


Ammonium Uptake, Mediated by Ammonium Transporters, Mitigates Manganese Toxicity in Duckweed, Spirodela polyrhiza

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
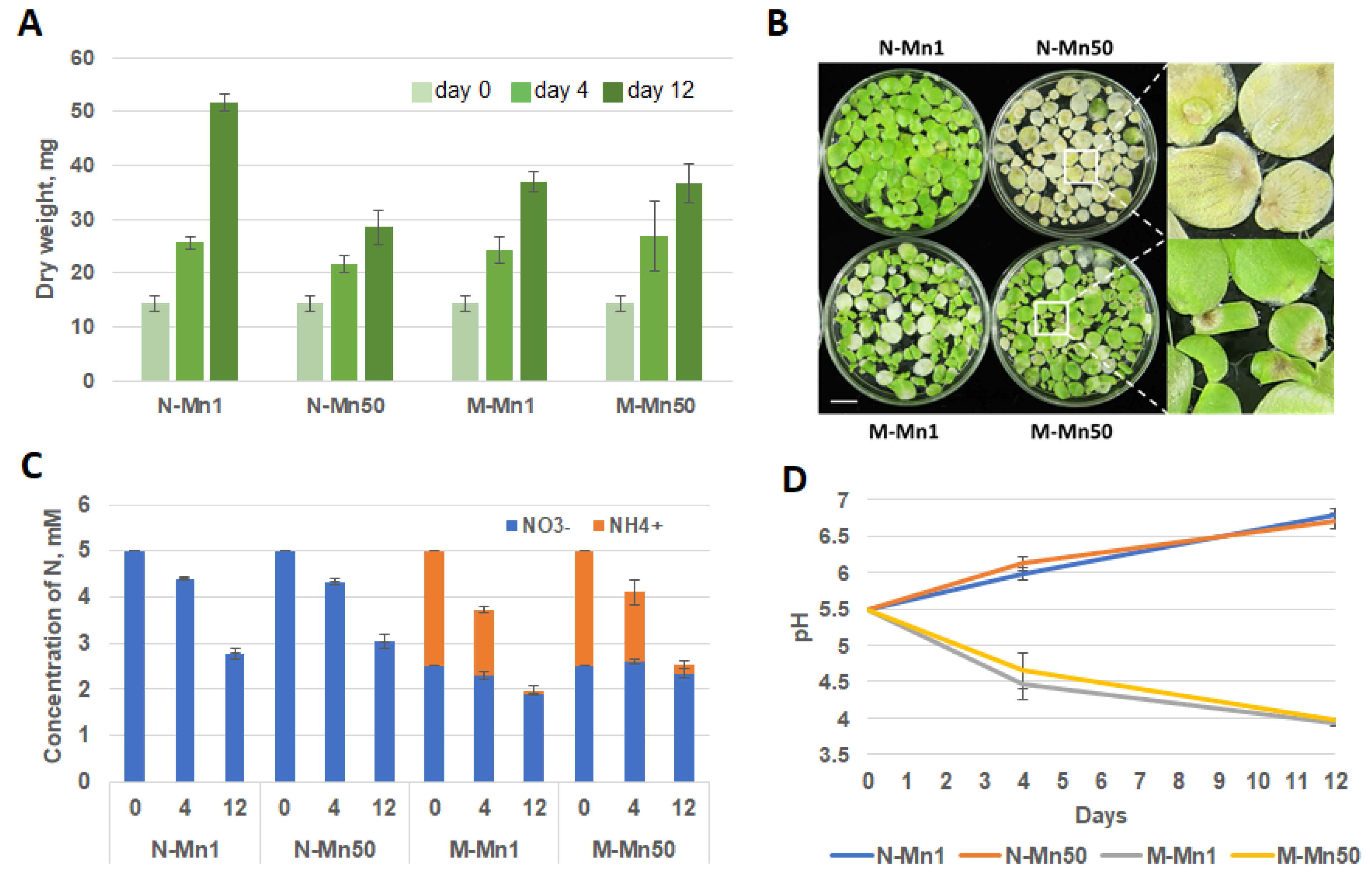
2.1. Parameters of the Experimental System: Dynamics of Biomass Accumulation, Nitrogen Uptake, Changes in Medium pH, during 12-Days of Duckweed Cultivation
2.2. Genome of S. polyrhiza Containes Four Genes Coding for Ammonium Transporters
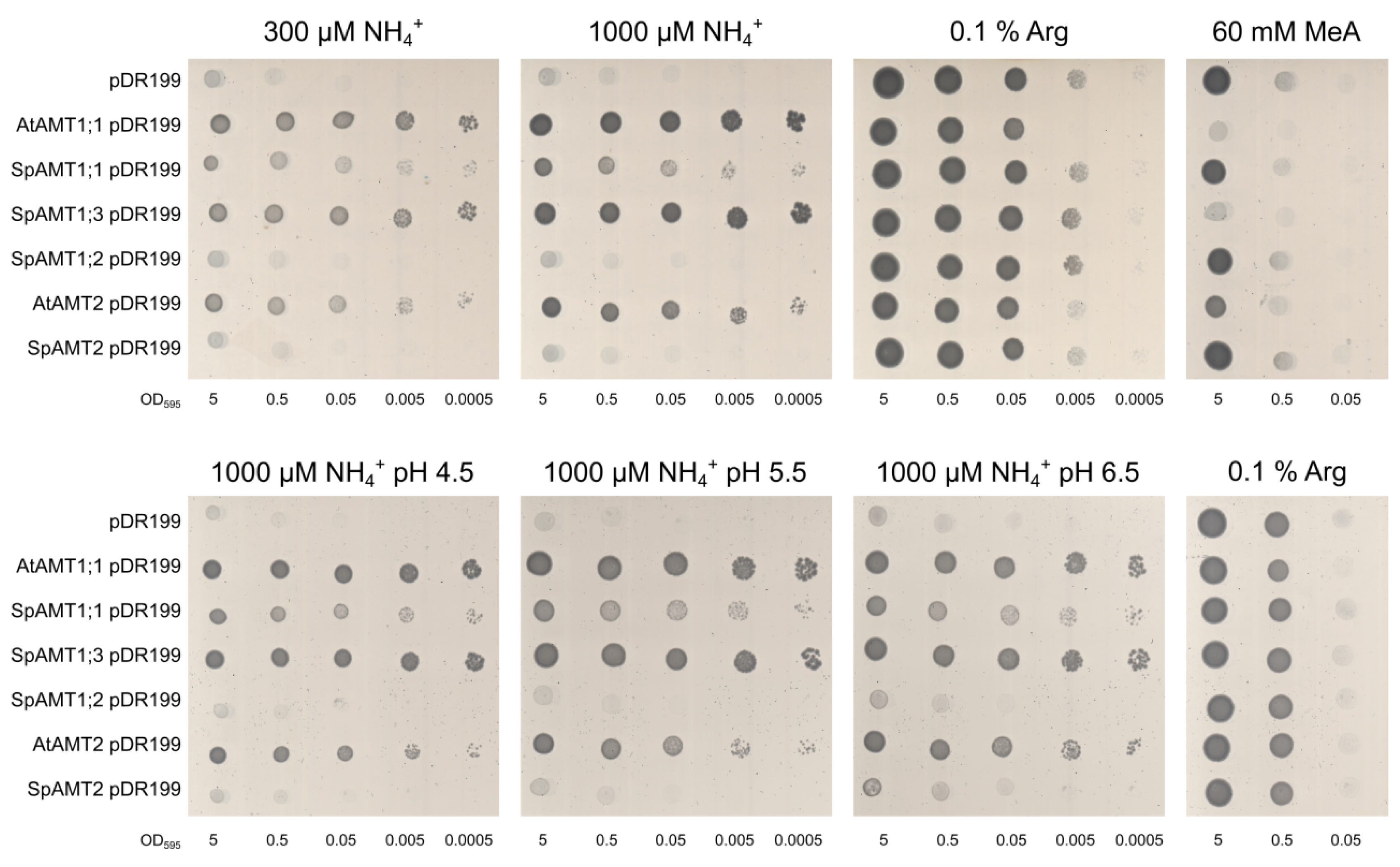
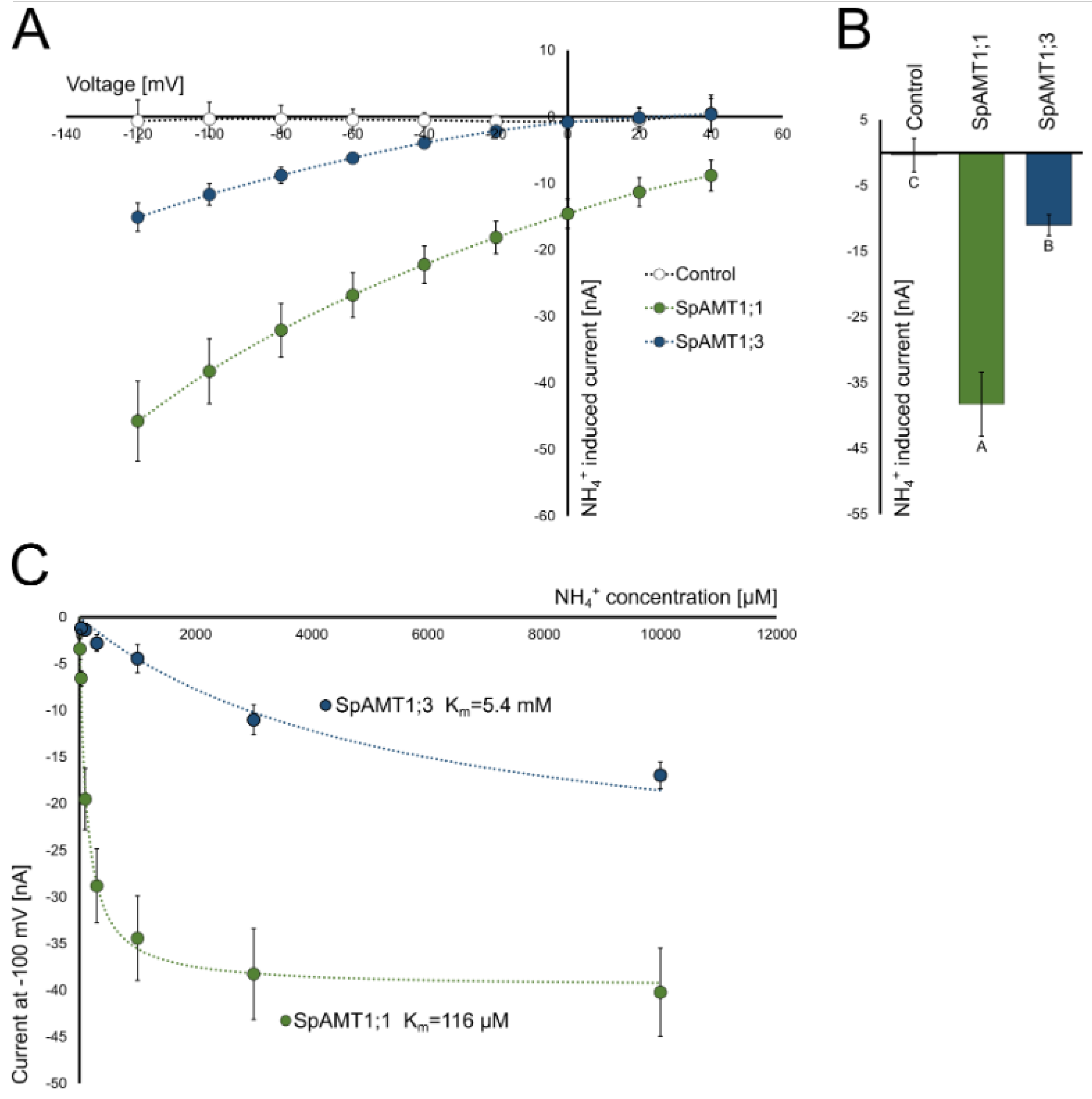
2.3. Functional Activity of the Ammonium Transporters in Yeast and Xenopus Oocytes Systems
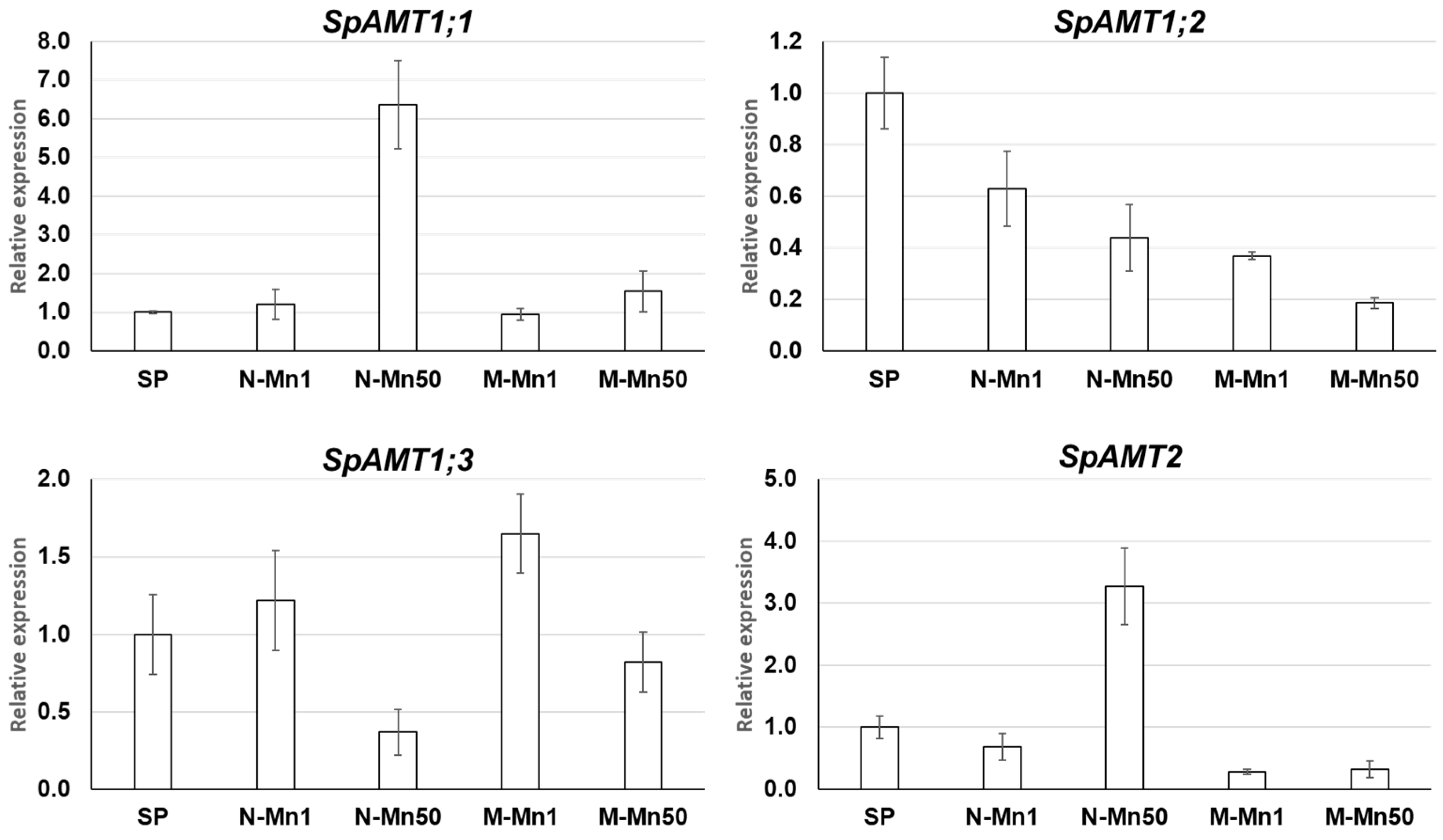
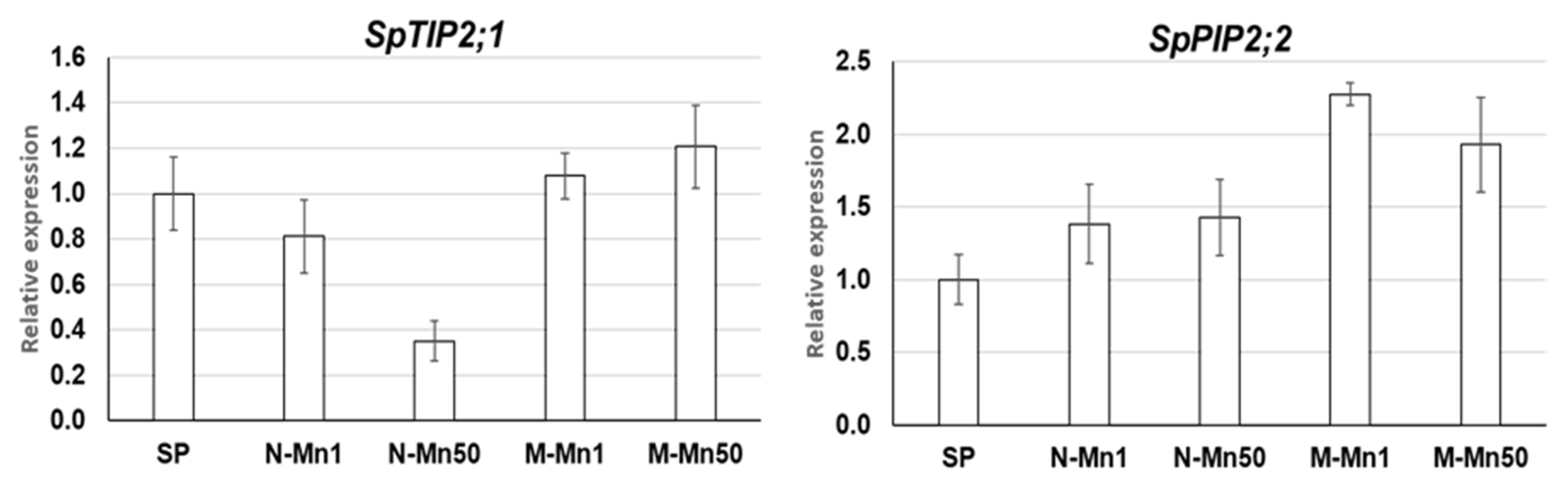
2.4. Expression Profiling of S. polyrhiza AMT Genes by qRT-PCR
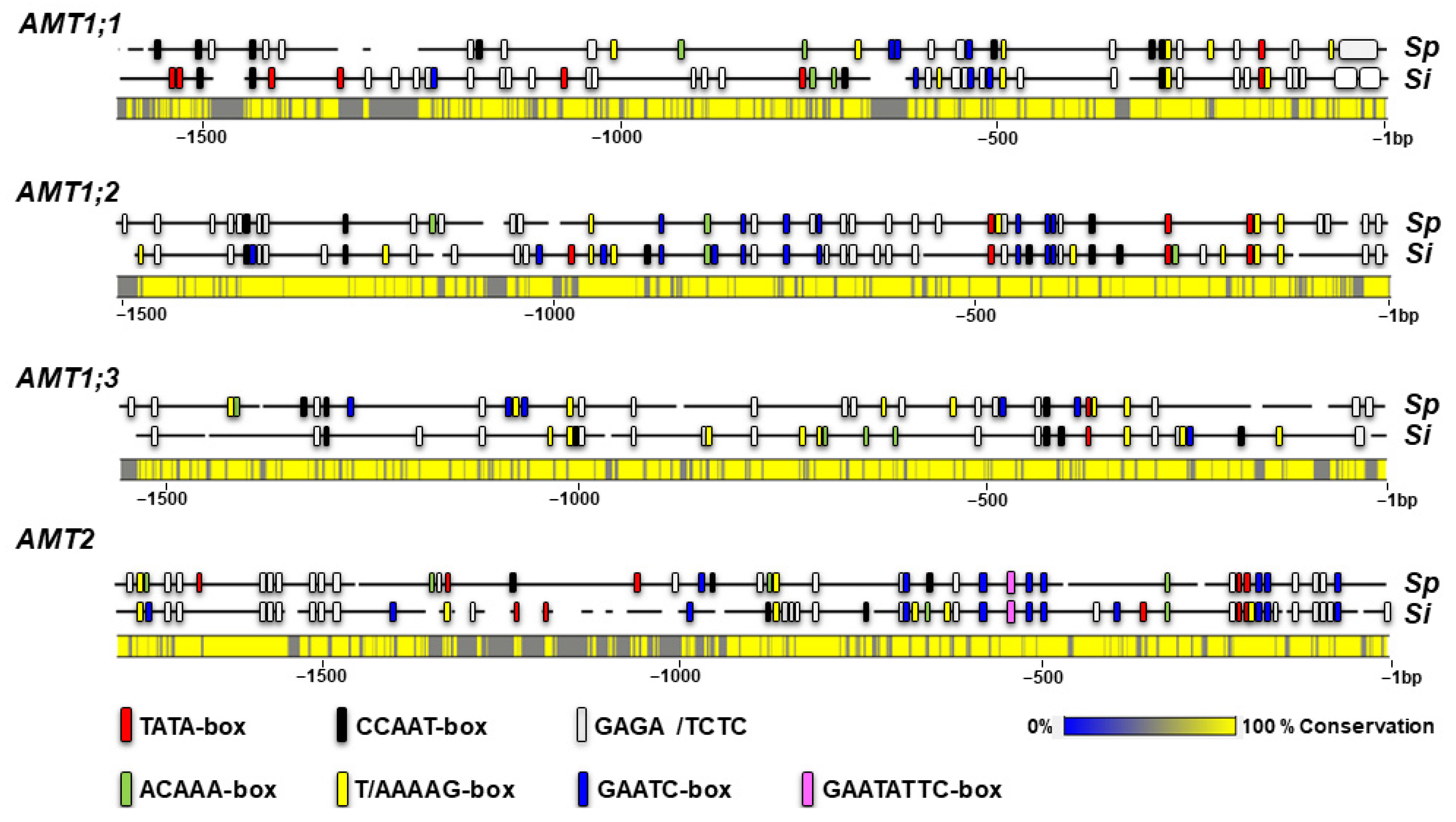
2.5. Regulatory Cis-Elements in the Promoters of Spirodela AMT Genes
3. Discussion
3.1. Alleviated Mn Stress Symptoms for S. polyrhiza Compared with
3.2. Ammonium Transporter (AMT) Genes in the Genome of S. polyrhiza
3.3. SpAMT1;1 and SpAMT1;3 Demonstrate Clear Activity in Yeast and Oocytes
3.4. The Four S. polyrhiza AMT Genes Show Different Expression Patterns in Response to Nitrogen Source and Mn
3.5. Distinctive Promoter Elements in the AMT Genes in S. polyrhiza
4. Summary
5. Materials and Methods
5.1. Plant Material
5.2. Duckweed Cultivation Parameters
5.3. Determination of N Concentration
5.4. Cloning and Analysis of S. polyrhiza Genes Encoding Ammonium Transporters
5.5. Phylogenetic Analysis
5.6. Construction of Expression Vectors and Testing AMTs Functionality in Yeast and Xenopus oocytes
5.7. Analysis of AMT Genes Expression in S. polyrhiza by RT-qPCR
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Scheible, W.-R.; Morcuende, R.; Czechowski, T.; Fritz, C.; Osuna, D.; Palacios-Rojas, N.; Schindelasch, D.; Thimm, O.; Udvardi, M.K.; Stitt, M. Genome-Wide Reprogramming of Primary and Secondary Metabolism, Protein Synthesis, Cellular Growth Processes, and the Regulatory Infrastructure of Arabidopsis in Response to Nitrogen. Plant Physiol. 2004, 136, 2483–2499. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Fan, X.; Miller, A.J. Plant Nitrogen Assimilation and Use Efficiency. Annu. Rev. Plant Biol. 2012, 63, 153–182. [Google Scholar] [CrossRef] [PubMed]
- Saloner, A.; Bernstein, N. Nitrogen Source Matters: High NH4/NO3 Ratio Reduces Cannabinoids, Terpenoids, and Yield in Medical Cannabis. Front. Plant Sci. 2022, 13, 830224. [Google Scholar] [CrossRef]
- Bloom, A.J. The Increasing Importance of Distinguishing among Plant Nitrogen Sources. Curr. Opin. Plant Biol. 2015, 25, 10–16. [Google Scholar] [CrossRef]
- Chen, P.; Xu, J.; Zhang, Z.; Nie, T. “Preferential” Ammonium Uptake by Rice Does Not Always Turn into Higher N Recovery of Fertilizer Sources under Water-Saving Irrigation. Agric. Water Manag. 2022, 272, 107867. [Google Scholar] [CrossRef]
- Cui, J.; Yu, C.; Qiao, N.; Xu, X.; Tian, Y.; Ouyang, H. Plant Preference for NH4+ versus NO3− at Different Growth Stages in an Alpine Agroecosystem. Field Crops Res. 2017, 201, 192–199. [Google Scholar] [CrossRef]
- Duan, Y.H.; Zhang, Y.L.; Ye, L.T.; Fan, X.R.; Xu, G.H.; Shen, Q.R. Responses of Rice Cultivars with Different Nitrogen Use Efficiency to Partial Nitrate Nutrition. Ann. Bot. 2007, 99, 1153–1160. [Google Scholar] [CrossRef]
- Hajari, E.; Snyman, S.J.; Watt, M.P. Nitrogen Use Efficiency of Sugarcane (Saccharum Spp.) Varieties under in Vitro Conditions with Varied N Supply. Plant Cell Tiss Organ Cult. 2015, 122, 21–29. [Google Scholar] [CrossRef]
- Patterson, K.; Cakmak, T.; Cooper, A.; Lager, I.; Rasmusson, A.G.; Escobar, M.A. Distinct Signalling Pathways and Transcriptome Response Signatures Differentiate Ammonium- and Nitrate-Supplied Plants: Transcriptome Signatures of Ammonium and Nitrate Responses. Plant Cell Environ. 2010, 33, 1486–1501. [Google Scholar] [CrossRef]
- Ding, L.; Lu, Z.; Gao, L.; Guo, S.; Shen, Q. Is Nitrogen a Key Determinant of Water Transport and Photosynthesis in Higher Plants Upon Drought Stress? Front. Plant Sci. 2018, 9, 1143. [Google Scholar] [CrossRef]
- Jiang, W.Z. Mn Use Efficiency in Different Wheat Cultivars. Environ. Exp. Bot. 2006, 57, 41–50. [Google Scholar] [CrossRef]
- Carrasco-Gil, S.; Estebaranz-Yubero, M.; Medel-Cuesta, D.; Millán, R.; Hernández, L.E. Influence of Nitrate Fertilization on Hg Uptake and Oxidative Stress Parameters in Alfalfa Plants Cultivated in a Hg-Polluted Soil. Environ. Exp. Bot. 2012, 75, 16–24. [Google Scholar] [CrossRef]
- Huo, K.; Shangguan, X.; Xia, Y.; Shen, Z.; Chen, C. Excess Copper Inhibits the Growth of Rice Seedlings by Decreasing Uptake of Nitrate. Ecotoxicol. Environ. Saf. 2020, 190, 110105. [Google Scholar] [CrossRef]
- Huang, J.; Wu, X.; Tian, F.; Chen, Q.; Luo, P.; Zhang, F.; Wan, X.; Zhong, Y.; Liu, Q.; Lin, T. Changes in Proteome and Protein Phosphorylation Reveal the Protective Roles of Exogenous Nitrogen in Alleviating Cadmium Toxicity in Poplar Plants. Int. J. Mol. Sci. 2019, 21, 278. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.Q.; Guo, S.W.; Shinmachi, F.; Sunairi, M.; Noguchi, A.; Hasegawa, I.; Shen, R.F. Aluminium Tolerance in Rice Is Antagonistic with Nitrate Preference and Synergistic with Ammonium Preference. Ann. Bot. 2013, 111, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.Q.; Cao, X.C.; Zhu, L.F.; Hu, W.J.; Hu, A.Y.; Bai, Z.G.; Zhong, C.; Sun, L.M.; Liang, Q.D.; Huang, J.; et al. Ammonium Mitigates Cd Toxicity in Rice (Oryza Sativa) via Putrescine-Dependent Alterations of Cell Wall Composition. Plant Physiol. Biochem. 2018, 132, 189–201. [Google Scholar] [CrossRef]
- Zhao, X.Q.; Shen, R.F. Aluminum–Nitrogen Interactions in the Soil–Plant System. Front. Plant Sci. 2018, 9, 807. [Google Scholar] [CrossRef]
- Yu, L.; Wang, X.; Li, X.; Wang, Y.; Kang, H.; Chen, G.; Fan, X.; Sha, L.; Zhou, Y.; Zeng, J. Protective Effect of Different Forms of Nitrogen Application on Cadmium-Induced Toxicity in Wheat Seedlings. Environ. Sci. Pollut. Res. 2019, 26, 13085–13094. [Google Scholar] [CrossRef]
- Afef, N.-H.; Chiraz, C.-H.; Houda, M.D.; Habib, G.; Houda, G. Substitution of NO3− by NH4 + Increases Ammonium-Assimilating Enzyme Activities and Reduces the Deleterious Effects of Cadmium on the Growth of Tomatoes. Fresenius Environ. Bull. 2012, 21, 665–671. [Google Scholar]
- De la Peña, M.; Marín-Peña, A.J.; Urmeneta, L.; Coleto, I.; Castillo-González, J.; van Liempd, S.M.; Falcón-Pérez, J.M.; Álvarez-Fernández, A.; González-Moro, M.B.; Marino, D. Ammonium Nutrition Interacts with Iron Homeostasis in Brachypodium Distachyon. J. Exp. Bot. 2022, 73, 263–274. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, H.; Zhang, S.; Wang, J.; Wei, C. NH4+-N Alleviates Iron Deficiency in Rice Seedlings under Calcareous Conditions. Sci. Rep. 2019, 9, 12712. [Google Scholar] [CrossRef] [PubMed]
- de Souza, J.C., Jr.; Nogueirol, R.C.; Monteiro, F.A. Nitrate and Ammonium Proportion Plays a Key Role in Copper Phytoextraction, Improving the Antioxidant Defense in Tanzania Guinea Grass. Ecotoxicol. Environ. Saf. 2019, 171, 823–832. [Google Scholar] [CrossRef] [PubMed]
- Broadley, M.; Brown, P.; Cakmak, I.; Rengel, Z.; Zhao, F. Function of Nutrients: Micronutrients. In Marschner’s Mineral Nutrition of Higher Plants; Elsevier: Amsterdam, The Netherlands, 2012; pp. 191–248. ISBN 978-0-12-384905-2. [Google Scholar]
- Schmidt, S.B.; Husted, S. The Biochemical Properties of Manganese in Plants. Plants 2019, 8, 381. [Google Scholar] [CrossRef]
- Schmidt, S.B.; Jensen, P.E.; Husted, S. Manganese Deficiency in Plants: The Impact on Photosystem II. Trends Plant Sci. 2016, 21, 622–632. [Google Scholar] [CrossRef] [PubMed]
- Socha, A.L.; Guerinot, M.L. Mn-Euvering Manganese: The Role of Transporter Gene Family Members in Manganese Uptake and Mobilization in Plants. Front. Plant Sci. 2014, 5, 106. [Google Scholar] [CrossRef]
- Pittman, J.K. Managing the Manganese: Molecular Mechanisms of Manganese Transport and Homeostasis. New Phytol. 2005, 167, 733–742. [Google Scholar] [CrossRef] [PubMed]
- Foy, C.D.; Scott, B.J.; Fisher, J.A. Genetic Differences in Plant Tolerance to Manganese Toxicity. In Manganese in Soils and Plants; Developments in Plant and Soil Sciences; ©Kluwer Academic Publishers: Norwell, MA, USA, 1988; Volume 33, pp. 293–307. ISBN 978-94-010-7768-2. [Google Scholar]
- Li, J.; Jia, Y.; Dong, R.; Huang, R.; Liu, P.; Li, X.; Wang, Z.; Liu, G.; Chen, Z. Advances in the Mechanisms of Plant Tolerance to Manganese Toxicity. Int. J. Mol. Sci. 2019, 20, 5096. [Google Scholar] [CrossRef]
- Wissemeier, A.H.; Horst, W.J. Effect of Light Intensity on Manganese Toxicity Symptoms and Callose Formation in Cowpea (Vigna Unguiculata (L.) Walp.). Plant Soil 1992, 143, 299–309. [Google Scholar] [CrossRef]
- Alejandro, S.; Höller, S.; Meier, B.; Peiter, E. Manganese in Plants: From Acquisition to Subcellular Allocation. Front. Plant Sci. 2020, 11, 300. [Google Scholar] [CrossRef]
- Quartin, V.M.L.; Antunes, M.L.; Muralha, M.C.; Sousa, M.M.; Nunes, M.A. Mineral Imbalance Due to Manganese Excess in Triticales. J. Plant Nutrit. 2001, 24, 175–189. [Google Scholar] [CrossRef]
- El Khalil, H.; El Hamiani, O.; Bitton, G.; Ouazzani, N.; Boularbah, A. Heavy Metal Contamination from Mining Sites in South Morocco: Monitoring Metal Content and Toxicity of Soil Runoff and Groundwater. Environ. Monit. Assess. 2008, 136, 147–160. [Google Scholar] [CrossRef]
- Li, P.; Qian, H.; Howard, K.W.F.; Wu, J.; Lyu, X. Anthropogenic Pollution and Variability of Manganese in Alluvial Sediments of the Yellow River, Ningxia, Northwest China. Environ. Monit. Assess. 2014, 186, 1385–1398. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Kuang, Y.; Lan, S.; Cao, W.; Yan, Z.; Chen, L.; Chen, Q.; Feng, Q.; Zhou, H. Pollution Distribution of Potentially Toxic Elements in a Karstic River Affected by Manganese Mining in Changyang, Western Hubei, Central China. Int. J. Environ. Res. Public Helth 2021, 18, 1870. [Google Scholar] [CrossRef] [PubMed]
- Trueman, B.F.; Gregory, B.S.; McCormick, N.E.; Gao, Y.; Gora, S.; Anaviapik-Soucie, T.; L’Hérault, V.; Gagnon, G.A. Manganese Increases Lead Release to Drinking Water. Environ. Sci. Technol. 2019, 53, 4803–4812. [Google Scholar] [CrossRef]
- Pinsino, A.; Matranga, V.; Roccheri, M.C. Manganese: A New Emerging Contaminant in the Environment. In Environmental Contamination; Srivastava, J., Ed.; IntechOpen: London, UK, 2012; pp. 17–37. ISBN 978-953-51-0120-8. [Google Scholar]
- Queiroz, H.M.; Ying, S.C.; Abernathy, M.; Barcellos, D.; Gabriel, F.A.; Otero, X.L.; Nóbrega, G.N.; Bernardino, A.F.; Ferreira, T.O. Manganese: The Overlooked Contaminant in the World Largest Mine Tailings Dam Collapse. Environ. Int. 2021, 146, 106284. [Google Scholar] [CrossRef] [PubMed]
- Acosta, K.; Appenroth, K.J.; Borisjuk, L.; Edelman, M.; Heinig, U.; Jansen, M.A.K.; Oyama, T.; Pasaribu, B.; Schubert, I.; Sorrels, S.; et al. Return of the Lemnaceae: Duckweed as a Model Plant System in the Genomics and Post-Genomics Era. Plant Cell 2021, 33, 3207–3234. [Google Scholar] [CrossRef]
- Zhou, Y.; Borisjuk, N. Small Aquatic Duckweed Plants with Big Potential for the Production of Valuable Biomass and Wastewater Remediation. Int. J. Environ. Sci. Nat. Resour. 2019, 16, 555942. [Google Scholar] [CrossRef]
- Ziegler, P.; Sree, K.S.; Appenroth, K.-J. Duckweeds for Water Remediation and Toxicity Testing. Toxicol. Environ. Chem. 2016, 98, 1127–1154. [Google Scholar] [CrossRef]
- Zhou, Y.; Kishchenko, O.; Stepanenko, A.; Chen, G.; Wang, W.; Zhou, J.; Pan, C.; Borisjuk, N. The Dynamics of NO3− and NH4+ Uptake in Duckweed Are Coordinated with the Expression of Major Nitrogen Assimilation Genes. Plants 2022, 11, 11. [Google Scholar] [CrossRef]
- Zhou, Y.; Bai, T.; Kishchenko, O. Potential of Lemnoideae Species for Phytoremediation of Fresh Water with Elevated Manganese Concentration. Innov. Biosyst. Bioeng. 2019, 3, 232–238. [Google Scholar] [CrossRef][Green Version]
- Michael, T.P.; Bryant, D.; Gutierrez, R.; Borisjuk, N.; Chu, P.; Zhang, H.; Xia, J.; Zhou, J.; Peng, H.; El Baidouri, M.; et al. Comprehensive Definition of Genome Features in Spirodela Polyrhiza by High-Depth Physical Mapping and Short-Read DNA Sequencing Strategies. Plant J. 2017, 89, 617–635. [Google Scholar] [CrossRef] [PubMed]
- Omasits, U.; Ahrens, C.H.; Müller, S.; Wollscheid, B. Protter: Interactive Protein Feature Visualization and Integration with Experimental Proteomic Data. Bioinformatics 2014, 30, 884–886. [Google Scholar] [CrossRef] [PubMed]
- Tippery, N.P.; Les, D.H. Tiny Plants with Enormous Potential: Phylogeny and Evolution of Duckweeds. In The Duckweed Genomes; Cao, X.H., Fourounjian, P., Wang, W., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 19–38. ISBN 978-3-030-11044-4. [Google Scholar]
- Lemoine, F.; Correia, D.; Lefort, V.; Doppelt-Azeroual, O.; Mareuil, F.; Cohen-Boulakia, S.; Gascuel, O. NGPhylogeny.Fr: New Generation Phylogenetic Services for Non-Specialists. Nucleic Acids Res. 2019, 47, W260–W265. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree of Life (ITOL) v5: An Online Tool for Phylogenetic Tree Display and Annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- Marini, A.M.; Soussi-Boudekou, S.; Vissers, S.; Andre, B. A Family of Ammonium Transporters in Saccharomyces Cerevisiae. Mol. Cell Biol. 1997, 17, 4282–4293. [Google Scholar] [CrossRef]
- Higo, K.; Ugawa, Y.; Iwamoto, M.; Korenaga, T. Plant Cis-Acting Regulatory DNA Elements (PLACE) Database: 1999. Nucleic Acids Res. 1999, 27, 297–300. [Google Scholar] [CrossRef]
- Lehmann, M. Anything Else but GAGA: A Nonhistone Protein Complex Reshapes Chromatin Structure. Trends Genet. 2004, 20, 15–22. [Google Scholar] [CrossRef]
- Hecker, A.; Brand, L.H.; Peter, S.; Simoncello, N.; Kilian, J.; Harter, K.; Gaudin, V.; Wanke, D. The Arabidopsis GAGA-Binding Factor BASIC PENTACYSTEINE6 Recruits the POLYCOMB-REPRESSIVE COMPLEX1 Component LIKE HETEROCHROMATIN PROTEIN1 to GAGA DNA Motifs. Plant Physiol. 2015, 168, 1013–1024. [Google Scholar] [CrossRef]
- Tsai, S.-Y.; Chang, Y.-L.; Swamy, K.B.S.; Chiang, R.-L.; Huang, D.-H. GAGA Factor, a Positive Regulator of Global Gene Expression, Modulates Transcriptional Pausing and Organization of Upstream Nucleosomes. Epigenet. Chromatin 2016, 9, 32. [Google Scholar] [CrossRef]
- Hernandez, N. TBP, a Universal Eukaryotic Transcription Factor? Genes Dev. 1993, 7, 1291–1308. [Google Scholar] [CrossRef]
- Komarnytsky, S.; Borisjuk, N. Functional Analysis of Promoter Elements in Plants. Genet. Eng. 2003, 25, 113–141. [Google Scholar] [CrossRef]
- Leyva-González, M.A.; Ibarra-Laclette, E.; Cruz-Ramírez, A.; Herrera-Estrella, L. Functional and Transcriptome Analysis Reveals an Acclimatization Strategy for Abiotic Stress Tolerance Mediated by Arabidopsis NF-YA Family Members. PLoS ONE 2012, 7, e48138. [Google Scholar] [CrossRef] [PubMed]
- Petroni, K.; Kumimoto, R.W.; Gnesutta, N.; Calvenzani, V.; Fornari, M.; Tonelli, C.; Holt, B.F.; Mantovani, R. The Promiscuous Life of Plant NUCLEAR FACTOR Y Transcription Factors. Plant Cell 2013, 24, 4777–4792. [Google Scholar] [CrossRef] [PubMed]
- Hao, D.-L.; Zhou, J.-Y.; Yang, S.-Y.; Qi, W.; Yang, K.-J.; Su, Y.-H. Function and Regulation of Ammonium Transporters in Plants. Int. J. Mol. Sci. 2020, 21, 3557. [Google Scholar] [CrossRef]
- Zhao, X.; Yang, J.; Li, X.; Li, G.; Sun, Z.; Chen, Y.; Chen, Y.; Xia, M.; Li, Y.; Yao, L.; et al. Identification and Expression Analysis of GARP Superfamily Genes in Response to Nitrogen and Phosphorus Stress in Spirodela Polyrhiza. BMC Plant Biol. 2022, 22, 308. [Google Scholar] [CrossRef]
- Wang, X.; Wang, H.-F.; Chen, Y.; Sun, M.-M.; Wang, Y.; Chen, Y.-F. The Transcription Factor NIGT1.2 Modulates Both Phosphate Uptake and Nitrate Influx during Phosphate Starvation in Arabidopsis and Maize. Plant Cell 2020, 32, 3519–3534. [Google Scholar] [CrossRef]
- Yanagisawa, S. Characterization of a Nitrate-Inducible Transcriptional Repressor NIGT1 Provides New Insights into DNA Recognition by the GARP Family Proteins. Plant Signal. Behav. 2013, 8, e24447. [Google Scholar] [CrossRef]
- Li, Q.; Zhou, L.; Li, Y.; Zhang, D.; Gao, Y. Plant NIGT1/HRS1/HHO Transcription Factors: Key Regulators with Multiple Roles in Plant Growth, Development, and Stress Responses. Int. J. Mol. Sci. 2021, 22, 8685. [Google Scholar] [CrossRef]
- Wang, W.; Haberer, G.; Gundlach, H.; Gläßer, C.; Nussbaumer, T.; Luo, M.C.; Lomsadze, A.; Borodovsky, M.; Kerstetter, R.A.; Shanklin, J.; et al. The Spirodela Polyrrhiza Genome Reveals Insights into Its Neotenous Reduction Fast Growth and Aquatic Lifestyle. Nat. Commun. 2014, 5, 3311. [Google Scholar] [CrossRef]
- Hoang, P.T.N.; Fiebig, A.; Novák, P.; Macas, J.; Cao, H.X.; Stepanenko, A.; Chen, G.; Borisjuk, N.; Scholz, U.; Schubert, I. Chromosome-Scale Genome Assembly for the Duckweed Spirodela Intermedia, Integrating Cytogenetic Maps, PacBio and Oxford Nanopore Libraries. Sci. Rep. 2020, 10, 19230. [Google Scholar] [CrossRef]
- Chong, Y.X.; Hu, H.Y.; Qian, Y. Growth feature of biomass of Lemna aequinoctialis and Spirodela polyrrhiza in medium with nutrient character of wastewater. Huan Jing Ke Xue 2004, 25, 59–64. [Google Scholar] [PubMed]
- Feng, H.; Fan, X.; Miller, A.J.; Xu, G. Plant Nitrogen Uptake and Assimilation: Regulation of Cellular PH Homeostasis. J. Exp. Bot. 2020, 71, 4380–4392. [Google Scholar] [CrossRef] [PubMed]
- Hachiya, T.; Sakakibara, H. Interactions between Nitrate and Ammonium in Their Uptake, Allocation, Assimilation, and Signaling in Plants. J. Exp. Bot. 2016, 68, 2501–2512. [Google Scholar] [CrossRef] [PubMed]
- Hu, A.Y.; Zheng, M.M.; Sun, L.M.; Zhao, X.Q.; Shen, R.F. Ammonium Alleviates Manganese Toxicity and Accumulation in Rice by Down-Regulating the Transporter Gene OsNramp5 Through Rhizosphere Acidification. Front. Plant Sci. 2019, 10, 1194. [Google Scholar] [CrossRef] [PubMed]
- Langheinrich, U.; Tischner, R.; Godbold, D.L. Influence of a High Mn Supply on Norway Spruce (Picea abies (L.) Karst.) Seedlings in Relation to the Nitrogen Source. Tree Physiol. 1992, 10, 259–271. [Google Scholar] [CrossRef]
- Gerbeau, P.; Amodeo, G.; Henzler, T.; Santoni, V.; Ripoche, P.; Maurel, C. The Water Permeability of Arabidopsis Plasma Membrane Is Regulated by Divalent Cations and PH. Plant J. 2002, 30, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Franz, M.C.; Pujol-Giménez, J.; Montalbetti, N.; Fernandez-Tenorio, M.; DeGrado, T.R.; Niggli, E.; Romero, M.F.; Hediger, M.A. Reassessment of the Transport Mechanism of the Human Zinc Transporter SLC39A2. Biochemistry 2018, 57, 3976–3986. [Google Scholar] [CrossRef]
- Chauvigné, F.; Zapater, C.; Stavang, J.A.; Taranger, G.L.; Cerdà, J.; Finn, R.N. The PH Sensitivity of Aqp0 Channels in Tetraploid and Diploid Teleosts. FASEB J. 2015, 29, 2172–2184. [Google Scholar] [CrossRef]
- Eide, D.J. Transcription Factors and Transporters in Zinc Homeostasis: Lessons Learned from Fungi. Crit. Rev. Biochem. Mol. Biol. 2020, 55, 88–110. [Google Scholar] [CrossRef]
- Wang, J.; Yu, X.; Ding, Z.J.; Zhang, X.; Luo, Y.; Xu, X.; Xie, Y.; Li, X.; Yuan, T.; Zheng, S.J.; et al. Structural Basis of ALMT1-Mediated Aluminum Resistance in Arabidopsis. Cell Res. 2022, 32, 89–98. [Google Scholar] [CrossRef]
- Tournaire-Roux, C.; Sutka, M.; Javot, H.; Gout, E.; Gerbeau, P.; Luu, D.-T.; Bligny, R.; Maurel, C. Cytosolic PH Regulates Root Water Transport during Anoxic Stress through Gating of Aquaporins. Nature 2003, 425, 393–397. [Google Scholar] [CrossRef] [PubMed]
- Scochera, F.; Zerbetto De Palma, G.; Canessa Fortuna, A.; Chevriau, J.; Toriano, R.; Soto, G.; Zeida, A.; Alleva, K. PIP Aquaporin PH-sensing Is Regulated by the Length and Charge of the C-terminal Region. FEBS J. 2022, 289, 246–261. [Google Scholar] [CrossRef] [PubMed]
- Soto, G.; Alleva, K.; Amodeo, G.; Muschietti, J.; Ayub, N.D. New Insight into the Evolution of Aquaporins from Flowering Plants and Vertebrates: Orthologous Identification and Functional Transfer Is Possible. Gene 2012, 503, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Maurel, C.; Boursiac, Y.; Luu, D.-T.; Santoni, V.; Shahzad, Z.; Verdoucq, L. Aquaporins in Plants. Physiol. Rev. 2015, 95, 1321–1358. [Google Scholar] [CrossRef]
- Kourghi, M.; Nourmohammadi, S.; Pei, J.; Qiu, J.; McGaughey, S.; Tyerman, S.; Byrt, C.; Yool, A. Divalent Cations Regulate the Ion Conductance Properties of Diverse Classes of Aquaporins. Int. J. Mol. Sci. 2017, 18, 2323. [Google Scholar] [CrossRef]
- Verdoucq, L.; Grondin, A.; Maurel, C. Structure–function Analysis of Plant Aquaporin AtPIP2;1 Gating by Divalent Cations and Protons. Biochem. J. 2008, 415, 409–416. [Google Scholar] [CrossRef]
- Schwacke, R.; Schneider, A.; van der Graaff, E.; Fischer, K.; Catoni, E.; Desimone, M.; Frommer, W.B.; Flügge, U.-I.; Kunze, R. ARAMEMNON, a Novel Database for Arabidopsis Integral Membrane Proteins. Plant Physiol. 2003, 131, 16–26. [Google Scholar] [CrossRef]
- Neuhäuser, B.; Dynowski, M.; Mayer, M.; Ludewig, U. Regulation of NH4 + Transport by Essential Cross Talk between AMT Monomers through the Carboxyl Tails. Plant Physiol. 2007, 143, 1651–1659. [Google Scholar] [CrossRef]
- Ludewig, U.; Neuhäuser, B.; Dynowski, M. Molecular Mechanisms of Ammonium Transport and Accumulation in Plants. FEBS Lett. 2007, 581, 2301–2308. [Google Scholar] [CrossRef]
- Sonoda, Y.; Ikeda, A.; Saiki, S.; von Wirén, N.; Yamaya, T.; Yamaguchi, J. Distinct Expression and Function of Three Ammonium Transporter Genes (OsAMT1;1–1;3) in Rice. Plant Cell Physiol. 2003, 44, 726–734. [Google Scholar] [CrossRef]
- Li, B.; Merrick, M.; Li, S.; Li, H.; Zhu, S.; Shi, W.; Su, Y. Molecular Basis and Regulation of Ammonium Transporter in Rice. Rice Sci. 2009, 16, 314–322. [Google Scholar] [CrossRef]
- Xu, S.; Stapley, J.; Gablenz, S.; Boyer, J.; Appenroth, K.J.; Sree, K.S.; Gershenzon, J.; Widmer, A.; Huber, M. Low Genetic Variation Is Associated with Low Mutation Rate in the Giant Duckweed. Nat. Commun. 2019, 10, 1243. [Google Scholar] [CrossRef] [PubMed]
- Bellinger, M.R.; Paudel, R.; Starnes, S.; Kambic, L.; Kantar, M.B.; Wolfgruber, T.; Lamour, K.; Geib, S.; Sim, S.; Miyasaka, S.C.; et al. Taro Genome Assembly and Linkage Map Reveal QTLs for Resistance to Taro Leaf Blight. G3 Genes Genomes Genet. 2020, 10, 2763–2775. [Google Scholar] [CrossRef] [PubMed]
- Shi, T.; Huneau, C.; Zhang, Y.; Li, Y.; Chen, J.; Salse, J.; Wang, Q. The Slow-Evolving Acorus Tatarinowii Genome Sheds Light on Ancestral Monocot Evolution. Nat. Plants 2022, 8, 764–777. [Google Scholar] [CrossRef]
- Wu, Z.; Gao, X.; Zhang, N.; Feng, X.; Huang, Y.; Zeng, Q.; Wu, J.; Zhang, J.; Qi, Y. Genome-Wide Identification and Transcriptional Analysis of Ammonium Transporters in Saccharum. Genomics 2021, 113, 1671–1680. [Google Scholar] [CrossRef]
- Straub, T.; Ludewig, U.; Neuhäuser, B. The Kinase CIPK23 Inhibits Ammonium Transport in Arabidopsis Thaliana. Plant Cell 2017, 29, 409–422. [Google Scholar] [CrossRef]
- Wu, X.; Liu, T.; Zhang, Y.; Duan, F.; Neuhäuser, B.; Ludewig, U.; Schulze, W.X.; Yuan, L. Ammonium and Nitrate Regulate NH4+ Uptake Activity of Arabidopsis Ammonium Transporter AtAMT1;3 via Phosphorylation at Multiple C-Terminal Sites. J. Exp. Bot. 2019, 70, 4919–4930. [Google Scholar] [CrossRef]
- Ganz, P.; Porras-Murillo, R.; Ijato, T.; Menz, J.; Straub, T.; Stührwohldt, N.; Moradtalab, N.; Ludewig, U.; Neuhäuser, B. Abscisic Acid Influences Ammonium Transport via Regulation of Kinase CIPK23 and Ammonium Transporters. Plant Physiol. 2022, 190, 1275–1288. [Google Scholar] [CrossRef]
- Kaiser, B.N. Functional Analysis of an Arabidopsis T-DNA “Knockout” of the High-Affinity NH4+ Transporter AtAMT1;1. Plant Physiol. 2002, 130, 1263–1275. [Google Scholar] [CrossRef]
- Loqué, D.; Lalonde, S.; Looger, L.L.; von Wirén, N.; Frommer, W.B. A Cytosolic Trans-Activation Domain Essential for Ammonium Uptake. Nature 2007, 446, 195–198. [Google Scholar] [CrossRef]
- Ludewig, U.; Wilken, S.; Wu, B.; Jost, W.; Obrdlik, P.; El Bakkoury, M.; Marini, A.-M.; André, B.; Hamacher, T.; Boles, E.; et al. Homo- and Hetero-Oligomerization of Ammonium Transporter-1 NH 4 + Uniporters. J. Biol. Chem. 2003, 278, 45603–45610. [Google Scholar] [CrossRef] [PubMed]
- Nühse, T.S.; Stensballe, A.; Jensen, O.N.; Peck, S.C. Phosphoproteomics of the Arabidopsis Plasma Membrane and a New Phosphorylation Site Database. Plant Cell 2004, 16, 2394–2405. [Google Scholar] [CrossRef] [PubMed]
- Gazzarrini, S.; Lejay, L.; Gojon, A.; Ninnemann, O.; Frommer, W.B.; von Wirén, N. Three Functional Transporters for Constitutive, Diurnally Regulated, and Starvation-Induced Uptake of Ammonium into Arabidopsis Roots. Plant Cell 1999, 11, 937–947. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Silim, S.N.; Okamoto, M.; Siddiqi, M.Y.; Glass, A.D.M. Differential Expression of Three Members of the AMT1 Gene Family Encoding Putative High-Affinity NH4+ Transporters in Roots of Oryza Sativa Subspecies Indica: Regulation of AMT1 Genes in Rice. Plant Cell Environ. 2003, 26, 907–914. [Google Scholar] [CrossRef] [PubMed]
- Gu, R.; Duan, F.; An, X.; Zhang, F.; von Wirén, N.; Yuan, L. Characterization of AMT-Mediated High-Affinity Ammonium Uptake in Roots of Maize (Zea mays L.). Plant Cell Physiol. 2013, 54, 1515–1524. [Google Scholar] [CrossRef]
- Wu, X.; Yang, H.; Qu, C.; Xu, Z.; Li, W.; Hao, B.; Yang, C.; Sun, G.; Liu, G. Sequence and Expression Analysis of the AMT Gene Family in Poplar. Front. Plant Sci. 2015, 6, 337. [Google Scholar] [CrossRef]
- Li, H.; Yang, Q.; Liu, W.; Lin, J.; Chang, Y. The AMT1 Family Genes from Malus Robusta Display Differential Transcription Features and Ammonium Transport Abilities. Mol. Biol. Rep. 2017, 44, 379–390. [Google Scholar] [CrossRef]
- Liu, L.-H.; Fan, T.-F.; Shi, D.-X.; Li, C.-J.; He, M.-J.; Chen, Y.-Y.; Zhang, L.; Yang, C.; Cheng, X.-Y.; Chen, X.; et al. Coding-Sequence Identification and Transcriptional Profiling of Nine AMTs and Four NRTs From Tobacco Revealed Their Differential Regulation by Developmental Stages, Nitrogen Nutrition, and Photoperiod. Front. Plant Sci. 2018, 9, 210. [Google Scholar] [CrossRef]
- Masclaux-Daubresse, C.; Reisdorf-Cren, M.; Orsel, M. Leaf Nitrogen Remobilization for Plant Development and Grain Filling. Plant Biol. 2008, 10, 23–36. [Google Scholar] [CrossRef]
- Van der Graaff, E.; Schwacke, R.; Schneider, A.; Desimone, M.; Flügge, U.-I.; Kunze, R. Transcription Analysis of Arabidopsis Membrane Transporters and Hormone Pathways during Developmental and Induced Leaf Senescence. Plant Physiol. 2006, 141, 776–792. [Google Scholar] [CrossRef]
- Zhao, H.; Wu, D.; Kong, F.; Lin, K.; Zhang, H.; Li, G. The Arabidopsis thaliana Nuclear Factor Y Transcription Factors. Front. Plant Sci. 2017, 7, 2045. [Google Scholar] [CrossRef] [PubMed]
- Myers, Z.A.; Holt, B.F. NUCLEAR FACTOR-Y: Still complex after all these years? Curr. Opin. Plant Biol. 2018, 45, 96–102. [Google Scholar] [CrossRef]
- Swain, S.; Myers, Z.A.; Siriwardana, C.L.; Holt, B.F. The multifaceted roles of NUCLEAR FACTOR-Y in Arabidopsis thaliana development and stress responses. Biochim. Biophys. Acta Gene Regul. Mech. 2017, 1860, 636–644. [Google Scholar] [CrossRef]
- Du, W.; Yang, J.; Li, Q.; He, C.; Pang, Y. Identification and Characterization of Abiotic Stress-Responsive NF-YB Family Genes in Medicago. Int. J. Mol. Sci. 2022, 23, 6906. [Google Scholar] [CrossRef] [PubMed]
- Schenk, R.U.; Hildebrandt, A.C. Medium and Techniques for Induction and Growth of Monocotyledonous and Dicotyledonous Plant Cell Cultures. Can. J. Bot. 1972, 50, 199–204. [Google Scholar] [CrossRef]
- Zhou, Y.; Chen, G.; Peterson, A.; Zha, X.; Cheng, J.; Li, S.; Cui, D.; Zhu, H.; Kishchenko, O.; Borisjuk, N. Biodiversity of Duckweeds in Eastern China and Their Potential for Bioremediation of Municipal and Industrial Wastewater. J. Geosci. Environ. Protect. 2018, 06, 108–116. [Google Scholar] [CrossRef][Green Version]
- Cedergreen, N.; Madsen, T.V. Nitrogen Uptake by the Floating Macrophyte Lemna Minor. New Phytol. 2002, 155, 285–292. [Google Scholar] [CrossRef]
- American Water Works Association (AWWA). Standard Methods for the Examination of Water and Wastewater, 21st ed.; AWWA: Washington, DC, USA, 2005. [Google Scholar]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Criscuolo, A.; Gribaldo, S. BMGE (Block Mapping and Gathering with Entropy): A New Software for Selection of Phylogenetic Informative Regions from Multiple Sequence Alignments. BMC Evol. Biol. 2010, 10, 210. [Google Scholar] [CrossRef]
- Ludewig, U.; von Wirén, N.; Frommer, W.B. Uniport of NH 4 + by the Root Hair Plasma Membrane Ammonium Transporter LeAMT1;1. J. Biol. Chem. 2002, 277, 13548–13555. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A New Mathematical Model for Relative Quantification in Real-Time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate Normalization of Real-Time Quantitative RT-PCR Data by Geometric Averaging of Multiple Internal Control Genes. Genome Biol. 2002, 3, RESEARCH0034. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kishchenko, O.; Stepanenko, A.; Straub, T.; Zhou, Y.; Neuhäuser, B.; Borisjuk, N. Ammonium Uptake, Mediated by Ammonium Transporters, Mitigates Manganese Toxicity in Duckweed, Spirodela polyrhiza. Plants 2023, 12, 208. https://doi.org/10.3390/plants12010208
Kishchenko O, Stepanenko A, Straub T, Zhou Y, Neuhäuser B, Borisjuk N. Ammonium Uptake, Mediated by Ammonium Transporters, Mitigates Manganese Toxicity in Duckweed, Spirodela polyrhiza. Plants. 2023; 12(1):208. https://doi.org/10.3390/plants12010208
Chicago/Turabian StyleKishchenko, Olena, Anton Stepanenko, Tatsiana Straub, Yuzhen Zhou, Benjamin Neuhäuser, and Nikolai Borisjuk. 2023. "Ammonium Uptake, Mediated by Ammonium Transporters, Mitigates Manganese Toxicity in Duckweed, Spirodela polyrhiza" Plants 12, no. 1: 208. https://doi.org/10.3390/plants12010208
APA StyleKishchenko, O., Stepanenko, A., Straub, T., Zhou, Y., Neuhäuser, B., & Borisjuk, N. (2023). Ammonium Uptake, Mediated by Ammonium Transporters, Mitigates Manganese Toxicity in Duckweed, Spirodela polyrhiza. Plants, 12(1), 208. https://doi.org/10.3390/plants12010208