxins 13, no. 11: 746. https://doi.org/10.3390/toxins13110746
APA StyleDubovskiy, I. M., Grizanova, E. V., Tereshchenko, D., Krytsyna, T. I., Alikina, T., Kalmykova, G., Kabilov, M., & Coates, C. J. (2021). Bacillus thuringiensis Spores and Cry3A Toxins Act Synergistically to Expedite Colorado Potato Beetle Mortality. Toxins, 13(11), 746. https://doi.org/10.3390/toxins13110746