Defining the Role of Attention in Hierarchical Auditory Processing


Abstract
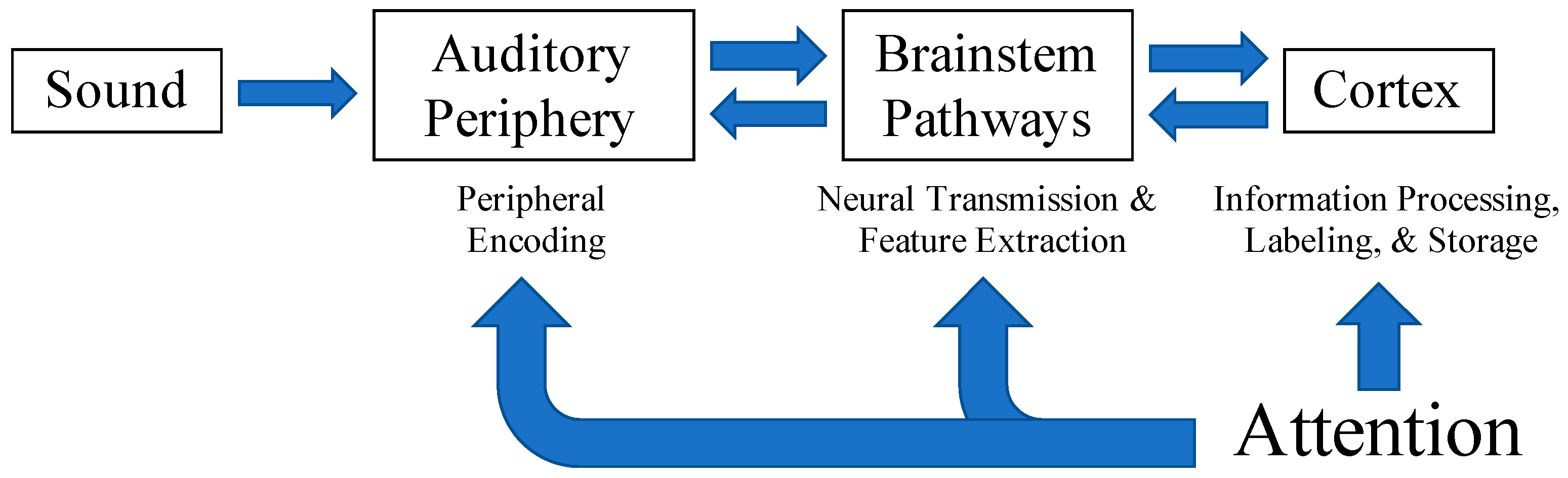
1. Introduction
2. Attentional Theories
2.1. Early vs. Late Filter Theories

{kind=link}
| Theory Type | Attentional Theory | Description | References |
|---|---|---|---|
| Filter | Early filter theory | All or nothing filtering mechanism Filtering occurs prior to perceptual analysis | Broadbent [51,55] |
| Attenuated filter model | Processing of unattended stimuli attenuated relative to attended inputs | Treisman [52,58] Treisman & Geffen [59] | |
| Late filter theory | All inputs processed fully Filtering occurs prior to response selection | Deutsch & Deutsch [53] | |
| Limited capacity | Limited capacity theory | All inputs fully processed when spare capacity available Attentional selection occurs when task demands exceed capacity | Kahneman [54] |
| Framework for Understanding Effortful Listening (FUEL) | Adapts Kahneman’s model to auditory perception providing more direct implications for listening adverse conditions and hearing loss | Pichora-Fuller et al. [60] | |
| Load theory | System comprised of 2 mechanisms: passive perceptual selection & active cognitive control Attentional selection only occurs under high perceptual load; otherwise, all inputs fully processed | Lavie [61] Lavie et al. [62] |
2.2. Limited Capacity Theories
3. Anatomical Studies
4. Electrophysiologic Studies
4.1. Late Auditory Evoked Potentials
4.2. Early Auditory Evoked Potentials
4.3. Simultaneous Recording of Brainstem and Cortical Evoked Responses
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mody, M.; Studdert-Kennedy, M.; Brady, S. Speech perception deficits in poor readers: Auditory processing or phonological coding? J. Exp. Child Psychol. 1997, 64, 199–231. [Google Scholar] [CrossRef] [PubMed]
- Nittrouer, S. From ear to cortex: A perspective on what clinicians need to understand about speech perception and language processing. Lang. Speech Hear. Serv. Sch. 2002, 33, 237–252. [Google Scholar] [CrossRef]
- Lewis, D.; Hoover, B.; Choi, S.; Stelmachowicz, P. Relationship between speech perception in noise and phonological awareness skills for children with normal hearing. Ear Hear. 2010, 31, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Helfer, K.S.; Wilber, L.A. Hearing loss, aging, and speech perception in reverberation and noise. J. Speech Lang. Hear. Res. 1990, 33, 149–155. [Google Scholar] [CrossRef]
- Wong, P.C.; Ettlinger, M.; Sheppard, J.P.; Gunasekera, G.M.; Dhar, S. Neuroanatomical characteristics and speech perception in noise in older adults. Ear Hear. 2010, 31, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Gordon-Salant, S.; Fitzgibbons, P.J. Profile of auditory temporal processing in older listeners. J. Speech Lang. Hear. Res. 1999, 42, 300–311. [Google Scholar] [CrossRef]
- Guest, H.; Munro, K.J.; Prendergast, G.; Millman, R.E.; Plack, C.J. Impaired speech perception in noise with a normal audiogram: No evidence for cochlear synaptopathy and no relation to lifetime noise exposure. Hear. Res. 2018, 364, 142–151. [Google Scholar] [CrossRef]
- Weinstein, B.E.; Ventry, I.M. Hearing impairment and social isolation in the elderly. J. Speech Hear. Res. 1982, 25, 593–599. [Google Scholar] [CrossRef]
- Gopinath, B.; Wang, J.; Schneider, J.; Burlutsky, G.; Snowdon, J.; McMahon, C.M.; Leeder, S.R.; Mitchell, P. Depressive symptoms in older adults with hearing impairment: The Blue Mountains study. J. Am. Geriatr. Soc. 2009, 57, 1306–1308. [Google Scholar] [CrossRef] [PubMed]
- Dalton, D.S.; Cruickshanks, K.J.; Klein, B.E.; Klein, R.; Wiley, T.L.; Nondahl, D.M. The impact of hearing loss on quality of life in older adults. Gerontologist 2003, 43, 661–668. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.R.; Ferrucci, L.; Metter, E.J.; An, Y.; Zonderman, A.B.; Resnick, S.M. Hearing loss and cognition in the Baltimore Longitudinal Study of Aging. Neuropsychology 2011, 25, 763–770. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.R.; Yaffe, K.; Xia, J.; Xue, Q.L.; Harris, T.B.; Purchase-Helzner, E.; Satterfield, S.; Ayonayon, H.N.; Ferrucci, L.; Simonsick, E.M.; et al. Hearing loss and cognitive decline in older adults. JAMA Intern. Med. 2013, 173, 293–299. [Google Scholar] [CrossRef]
- Moore, D.R. Listening difficulties in children: Bottom-up and top-down contributions. J. Commun. Disord. 2012, 45, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Werner, L.A. Issues in human auditory development. J. Commun. Disord. 2007, 40, 275–283. [Google Scholar] [CrossRef]
- Moore, D.R. Sources of pathology underlying listening disorders in children. Int. J. Psychophysiol. 2015, 95, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Humes, L.E. Speech understanding in the elderly. J. Am. Acad. Audiol. 1996, 7, 161–167. [Google Scholar] [PubMed]
- Cunningham, J.; Nicol, T.; Zecker, S.; Bradlow, A.; Kraus, N. Neurobiologic responses to speech in noise in children with learning problems: Deficits and strategies for improvement. Clin. Neurophysiol. 2001, 112, 758–767. [Google Scholar] [CrossRef]
- Rocha-Muniz, C.N.; Befi-Lopes, D.M.; Schochat, E. Investigation of auditory processing disorder and language impairment using the speech-evoked auditory brainstem response. Hear. Res. 2012, 294, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Billiet, C.R.; Bellis, T.J. The relationship between brainstem temporal processing and performance on tests of central auditory function in children with reading disorders. J. Speech Lang. Hear. Res. 2011, 54, 228–242. [Google Scholar] [CrossRef]
- Anderson, S.; Skoe, E.; Chandrasekaran, B.; Kraus, N. Neural timing is linked to speech perception in noise. J. Neurosci. 2010, 30, 4922–4926. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Skoe, E.; Banai, K.; Kraus, N. Perception of speech in noise: Neural correlates. J. Cogn. Neurosci. 2010, 23, 2268–2279. [Google Scholar] [CrossRef] [PubMed]
- Parbery-Clark, A.; Marmel, F.; Bair, J.; Kraus, N. What subcortical-cortical relationships tell us about processing speech in noise. Eur. J. Neurosci. 2011, 33, 549–557. [Google Scholar] [CrossRef]
- Billings, C.J.; McMillan, G.P.; Penman, T.M.; Gille, S.M. Predicting perception in noise using cortical auditory evoked potentials. J. Assoc. Res. Otolaryngol. 2013, 14, 891–903. [Google Scholar] [CrossRef]
- Anderson, S.; Parbery-Clark, A.; Yi, H.G.; Kraus, N. A neural basis of speech-in-noise perception in older adults. Ear Hear. 2011, 32, 750–757. [Google Scholar] [CrossRef] [PubMed]
- Wingfield, A. Cognitive factors in auditory performance: Context, speed of processing, and constraints of memory. J. Am. Acad. Audiol. 1996, 7, 175–182. [Google Scholar] [PubMed]
- Dryden, A.; Allen, H.A.; Henshaw, H.; Heinrich, A. The Association Between Cognitive Performance and Speech-in-Noise Perception for Adult Listeners: A Systematic Literature Review and Meta-Analysis. Trends Hear. 2017, 21, 2331216517744675. [Google Scholar] [CrossRef] [PubMed]
- Hadar, B.; Skrzypek, J.E.; Wingfield, A.; Ben-David, B.M. Working Memory Load Affects Processing Time in Spoken Word Recognition: Evidence from Eye-Movements. Front. Neurosci. 2016, 10, 221. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Dhamani, I.; Leung, J.; Carlile, S. Attention, memory, and auditory processing in 10- to 15-year-old children with listening difficulties. J. Speech Lang. Hear. Res. 2014, 57, 2308–2321. [Google Scholar] [CrossRef] [PubMed]
- Price, C.N.; Alain, C.; Bidelman, G.M. Auditory-frontal channeling in alpha and beta bands is altered by age-related hearing loss and relates to speech perception in noise. Neuroscience 2019, 423, 18–28. [Google Scholar] [CrossRef] [PubMed]
- van Knijff, E.C.; Coene, M.; Govaerts, P.J. Speech understanding in noise in elderly adults: The effect of inhibitory control and syntactic complexity. Int. J. Lang. Commun. Disord. 2018, 53, 628–642. [Google Scholar] [CrossRef] [PubMed]
- Roque, L.; Karawani, H.; Gordon-Salant, S.; Anderson, S. Effects of Age, Cognition, and Neural Encoding on the Perception of Temporal Speech Cues. Front. Neurosci. 2019, 13, 749. [Google Scholar] [CrossRef]
- Petersen, S.E.; Posner, M.I. The attention system of the human brain: 20 years after. Annu. Rev. Neurosci. 2012, 35, 73–89. [Google Scholar] [CrossRef]
- Hernandez-Peon, R. Physiological mechanisms in attention. In Frontiers in Physiological Psychology; Russell, R.W., Ed.; Academic: New York, NY, USA, 1966. [Google Scholar]
- Lukas, J.H. Human auditory attention: The olivocochlear bundle may function as a peripheral filter. Psychophysiology 1980, 17, 444–452. [Google Scholar] [CrossRef] [PubMed]
- Lukas, J.H. The role of efferent inhibition in human auditory attention: An examination of the auditory brainstem potentials. Int. J. Neurosci. 1981, 12, 137–145. [Google Scholar] [CrossRef]
- Picton, T.W.; Hillyard, S.A. Human auditory evoked potentials. II. Effects of attention. Electroencephalogr. Clin. Neurophysiol. 1974, 36, 191–199. [Google Scholar] [CrossRef]
- Galbraith, G.C.; Olfman, D.M.; Huffman, T.M. Selective attention affects human brain stem frequency-following response. Neuroreport 2003, 14, 735–738. [Google Scholar] [CrossRef] [PubMed]
- Suga, N. Role of corticofugal feedback in hearing. J. Comp. Physiol. A 2008, 194, 169–183. [Google Scholar] [CrossRef]
- Suga, N.; Ma, X. Multiparametric corticofugal modulation and plasticity in the auditory system. Nat. Rev. Neurosci. 2003, 4, 783–794. [Google Scholar] [CrossRef] [PubMed]
- Schofield, B. Structural organization of the descending auditory pathway. In Oxford Handbook of Auditory Science: The Auditory Brain; Rees, A., Palmer, A.R., Eds.; Oxford University Press: New York, NY, USA, 2010; pp. 43–64. [Google Scholar]
- Perrot, X.; Ryvlin, P.; Isnard, J.; Guenot, M.; Catenoix, H.; Fischer, C.; Mauguiere, F.; Collet, L. Evidence for corticofugal modulation of peripheral auditory activity in humans. Cereb. Cortex 2006, 16, 941–948. [Google Scholar] [CrossRef] [PubMed]
- Terreros, G.; Delano, P.H. Corticofugal modulation of peripheral auditory responses. Front. Syst. Neurosci. 2015, 9, 134. [Google Scholar] [CrossRef]
- Gao, E.; Suga, N. Experience-dependent plasticity in the auditory cortex and the inferior colliculus of bats: Role of the corticofugal system. Proc. Natl. Acad. Sci. USA 2000, 97, 8081–8086. [Google Scholar] [CrossRef] [PubMed]
- Atiani, S.; Elhilali, M.; David, S.V.; Fritz, J.B.; Shamma, S.A. Task difficulty and performance induce diverse adaptive patterns in gain and shape of primary auditory cortical receptive fields. Neuron 2009, 61, 467–480. [Google Scholar] [CrossRef] [PubMed]
- Jaaskelainen, I.P.; Ahveninen, J. Auditory-cortex short-term plasticity induced by selective attention. Neural Plast. 2014, 2014, 216731. [Google Scholar] [CrossRef] [PubMed]
- Gallun, F.J.; Jakien, K.M. The ability to allocate attentional resources to a memory task predicts speech-on-speech masking for older listeners. In Proceedings of the 23rd International Congress on Acoustics, Aachen, Germany, 9–13 September 2019. [Google Scholar]
- Luck, S. An Introduction to the Event-Related Potential Technique; MIT Press: Cambridge, MA, USA, 2005. [Google Scholar]
- Starr, A.; Golob, E.J. Cognitive factors modulating auditory cortical potentials. In Auditory Evoked Potentials: Basic Principles and Clinical Application; Burkard, R.F., Eggermont, J.J., Don, M., Eds.; Lippincott Williams & Wilkins: Baltimore, MD, USA, 2007; pp. 508–524. [Google Scholar]
- Sussman, E.S. Auditory Scene Analysis: An Attention Perspective. J. Speech Lang. Hear. Res. 2017, 60, 2989–3000. [Google Scholar] [CrossRef] [PubMed]
- Marsh, J.E.; Campbell, T.A. Processing Complex Sounds Passing through the Rostral Brainstem: The New Early Filter Model. Front. Neurosci. 2016, 10, 136. [Google Scholar] [CrossRef]
- Broadbent, D.E. Perception and Communication; Pergamon Press: New York, NY, USA, 1958; 338p. [Google Scholar]
- Treisman, A.M. Contextual cues in selective listening. Q. J. Exp. Psychol. 1960, 12, 242–248. [Google Scholar] [CrossRef]
- Deutsch, J.A.; Deutsch, D. Attention: Some theoretical considerations. Psychol. Rev. 1963, 70, 80–90. [Google Scholar] [CrossRef]
- Kahneman, D. Attention and Effort; Prentice-Hall: Englewood Cliffs, NJ, USA, 1973; 246p. [Google Scholar]
- Broadbent, D.E. Decision and stress; Academic P.: London, UK; New York, NY, USA, 1971. [Google Scholar]
- Cherry, E.C. Some experiments on the recognition of speech, with one and with two ears. J. Acoust. Soc. Am. 1953, 25, 975–979. [Google Scholar] [CrossRef]
- Moray, J. Attention in dichotic listening: Affective cues and the influence of instructions. Q. J. Exp. Psychol. 1959, 56–60. [Google Scholar] [CrossRef]
- Treisman, A.M. Strategies and models of selective attention. Psychol. Rev. 1969, 76, 282–299. [Google Scholar] [CrossRef]
- Treisman, A.M.; Geffen, G. Selective attention: Perception or response? Q. J. Exp. Psychol. 1967, 109, 1–17. [Google Scholar] [CrossRef]
- Pichora-Fuller, M.K.; Kramer, S.E.; Eckert, M.A.; Edwards, B.; Hornsby, B.W.; Humes, L.E.; Lemke, U.; Lunner, T.; Matthen, M.; Mackersie, C.L.; et al. Hearing Impairment and Cognitive Energy: The Framework for Understanding Effortful Listening (FUEL). Ear Hear. 2016, 37 (Suppl. 1), 5S–27S. [Google Scholar] [CrossRef]
- Lavie, N. Perceptual load as a necessary condition for selective attention. J. Exp. Psychol. Hum. Percept. Perform. 1995, 21, 451–468. [Google Scholar] [CrossRef] [PubMed]
- Lavie, N.; Hirst, A.; de Fockert, J.W.; Viding, E. Load theory of selective attention and cognitive control. J. Exp. Psychol. Gen. 2004, 133, 339–354. [Google Scholar] [CrossRef] [PubMed]
- Laberge, D.; Tweedy, J.R. Presentation Probability and Choice Time. J. Exp. Psychol. 1964, 68, 477–481. [Google Scholar] [CrossRef] [PubMed]
- Murphy, S.; Spence, C.; Dalton, P. Auditory perceptual load: A review. Hear. Res. 2017, 352, 40–48. [Google Scholar] [CrossRef]
- Beck, D.M.; Lavie, N. Look here but ignore what you see: Effects of distractors at fixation. J. Exp. Psychol. Hum. Percept. Perform. 2005, 31, 592–607. [Google Scholar] [CrossRef]
- Cartwright-Finch, U.; Lavie, N. The role of perceptual load in inattentional blindness. Cognition 2007, 102, 321–340. [Google Scholar] [CrossRef]
- Forster, S.; Lavie, N. High perceptual load makes everybody equal: Eliminating individual differences in distractibility with load. Psychol. Sci. 2007, 18, 377–381. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, T.; Sato, T. Perceptual limits in a simulated “Cocktail party”. Atten. Percept. Psychophys. 2015, 77, 2108–2120. [Google Scholar] [CrossRef]
- Fairnie, J.; Moore, B.C.; Remington, A. Missing a trick: Auditory load modulates conscious awareness in audition. J. Exp. Psychol. Hum. Percept. Perform. 2016, 42, 930–938. [Google Scholar] [CrossRef] [PubMed]
- Murphy, S.; Fraenkel, N.; Dalton, P. Perceptual load does not modulate auditory distractor processing. Cognition 2013, 129, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Alain, C.; Izenberg, A. Effects of attentional load on auditory scene analysis. J. Cogn. Neurosci. 2003, 15, 1063–1073. [Google Scholar] [CrossRef] [PubMed]
- Zion Golumbic, E.M.; Ding, N.; Bickel, S.; Lakatos, P.; Schevon, C.A.; McKhann, G.M.; Goodman, R.R.; Emerson, R.; Mehta, A.D.; Simon, J.Z.; et al. Mechanisms underlying selective neuronal tracking of attended speech at a “cocktail party”. Neuron 2013, 77, 980–991. [Google Scholar] [CrossRef] [PubMed]
- Galbraith, G.C.; Bhuta, S.M.; Choate, A.K.; Kitahara, J.M.; Mullen, T.A. Brainstem frequency-following response to dichotic vowels during attention. NeuroReport 1998, 9, 1889–1893. [Google Scholar] [CrossRef] [PubMed]
- Galbraith, G.C.; Doan, B.Q. Brainstem frequency-following and behavioral responses during selective attention to pure tones and missing fundamental stimuli. Int. J. Psychophysiol. 1995, 19, 203–214. [Google Scholar] [CrossRef]
- Sowell, E.R.; Thompson, P.M.; Welcome, S.E.; Henkenius, A.L.; Toga, A.W.; Peterson, B.S. Cortical abnormalities in children and adolescents with attention-deficit hyperactivity disorder. Lancet 2003, 362, 1699–1707. [Google Scholar] [CrossRef]
- Seidman, L.J.; Valera, E.M.; Makris, N. Structural brain imaging of attention-deficit/hyperactivity disorder. Biol. Psychiatry 2005, 57, 1263–1272. [Google Scholar] [CrossRef] [PubMed]
- Castellanos, F.X.; Lee, P.P.; Sharp, W.; Jeffries, N.O.; Greenstein, D.K.; Clasen, L.S.; Blumenthal, J.D.; James, R.S.; Ebens, C.L.; Walter, J.M.; et al. Developmental trajectories of brain volume abnormalities in children and adolescents with attention-deficit/hyperactivity disorder. JAMA 2002, 288, 1740–1748. [Google Scholar] [CrossRef]
- Heilman, K.M.; Pandya, D.N.; Geschwind, N. Trimodal inattention following parietal lobe ablations. Trans. Am. Neurol. Assoc. 1970, 95, 259–261. [Google Scholar]
- Alexander, G.E.; DeLong, M.R.; Strick, P.L. Parallel organization of functionally segregated circuits linking basal ganglia and cortex. Annu. Rev. Neurosci. 1986, 9, 357–381. [Google Scholar] [CrossRef] [PubMed]
- Filipek, P.A.; Semrud-Clikeman, M.; Steingard, R.J.; Renshaw, P.F.; Kennedy, D.N.; Biederman, J. Volumetric MRI analysis comparing subjects having attention-deficit hyperactivity disorder with normal controls. Neurology 1997, 48, 589–601. [Google Scholar] [CrossRef] [PubMed]
- Shaw, P.; Eckstrand, K.; Sharp, W.; Blumenthal, J.; Lerch, J.P.; Greenstein, D.; Clasen, L.; Evans, A.; Giedd, J.; Rapoport, J.L. Attention-deficit/hyperactivity disorder is characterized by a delay in cortical maturation. Proc. Natl. Acad. Sci. USA 2007, 104, 19649–19654. [Google Scholar] [CrossRef] [PubMed]
- Toga, A.W.; Thompson, P.M.; Sowell, E.R. Mapping brain maturation. Trends Neurosci. 2006, 29, 148–159. [Google Scholar] [CrossRef]
- Garrard, P.; Bradshaw, D.; Jager, H.R.; Thompson, A.J.; Losseff, N.; Playford, D. Cognitive dysfunction after isolated brain stem insult. An underdiagnosed cause of long term morbidity. J. Neurol. Neurosurg. Psychiatry 2002, 73, 191–194. [Google Scholar] [CrossRef] [PubMed]
- Hurley, R.A.; Flashman, L.A.; Chow, T.W.; Taber, K.H. The brainstem: Anatomy, assessment, and clinical syndromes. J. Neuropsychiatry Clin. Neurosci. 2010, 22, 1–7. [Google Scholar] [CrossRef]
- Del Campo, N.; Chamberlain, S.R.; Sahakian, B.J.; Robbins, T.W. The roles of dopamine and noradrenaline in the pathophysiology and treatment of attention-deficit/hyperactivity disorder. Biol. Psychiatry 2011, 69, e145–e157. [Google Scholar] [CrossRef]
- Hillyard, S.A.; Kutas, M. Electrophysiology of cognitive processing. Annu. Rev. Psychol. 1983, 34, 33–61. [Google Scholar] [CrossRef]
- Hansen, J.C.; Hillyard, S.A. Endogenous brain potentials associated with selective auditory attention. Electroencephalogr. Clin. Neurophysiol. 1980, 49, 277–290. [Google Scholar] [CrossRef]
- Naatanen, R. Selective attention and evoked potentials in humans—A critical review. Biol. Psychol. 1975, 2, 237–307. [Google Scholar] [CrossRef]
- Alho, K.; Rinne, T.; Herron, T.J.; Woods, D.L. Stimulus-dependent activations and attention-related modulations in the auditory cortex: A meta-analysis of fMRI studies. Hear. Res. 2014, 307, 29–41. [Google Scholar] [CrossRef]
- Varghese, L.; Bharadwaj, H.M.; Shinn-Cunningham, B.G. Evidence against attentional state modulating scalp-recorded auditory brainstem steady-state responses. Brain Res. 2015, 1626, 146–164. [Google Scholar] [CrossRef]
- Saiz-Alia, M.; Forte, A.E.; Reichenbach, T. Individual differences in the attentional modulation of the human auditory brainstem response to speech inform on speech-in-noise deficits. Sci. Rep. 2019, 9, 14131. [Google Scholar] [CrossRef]
- Forte, A.E.; Etard, O.; Reichenbach, T. The human auditory brainstem response to running speech reveals a subcortical mechanism for selective attention. eLife 2017, 6. [Google Scholar] [CrossRef]
- Naatanen, R.; Paavilainen, P.; Rinne, T.; Alho, K. The mismatch negativity (MMN) in basic research of central auditory processing: A review. Clin. Neurophysiol. 2007, 118, 2544–2590. [Google Scholar] [CrossRef]
- Woldorff, M.G.; Hackley, S.A.; Hillyard, S.A. The effects of channel-selective attention on the mismatch negativity wave elicited by deviant tones. Psychophysiology 1991, 28, 30–42. [Google Scholar] [CrossRef] [PubMed]
- Picton, T.W.; Woods, D.L.; Baribeau-Braun, J.; Healey, T.M. Evoked potential audiometry. J. Otolaryngol. 1976, 6, 90–119. [Google Scholar] [PubMed]
- Key, A.P.; Dove, G.O.; Maguire, M.J. Linking brainwaves to the brain: An ERP primer. Dev. Neuropsychol. 2005, 27, 183–215. [Google Scholar] [CrossRef] [PubMed]
- Hillyard, S.A.; Picton, T.W.; Regan, D.M. Sensation, perception and attention: Analysis using ERPs. In Event-Related Brain Potentials in Man; Callaway, E., Tueting, P., Koslow, S., Eds.; Academic: New York, NY, USA, 1978. [Google Scholar]
- Regev, M.; Simony, E.; Lee, K.; Tan, K.M.; Chen, J.; Hasson, U. Propagation of Information Along the Cortical Hierarchy as a Function of Attention While Reading and Listening to Stories. Cereb. Cortex 2019, 29, 4017–4034. [Google Scholar] [CrossRef] [PubMed]
- Oatman, L.C.; Anderson, B.W. Suppression of the auditory frequency following response during visual attention. Electroencephalogr. Clin. Neurophysiol. 1980, 49, 314–322. [Google Scholar] [CrossRef]
- Skoe, E.; Kraus, N. Auditory brain stem response to complex sounds: A tutorial. Ear Hear. 2010, 31, 302–324. [Google Scholar] [CrossRef] [PubMed]
- Bidelman, G.M. Subcortical sources dominate the neuroelectric auditory frequency-following response to speech. Neuroimage 2018, 175, 56–69. [Google Scholar] [CrossRef]
- Picton, T.W.; Hillyard, S.A.; Galambos, R.; Schiff, M. Human auditory attention: A central or peripheral process? Science 1971, 173, 351–353. [Google Scholar] [CrossRef]
- Woods, D.L.; Hillyard, S.A. Attention at the cocktail party: Brainstem evoked responses reveal no peripheral gating. In Multidisciplinary Perspectives in Event-Related Brain Potential Research; Otto, D., Ed.; GPO: Washington, DC, USA, 1978. [Google Scholar]
- Holmes, E.; Purcell, D.W.; Carlyon, R.P.; Gockel, H.E.; Johnsrude, I.S. Attentional Modulation of Envelope-Following Responses at Lower (93–109 Hz) but Not Higher (217–233 Hz) Modulation Rates. J. Assoc. Res. Otolaryngol. 2018, 19, 83–97. [Google Scholar] [CrossRef]
- Hartmann, T.; Weisz, N. Auditory cortical generators of the Frequency Following Response are modulated by intermodal attention. Neuroimage 2019, 203, 116185. [Google Scholar] [CrossRef]
- Galbraith, G.C.; Kane, J.M. Brainstem frequency-following responses and cortical event-related potentials during attention. Percept. Mot. Ski. 1993, 76, 1231–1241. [Google Scholar] [CrossRef]
- Bidelman, G.; Powers, L. Response properties of the human frequency-following response (FFR) to speech and non-speech sounds: Level dependence, adaptation and phase-locking limits. Int. J. Audiol. 2018, 665–672. [Google Scholar] [CrossRef]
- Coffey, E.B.; Herholz, S.C.; Chepesiuk, A.M.; Baillet, S.; Zatorre, R.J. Cortical contributions to the auditory frequency-following response revealed by MEG. Nat. Commun. 2016, 7, 11070. [Google Scholar] [CrossRef]
- Sohmer, H.; Feinmesser, M. Cochlear and cortical audiometry conveniently recorded in the same subject. ISR J. Med. Sci. 1970, 6, 219–223. [Google Scholar]
- Bidelman, G.M. Towards an optimal paradigm for simultaneously recording cortical and brainstem auditory evoked potentials. J. Neurosci. Methods 2015, 241, 94–100. [Google Scholar] [CrossRef]
- Musacchia, G.; Strait, D.; Kraus, N. Relationships between behavior, brainstem and cortical encoding of seen and heard speech in musicians and non-musicians. Hear. Res. 2008, 241, 34–42. [Google Scholar] [CrossRef]
- Bidelman, G.M.; Moreno, S.; Alain, C. Tracing the emergence of categorical speech perception in the human auditory system. Neuroimage 2013, 79, 201–212. [Google Scholar] [CrossRef]
- Krishnan, A.; Bidelman, G.M.; Smalt, C.J.; Ananthakrishnan, S.; Gandour, J.T. Relationship between brainstem, cortical and behavioral measures relevant to pitch salience in humans. Neuropsychologia 2012, 50, 2849–2859. [Google Scholar] [CrossRef]
- Bidelman, G.M.; Price, C.N.; Shen, D.; Arnott, S.R.; Alain, C. Afferent-efferent connectivity between auditory brainstem and cortex accounts for poorer speech-in-noise comprehension in older adults. Hear. Res. 2019, 382, 107795. [Google Scholar] [CrossRef]
- Bidelman, G.M.; Alain, C. Hierarchical neurocomputations underlying concurrent sound segregation: Connecting periphery to percept. Neuropsychologia 2015, 68, 38–50. [Google Scholar] [CrossRef]
- Bidelman, G.M.; Weiss, M.W.; Moreno, S.; Alain, C. Coordinated plasticity in brainstem and auditory cortex contributes to enhanced categorical speech perception in musicians. Eur. J. Neurosci. 2014, 40, 2662–2673. [Google Scholar] [CrossRef]
- Bidelman, G.M.; Alain, C. Musical Training Orchestrates Coordinated Neuroplasticity in Auditory Brainstem and Cortex to Counteract Age-Related Declines in Categorical Vowel Perception. J. Neurosci. 2015, 35, 1240–1249. [Google Scholar] [CrossRef]
- Bidelman, G.M.; Villafuerte, J.W.; Moreno, S.; Alain, C. Age-related changes in the subcortical-cortical encoding and categorical perception of speech. Neurobiol. Aging 2014, 35, 2526–2540. [Google Scholar] [CrossRef] [PubMed]
- Bidelman, G.M.; Davis, M.K.; Pridgen, M.H. Brainstem-cortical functional connectivity for speech is differentially challenged by noise and reverberation. Hear. Res. 2018, 367, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Presacco, A.; Simon, J.Z.; Anderson, S. Evidence of degraded representation of speech in noise, in the aging midbrain and cortex. J. Neurophysiol. 2016, 116, 2346–2355. [Google Scholar] [CrossRef]
- Presacco, A.; Simon, J.Z.; Anderson, S. Speech-in-noise representation in the aging midbrain and cortex: Effects of hearing loss. PLoS ONE 2019, 14, e0213899. [Google Scholar] [CrossRef]
- Bidelman, G.M.; Krishnan, A. Effects of reverberation on brainstem representation of speech in musicians and non-musicians. Brain Res. 2010, 1355, 112–125. [Google Scholar] [CrossRef]
- ASHA. Technical Report: (Central) Auditory Processing Disorders; American Speech-Language-Hearing Association: Rockville, MD, USA, 2005. [Google Scholar]
- Moore, D.R.; Ferguson, M.A.; Edmondson-Jones, A.M.; Ratib, S.; Riley, A. Nature of auditory processing disorder in children. Pediatrics 2010, 126, e382–e390. [Google Scholar] [CrossRef] [PubMed]
- Popescu, M.V.; Polley, D.B. Monaural deprivation disrupts development of binaural selectivity in auditory midbrain and cortex. Neuron 2010, 65, 718–731. [Google Scholar] [CrossRef]
- Polley, D.B.; Thompson, J.H.; Guo, W. Brief hearing loss disrupts binaural integration during two early critical periods of auditory cortex development. Nat. Commun. 2013, 4, 2547. [Google Scholar] [CrossRef]
- Andersson, M.; Llera, J.E.; Rimol, L.M.; Hugdahl, K. Using Dichotic Listening to Study Bottom-up and Top-down Processing in Children and Adults. Child. Neuropsychol. 2008, 14, 470–479. [Google Scholar] [CrossRef]
- Hiscock, M.; Kinsbourne, M. Attention and the right-ear advantage: What is the connection? Brain Cogn. 2011, 76, 263–275. [Google Scholar] [CrossRef]
- Kinsbourne, M. The cerebral basis of lateral asymmetries in attention. Acta Psychol. 1970, 33, 193–201. [Google Scholar] [CrossRef]
- Gagne, J.; Jennings, M.B. Incorporating a Client-Centered Approach in Audiologic Rehabilitation. ASHA Leader 2011, 16, 10–13. [Google Scholar] [CrossRef]
- Erdman, S.A. Audiologic counseling: A biopsychosocial approach. In Adult Audiologic Rehabilitation; Motano, J.J., Spitzer, J.B., Eds.; Plural: San Diego, CA, USA, 2009; pp. 171–215. [Google Scholar]

| Response | Attentional Effects? | Study | Results |
|---|---|---|---|
| Transient (ABR) | Yes | Lukas [34,35] | Enhanced amplitude and decreased latencies when attending to auditory stimuli |
| No | Picton, Hillyard, Galambos, & Schiff [102] | Enhanced cortical response but no attentional change observed in ABR | |
| No | Picton & Hillyard [36] | No significant attention-related changes in auditory evoked response until cortical N1-P2 ERP components | |
| No | Woods & Hillyard [103] | No change in ABR amplitude or latency as a function of attention | |
| Sustained (FFR) | Yes | Forte et al. [92] | More robust FFR reported when attending than when ignoring speech |
| Yes | Galbraith & Doan [74] | Enhanced FFR amplitudes with attention | |
| Yes | Galbraith et al. [73] | Enhanced FFR F0 for attended stimuli | |
| Yes | Galbraith et al. [37] | Attentional enhancements of overall FFR amplitude | |
| Mixed | Holmes et al. [104] | Effects of attention observed for low- (<110 Hz) but not high-frequency stimuli | |
| Mixed | Hartmann & Weisz [105] | Attentional modulation only within cortical generators of FFR—not brainstem | |
| No | Galbraith & Kane [106] | Enhancement of cortical ERPs but stable FFRs with attention | |
| No | Varghese et al. [90] | Attention-related enhancement noted in cortical responses but not FFR |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Price, C.N.; Moncrieff, D. Defining the Role of Attention in Hierarchical Auditory Processing. Audiol. Res. 2021, 11, 112-128. https://doi.org/10.3390/audiolres11010012
Price CN, Moncrieff D. Defining the Role of Attention in Hierarchical Auditory Processing. Audiology Research. 2021; 11(1):112-128. https://doi.org/10.3390/audiolres11010012
Chicago/Turabian StylePrice, Caitlin N., and Deborah Moncrieff. 2021. "Defining the Role of Attention in Hierarchical Auditory Processing" Audiology Research 11, no. 1: 112-128. https://doi.org/10.3390/audiolres11010012
APA StylePrice, C. N., & Moncrieff, D. (2021). Defining the Role of Attention in Hierarchical Auditory Processing. Audiology Research, 11(1), 112-128. https://doi.org/10.3390/audiolres11010012