
Antiviral Activity of Flavonoids from Bauhinia holophylla Leaves against Zika virus
 , , , ,
, , , ,  ,
,  ,
, 

Abstract
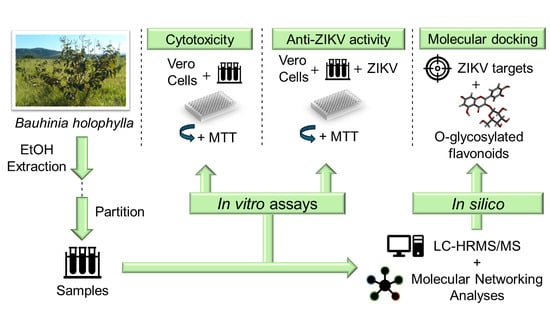
1. Introduction
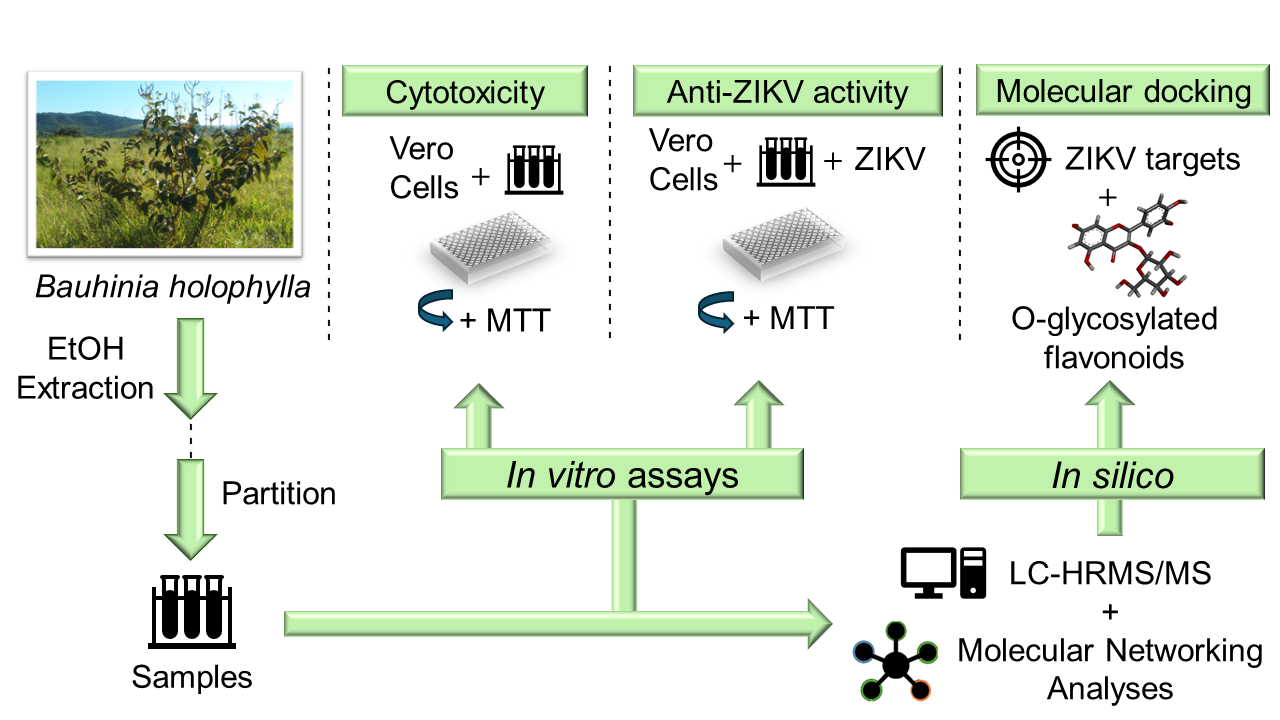
2. Materials and Methods
2.1. Chemicals
2.2. Plant Material, Extraction of Crude Extracts and Fractions, and Chemical Characterization
2.3. Cell Line and Virus
2.4. Determination of Cytotoxicity by the MTT Assay
2.5. Global Antiviral Assay
2.6. Global Natural Products Social Molecular Networking (GNPS)
2.7. Molecular Docking
2.8. Statistical Analysis
3. Results
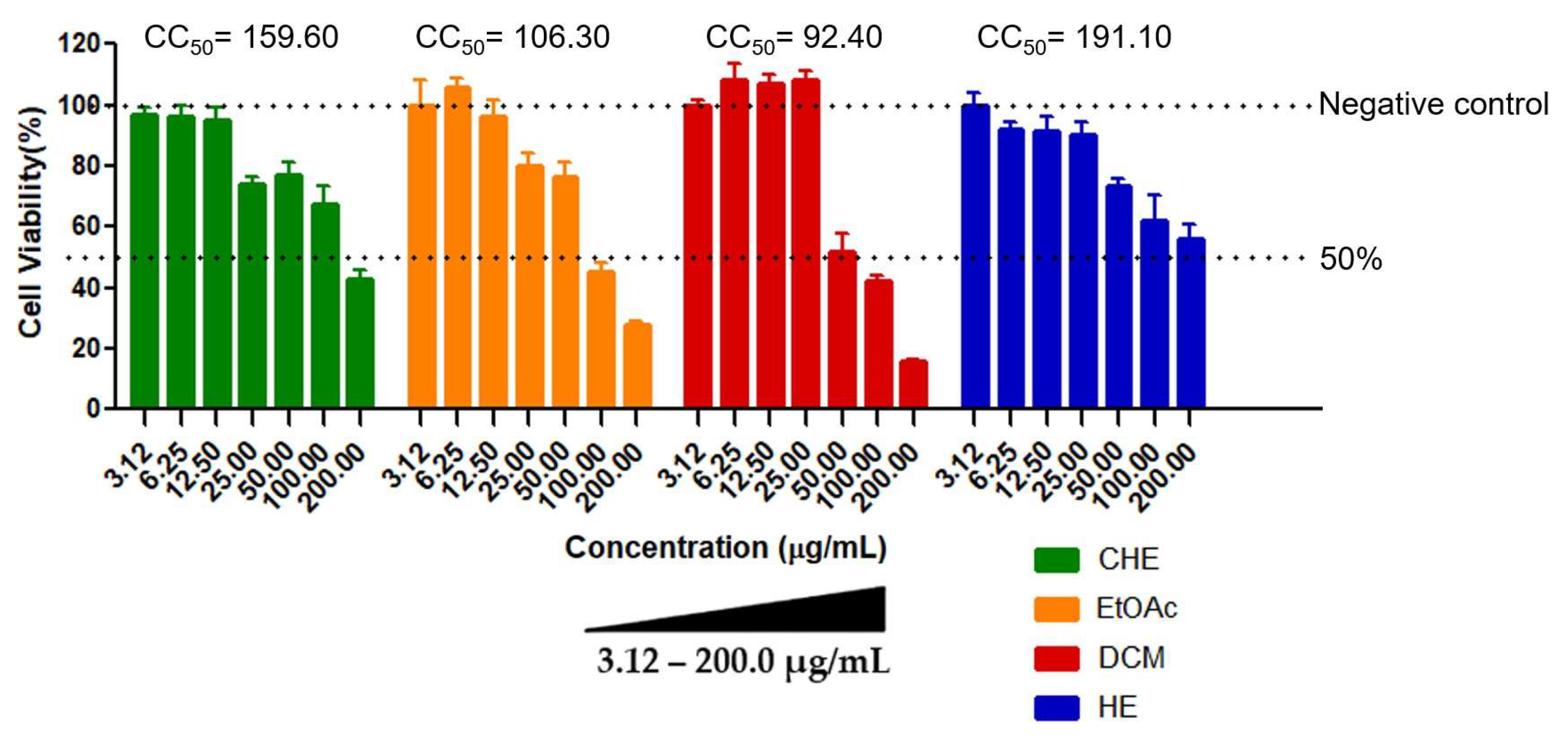
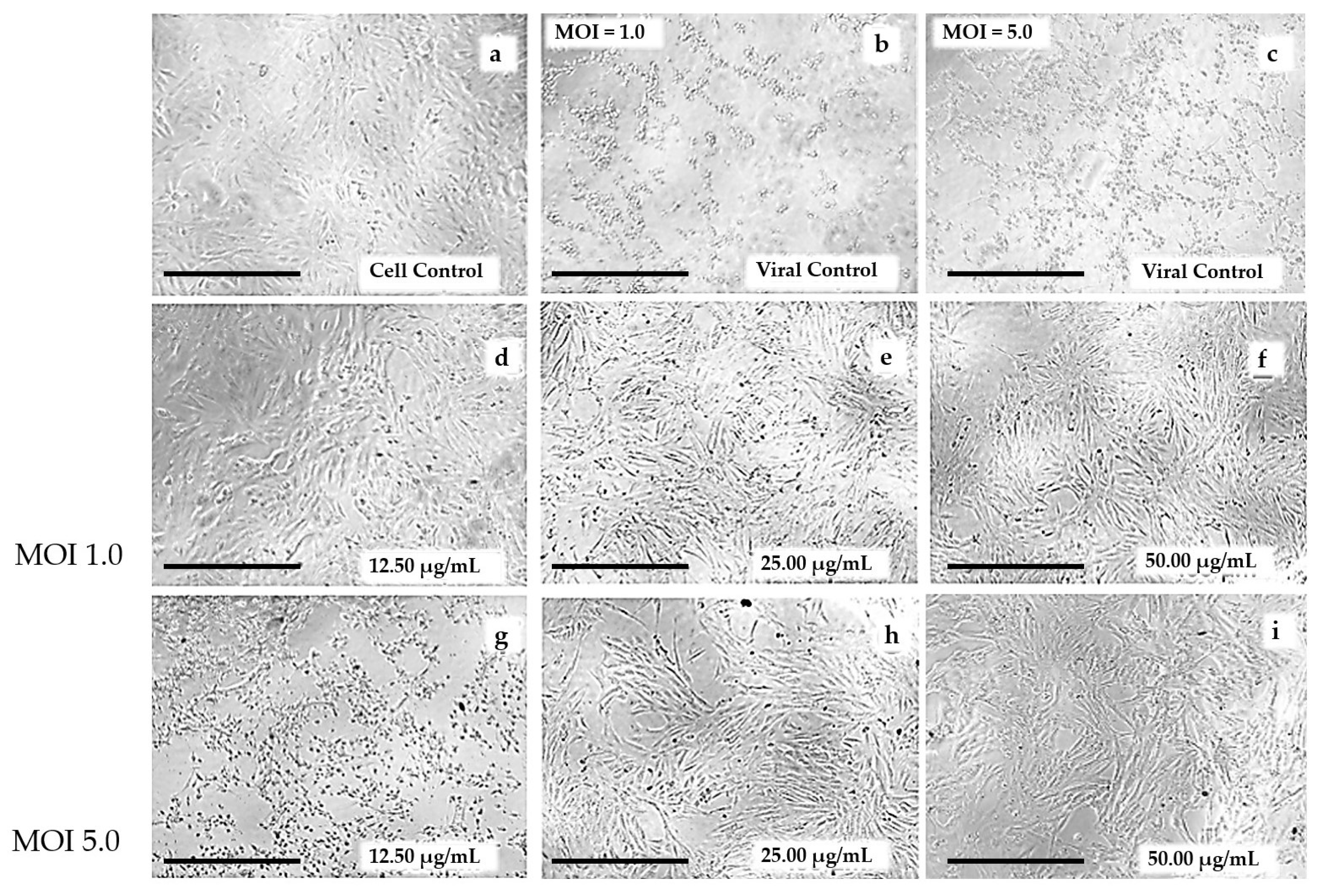
3.1. Determination of Cytotoxicity by the MTT Assay
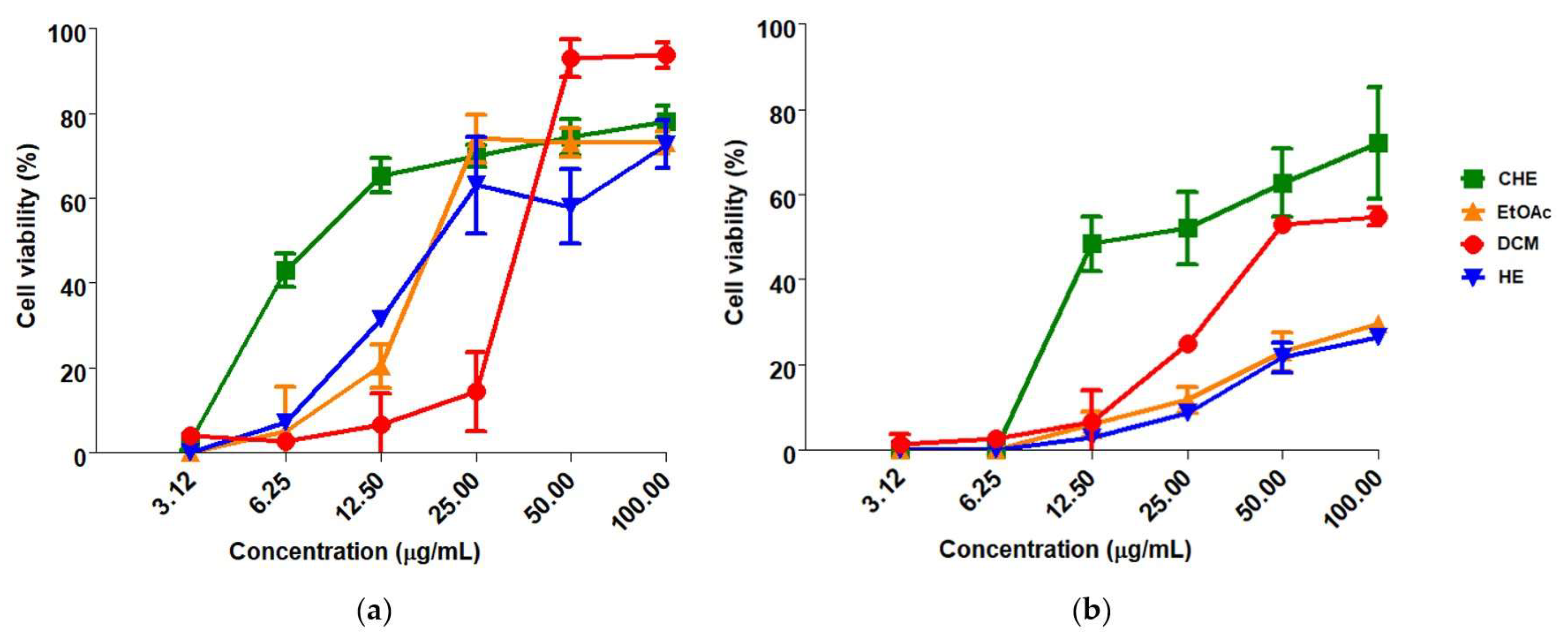
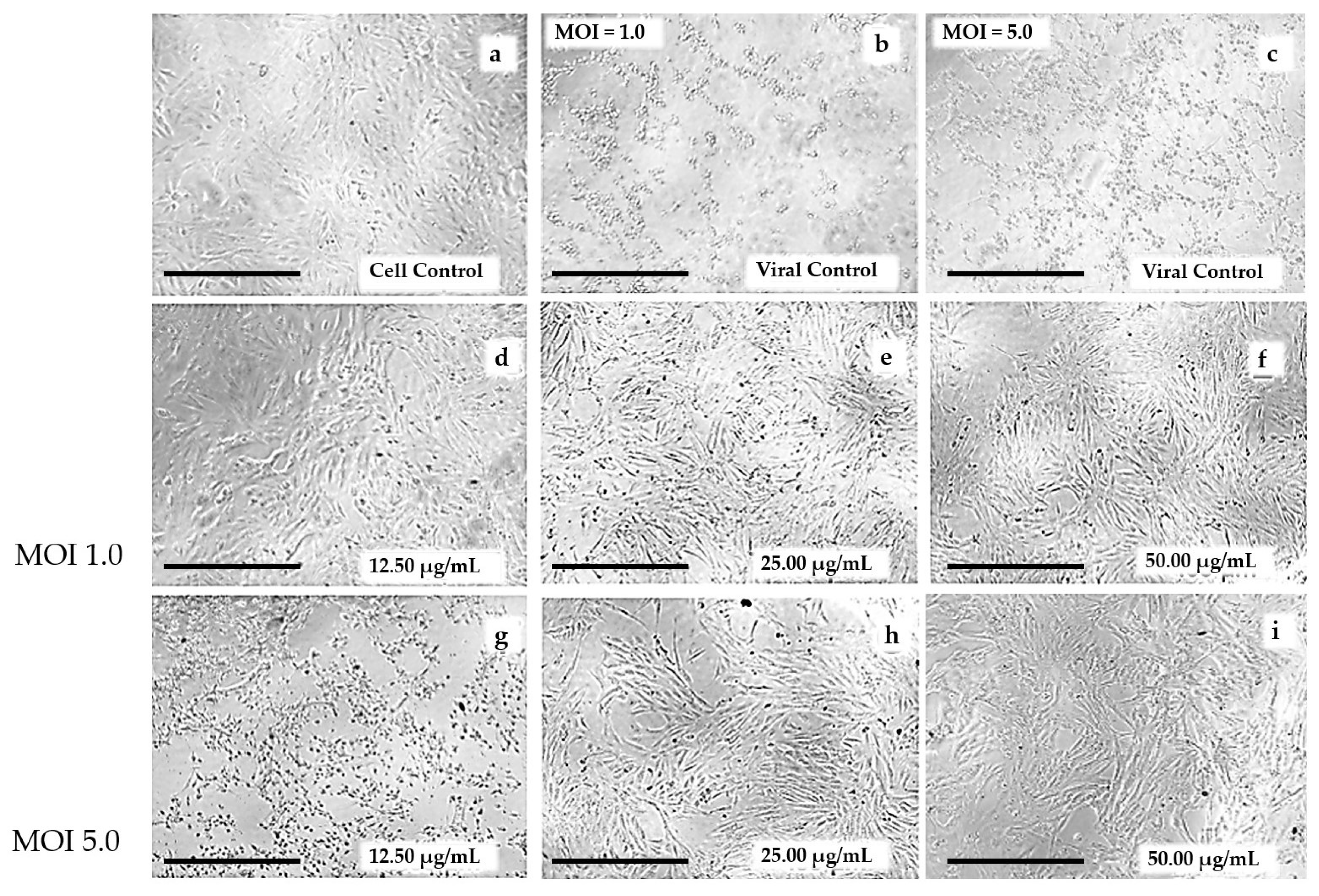
3.2. Global Antiviral Assay
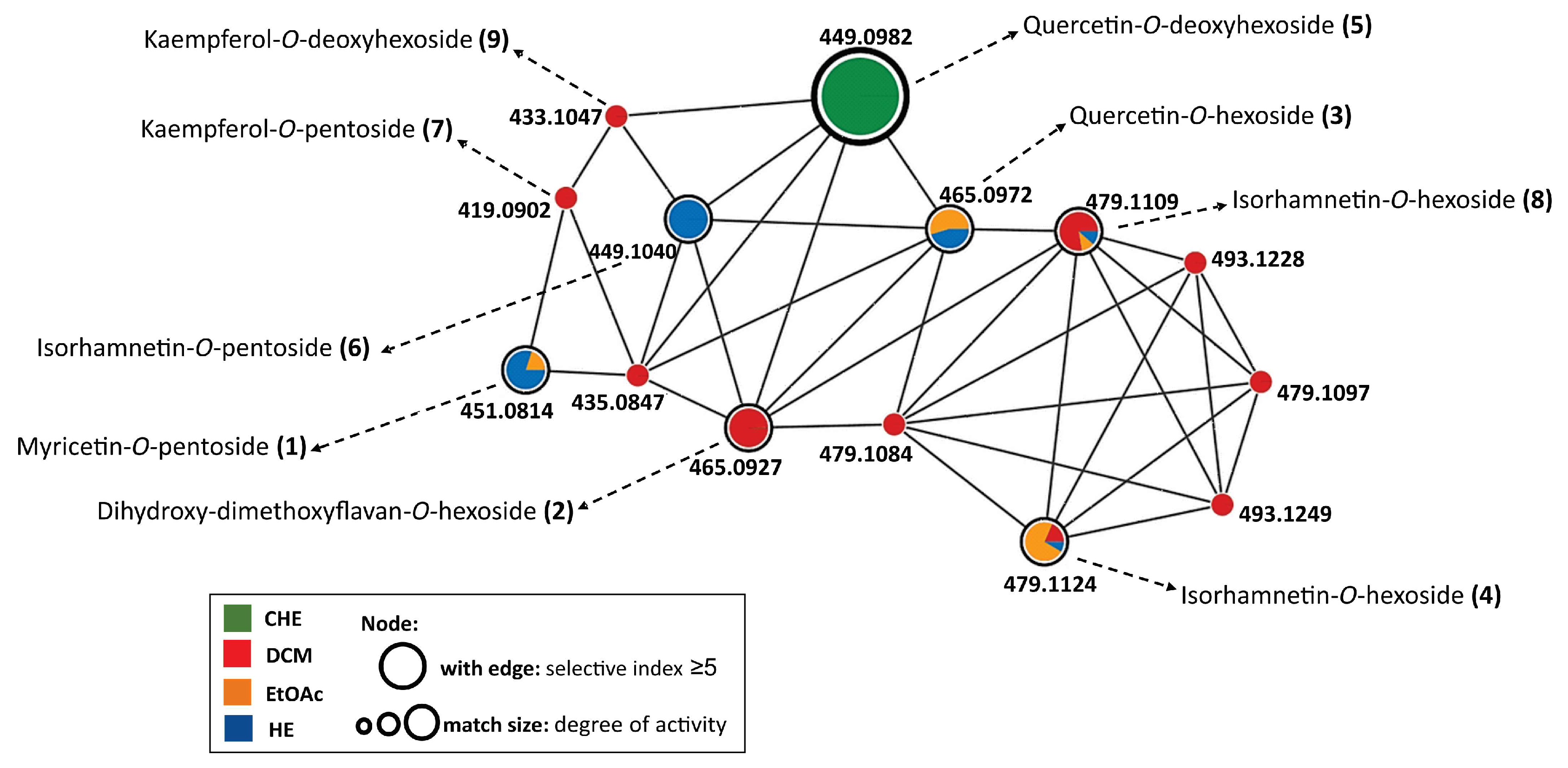
3.3. LC-HRMS/MS and Molecular Networking Analyses
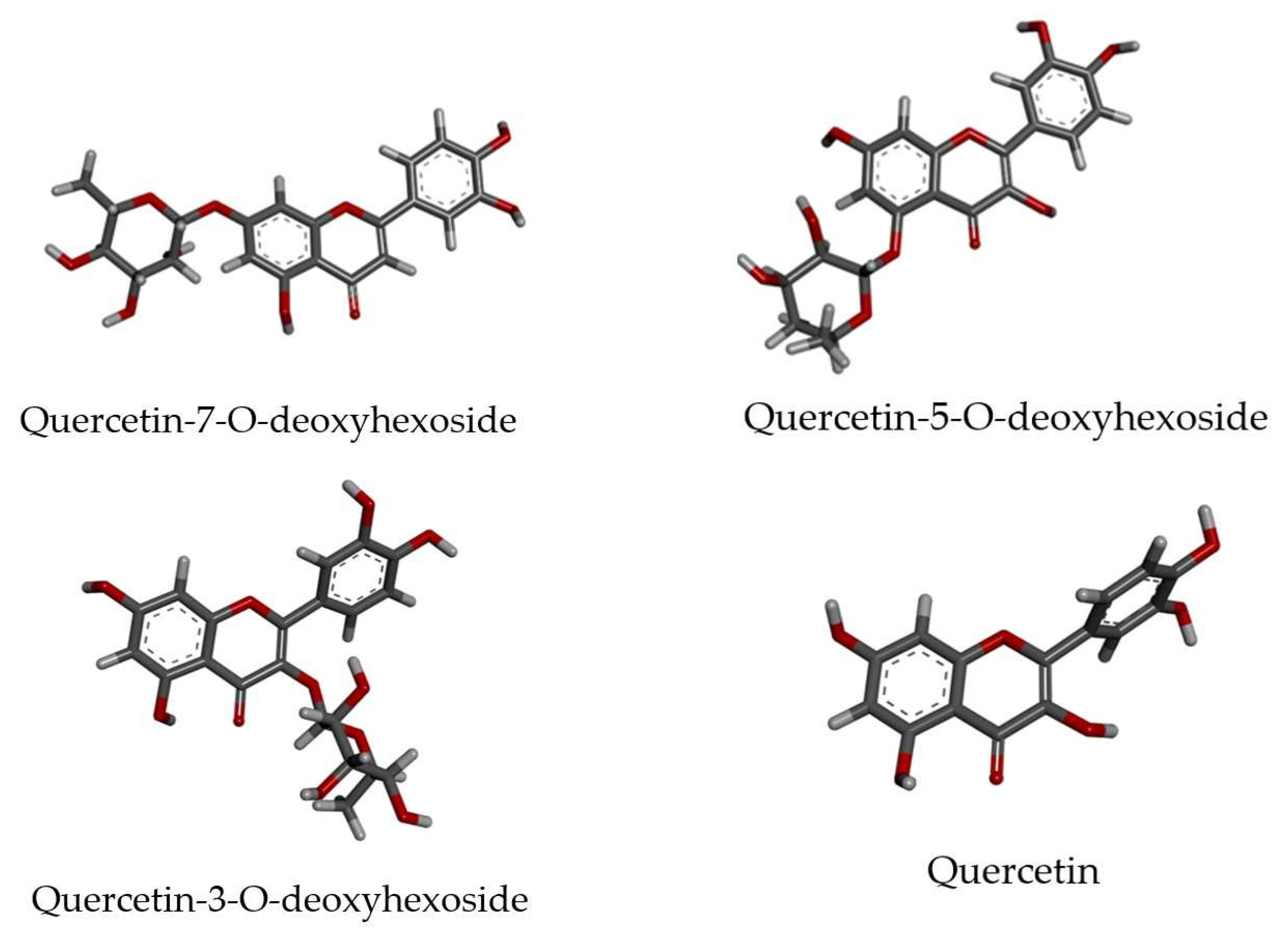
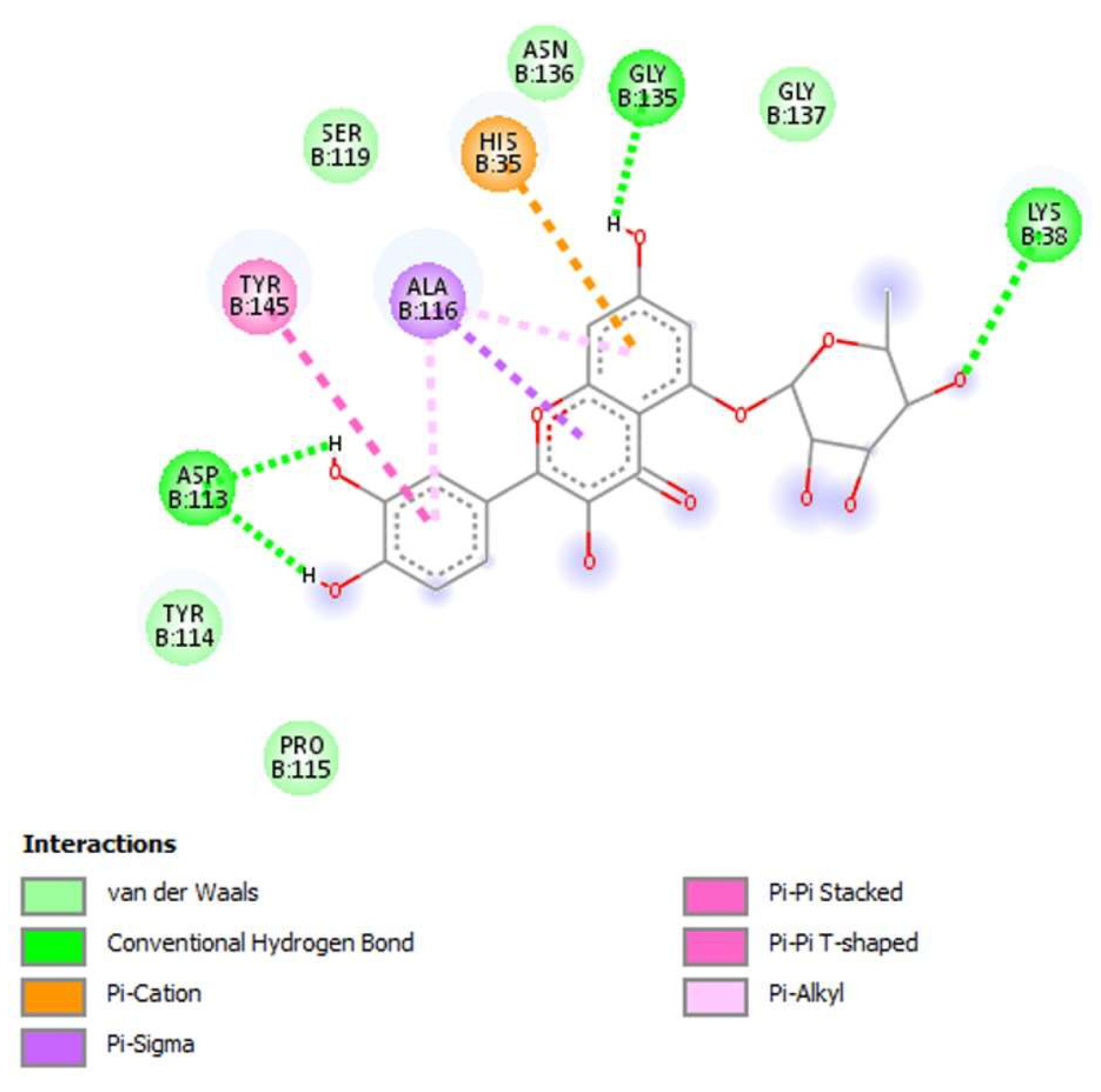
3.4. Molecular Docking
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Faye, O.; Freire, C.C.M.; Iamarino, A.; Faye, O.; de Oliveira, J.V.C.; Diallo, M.; Zanotto, P.M.A.; Sall, A.A. Molecular evolution of Zika virus during its emergence in the 20th century. PLoS Negl. Trop. Dis. 2014, 8, e2636. [Google Scholar] [CrossRef] [PubMed]
- Baud, D.; Gubler, D.J.; Schaub, B.; Lanteri, M.C.; Musso, D. An update on Zika virus infection. Lancet 2017, 390, 2099–2109. [Google Scholar] [CrossRef] [PubMed]
- Polen, K.D.; Gilboa, S.M.; Hills, S.; Oduyebo, T.; Kohl, K.S.; Brooks, J.T.; Adamski, A.; Simeone, R.M.; Walker, A.T.; Kissin, D.M.; et al. Update: Interim guidance for preconception counseling and prevention of sexual transmission of Zika virus for men with possible Zika virus exposure—United States. MMWR Morb. Mortal. Wkly. Rep. 2018, 67, 868–871. [Google Scholar] [CrossRef] [PubMed]
- Pielnaa, P.; Al-Saadawe, M.; Saro, A.; Dama, M.F.; Zhou, M.; Huang, Y.; Huang, J.; Xia, Z. Zika virus-spread, epidemiology, genome, transmission cycle, clinical manifestation, associated challenges, vaccine, and antiviral drug development. Virology 2020, 543, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Hennessey, M.; Fischer, M.; Staples, J.E. Zika virus spreads to new areas—Region of the Americas, May 2015–January 2016. MMWR Morb. Mortal. Wkly. Rep. 2016, 65, 55–58. [Google Scholar] [CrossRef] [PubMed]
- Petersen, L.R.; Jamieson, D.J.; Powers, A.M.; Honein, M.A. Zika virus. N. Eng. J. Med. 2016, 374, 1552–1563. [Google Scholar] [CrossRef] [PubMed]
- Oo, A.; Teoh, B.T.; Sam, S.S.; Bakar, S.A.; Zandi, K. Baicalein and baicalin as Zika virus inhibitors. Arch. Virol. 2018, 164, 585–593. [Google Scholar] [CrossRef] [PubMed]
- Musso, D.; Gubler, D.J. Zika virus. Clin. Microbiol. Rev. 2016, 29, 487–524. [Google Scholar] [CrossRef] [PubMed]
- Ventura, C.V.; Maia, M.; Bravo-Filho, V.; Góis, A.L.; Belfort, R., Jr. Zika virus in Brazil and macular atrophy in a child with microcephaly. Lancet 2016, 387, 228. [Google Scholar] [CrossRef]
- Meewan, I.; Shiryaev, S.A.; Kattoula, J.; Huang, C.T.; Lin, V.; Chuang, C.H.; Terskikh, A.V.; Abagyan, R. Allosteric inhibitors of Zika virus NS2B-NS3 protease targeting protease in “super-open” conformation. Viruses 2023, 15, 1106. [Google Scholar] [CrossRef]
- Silva, P.G.D.; Chaves, E.J.F.; Silva, T.M.S.; Rocha, G.B.; Dantas, W.M.; Oliveira, R.N.; Pena, L.J. Antiviral activity of flavonoids from geopropolis of the brazilian jandaira bee against Zika and Dengue viruses. Pharmaceutics 2023, 15, 2494. [Google Scholar] [CrossRef] [PubMed]
- da Vaz, A.M.S.; Bortoluzzi, R.L.C.; da Silva, L.A.E. Checklist of Bauhinia sensu stricto (Caesalpiniaceae) in Brazil. Plant Ecol. Evol. 2010, 143, 212–221. [Google Scholar] [CrossRef]
- Pinheiro, M.S.; Rodrigues, L.S.; Neto, L.S.; Moraes-Souza, R.Q.; Soares, T.S.; Americo, M.F. Effect of Bauhinia holophylla treatment in streptozotocin-induced diabetic rats. An. Acad. Bras. Cienc. 2017, 89, 263–272. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, M.; Teixeira, T.R.; Santos, F.R.S.; Lima, W.G.; Ferraz, A.C.; Silva, N.L.; Leite, F.J.; Siqueira, J.M.; Luyten, W.; Castro, A.H.F.; et al. Bauhinia holophylla (Bong.) Steud. leaves-derived extracts as potent anti-dengue serotype 2. Nat. Prod. Res. 2021, 35, 2804–2809. [Google Scholar] [CrossRef]
- Ribeiro, D.L.; Cilião, H.L.; Specian, A.F.L.; Serpeloni, J.M.; de Oliveira, M.T.; Varanda, E.A.; Vilegas, W.; Saldanha, L.L.; MartínezLópez, W.; Dokkedal, A.L.; et al. Phytochemical study and evaluation of cytotoxicity, mutagenicity, cell cycle kinetics and gene expression of Bauhinia holophylla (Bong.) Steud. in HepG2 cells in vitro. Cytotechnology 2018, 70, 713–728. [Google Scholar] [CrossRef] [PubMed]
- Marena, G.D.; Saldanha, L.L.; Ramos, M.A.S.; de Grandis, R.A.; Dokkedal, A.L.; Bauab, T.M.; Pavan, F.R.; Resende, F.A. Antimicrobial, cytotoxic and mutagenic activities of Bauhinia holophylla hydroalcoholic extract. Am. J. Essent. Oil. Nat. Prod. 2021, 9, 1–6. [Google Scholar]
- Wu, K.X.; Chu, J.J.H. Antiviral screen identifies EV71 inhibitors and reveals camptothecin-target, DNA topoisomerase 1 as a novel EV71 host factor. Antivir. Res. 2017, 143, 122–133. [Google Scholar] [CrossRef] [PubMed]
- Badshah, S.L.; Faisal, S.; Muhammad, A.; Poulson, B.G.; Emwas, A.H.; Jarenko, M. Antiviral activities of flavonoids. Biomed. Pharmacother. 2021, 140, 111596. [Google Scholar] [CrossRef] [PubMed]
- Di Petrillo, A.; Orrù, G.; Fais, A.; Fantini, M.C. Quercetin and its derivates as antiviral potentials: A comprehensive review. Phytother. Res. 2022, 36, 266–278. [Google Scholar] [CrossRef]
- Suebsaard, P.; Charerntantanakul, W. Rutin, α-tocopherol, and l-ascorbic acid up-regulate type I interferon-regulated gene and type I and II interferon expressions and reduce inflammatory cytokine expressions in monocyte-derived macrophages infected with highly pathogenic porcine reproductive and respiratory syndrome virus. Vet. Immunol. Immunopathol. 2021, 235, 110231. [Google Scholar] [CrossRef]
- Vissenaekens, H.; Criel, H.; Grootaert, C.; Raes, K.; Smagghe, G.; Van Camp, J. Flavonoids, and cellular stress: A complex interplay affecting human health. Crit. Rev. Food Sci. Nutr. 2022, 62, 8535–8566. [Google Scholar] [CrossRef] [PubMed]
- Chanu, N.R.; Gogoi, P.; Barbhuiya, P.A.; Dutta, P.P.; Pathak, M.P.; Sen, S. Natural flavonoids as potential therapeutics in the management of diabetic wound: A review. Curr. Top. Med. Chem. 2023, 23, 690–710. [Google Scholar] [CrossRef] [PubMed]
- Salatin, S.; Bazmani, A.; Shahi, S.; Naghili, B.; Memar, M.Y.; Dizaj, S.M. Antimicrobial benefits of flavonoids and their nanoformulations. Curr. Pharm. Des. 2022, 17, 1419–1432. [Google Scholar] [CrossRef] [PubMed]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [PubMed]
- Mendes, L.A.O.; Ponciano, C.S.; Caetano, A.H.D.; Wowk, P.F.; Bordignon, J.; Silva, H.; Almeida, M.V.; Ávila, E.P. The anti-Zika virus and anti-tumoral activity of the citrus flavanone lipophilic naringenin-based compounds. Chem. Biol. Interact. 2020, 331, 109218. [Google Scholar] [CrossRef]
- Eberle, R.J.; Olivier, D.S.; Pacca, C.C.; Avilla, C.M.S.; Nogueira, M.L.; Amaral, M.S.; Willbold, D.; Arni, R.K.; Coronado, M.A. In vitro study of Hesperetin and Hesperidin as inhibitors of Zika and Chikungunya virus proteases. PLoS ONE 2021, 16, e0246319. [Google Scholar] [CrossRef] [PubMed]
- Gaudry, A.; Bos, S.; Viranaicken, W.; Roche, M.; Krejbich-Trotot, P.; Gadea, G.; Desprès, P.; El-Kalamouni, C. The flavonoid isoquercitrin precludes initiation of Zika virus infection in human cells. Int. J. Mol. Sci. 2018, 19, 1093. [Google Scholar] [CrossRef] [PubMed]
- Quinn, R.A.; Nothias, L.F.; Vining, O.; Meehan, M.; Esquenazi, E.; Dorrestein, P.C. Molecular networking as a drug discovery, drug metabolism, and precision medicine strategy. Trends Pharmacol. 2017, 38, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Vincenti, F.; Montesano, C.; Di Ottavio, F.; Gregori, A.; Compagnone, D.; Sergi, M.; Dorrestein, P. Molecular networking: A useful tool for the identification of new psychoactive substances in seizures by LC-HRMS. Front. Chem. 2020, 8, 572952. [Google Scholar] [CrossRef]
- Schmid, R.; Petras, D.; Nothias, L.-F.; Wang, M.; Aron, A.T.; Jagels, A.; Tsugawa, H.; Rainer, J.; Garcia-Aloy, M.; Dührkop, K.; et al. Ion identity molecular networking for mass spectrometry-based metabolomics in the GNPS environment. Nat. Commun. 2021, 12, 3832. [Google Scholar] [CrossRef]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef]
- Xu, R.; Lee, J.; Chen, L.; Zhu, J. Enhanced detection, and annotation of small molecules in metabolomics using molecular-network oriented parameter optimization. Mol. Omics 2021, 17, 665–676. [Google Scholar] [CrossRef]
- Afoullouss, S.; Balsam, A.; Allcock, A.L.; Thomas, O.P. Optimization of LC-MS2 data acquisition parameters for molecular networking applied to marine natural products. Metabolites 2022, 12, 245. [Google Scholar] [CrossRef]
- Ferreira, L.G.; Dos Santos, R.N.; Oliva, G.; Andricopulo, A.D. Molecular docking and structure-based drug design strategies. Molecules 2015, 20, 13384–13421. [Google Scholar] [CrossRef] [PubMed]
- Wani, T.A.; Zargar, S.; Hussain, A. Spectroscopic, thermodynamic, and molecular docking studies on molecular mechanisms of drug binding to proteins. Molecules 2022, 27, 8405. [Google Scholar] [CrossRef] [PubMed]
- Pinzi, L.; Rastelli, G. Molecular docking: Shifting paradigms in drug discovery. Int. J. Mol. Sci. 2019, 20, 4331. [Google Scholar] [CrossRef] [PubMed]
- Agu, P.C.; Afiukwa, C.A.; Orji, O.U.; Ezeh, E.M.; Ofoke, I.H.; Ogbu, C.O.; Ugwuja, E.L.; Aja, P.M. Molecular docking as a tool for the discovery of molecular targets of nutraceuticals in diseases management. Sci. Rep. 2023, 13, 13398. [Google Scholar] [CrossRef] [PubMed]
- Da Fonseca, S.T.M.; Teixeira, T.T.; Siqueira, J.M.F.; Lima, L.A.R.S.; Luyten, W.; Castro, A.H.F. Flavonoid-rich fractions of Bauhinia holophylla leaves inhibit Candida albicans biofilm formation and hyphae growth. Plants 2022, 11, 1796. [Google Scholar] [CrossRef]
- Zandi, K.; Ramedani, E.; Mahammadi, K.; Tajbakhsh, S.; Deilami, I.; Rastian, Z.; Farshadpour, F. Evaluation of antiviral activities of curcumin derivatives against HSV-1 in Vero cell line. Nat. Prod. Commun. 2010, 5, 1935–1938. [Google Scholar] [CrossRef]
- Aguilar, M.G.D.; Sousa, G.F.D.; Evangelista, F.C.; Sabino, A.P.; Camargo, K.C.; Vieira Filho, S.A.; Nunes, Y.R.F.; Lucienir, P.; Duarte, L.P. Friedelane triterpenes with cytotoxic activity from the leaves of Maytenus quadrangulata (Celastraceae). Braz. Chem. Soc. 2022, 33, 1281–1290. [Google Scholar] [CrossRef]
- Dulbecco, R. Production of plaques in monolayer tissue cultures by single particles of an animal virus. Proc. Nat. Acad. Sci. USA 1952, 38, 747–752. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Osti, R.Z.; Serrano, F.A.; Paschoalin, T.; Massaoka, M.H.S.; Travassos, L.R. The in vitro and in vivo antitumour activities of nitrosyl ruthenium amine complexes. Aust. J. Chem. 2012, 65, 1333–1341. [Google Scholar] [CrossRef]
- Ferraz, A.C.; Moraes, T.D.F.S.; da Cruz Nizer, W.S.; Dos Santos, M.; Totola, A.H.; Ferreira, J.M.S.; de Magalhaes, J.C. Virucidal activity of proanthocyanidin against Mayaro virus. Antivir. Res. 2019, 168, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Brandão, G.C.; Kroon, E.G.; Santos, J.R.; Stehmann, J.R.; Lombardi, J.A.; Oliveira, A.B. Antiviral activities of plants occurring in the state of Minas Gerais, Brazil. Part 2. Screening Bignoniaceae species. Rev. Bras. Farmacogn. 2010, 20, 742–750. [Google Scholar] [CrossRef]
- Holman, J.D.; Tabb, D.L.; Mallick, P. Employing ProteoWizard to convert raw mass spectrometry data. Curr. Protoc. Bioinform. 2014, 46, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Aron, A.T.; Gentry, E.C.; McPhail, K.L.; Nothias, L.-F.; Nothias-Esposito, M.; Bouslimani, A.; Petras, D.; Gauglitz, J.M.; Sikora, N.; Vargas, F.; et al. Reproducible molecular networking of untargeted mass spectrometry data using GNPS. Nat. Protoc. 2020, 15, 1954–1991. [Google Scholar] [CrossRef] [PubMed]
- Ihlenfeldt, W.D.; Bolton, E.E.; Bryant, S.H. The PubChem chemical structure sketcher. J. Cheminform. 2009, 1, 20. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Cough, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrini, T.E. UCSF Chimera--a visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef]
- Guedes, I.A.; Barreto, A.M.S.; Marinho, D.; Krempser, E.; Kuenemann, M.A.; Sperandio, O.; Dardenne, L.E.; Miteva, M.A. New machine learning and physics-based scoring functions for drug discovery. Sci. Rep. 2021, 11, 3198. [Google Scholar] [CrossRef]
- Wolfender, J.L.; Waridel, P.; Ndjoko, K.; Hobby, K.R.; Major, H.J.; Hostettmann, K. Evaluation of Q-TOF-MS/MS and Multiple Stage IT-MSn for the Dereplication of Flavonoids and Related Compounds in Crude Plant Extracts. Analysis 2000, 28, 895–906. [Google Scholar] [CrossRef]
- Zaynab, M.; Fatima, M.; Abbas, S.; Sharif, Y.; Umair, M.; Zafar, M.H.; Bahadar, K. Role of secondary metabolites in plant defense against pathogens. Microb. Pathog. 2018, 124, 198–202. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Liu, Z.; Guo, Y.; Lu, S.; Du, H.; Cao, Y. Antioxidant capacity of flavonoids from Folium Artemisiae Argyi and the molecular mechanism in Caenorhabditis elegans. J. Ethnopharmacol. 2021, 279, 114398. [Google Scholar] [CrossRef] [PubMed]
- Aquino, A.J.; Pereira-Filho, E.R.; Oliveira, R.V.; Cass, Q.B. Chromatography conditions development by design of experiments for the chemotype differentiation of four Bauhinia species. Front. Chem. 2022, 10, 800729. [Google Scholar] [CrossRef] [PubMed]
- Chagas, M.D.S.S.; Behrens, M.D.; Moragas-Tellis, C.J.; Penedo, G.X.M.; Silva, A.R.; Gonçalves-de-Albuquerque, C.F. Flavonols and flavones as potential anti-inflammatory, antioxidant, and antibacterial compounds. Oxid. Med. Cell. Longev. 2022, 2022, 9966750. [Google Scholar] [CrossRef] [PubMed]
- Patel, K.; Kumar, V.; Rahman, M.; Verma, A.; Patel, D.K. New insights into the medicinal importance, physiological functions, and bioanalytical aspects of an important bioactive compound of foods ‘Hyperin’: Health benefits of the past, the present, the future. Beni-Suef Univ. J. Basic Appl. Sci. 2018, 7, 31–42. [Google Scholar] [CrossRef]
- Zhao, L.; Yuan, X.; Wang, J.; Feng, Y.; Ji, F.; Li, Z.; Bian, J. A review on flavones targeting serine/threonine protein kinases for potential anticancer drugs. Bioorg. Med. Chem. 2019, 27, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Phonghanpot, S.; Jarintanan, F. Antiproliferative, antibacterial, and antioxidant activities of Bauhinia strychnifolia Craib aqueous extracts in gut and liver perspective. BMC Complement. Med. Ther. 2021, 21, 276. [Google Scholar] [CrossRef] [PubMed]
- Da Rocha, P.S.; Orué, S.L.; Leite, D.F.; de Toledo Espindola, P.P.; Cassemiro, N.S.; Da Silva, D.B.; Carollo, C.A.; Nunes-Souza, V.; Rabelo, L.A.; Campos, J.F.; et al. Beneficial effects of Bauhinia rufa leaves on oxidative stress, prevention, and treatment of obesity in high-fat diet-fed c57bl/6 mice. Oxid. Med. Cell. Longev. 2022, 2014, 8790810. [Google Scholar] [CrossRef]
- Ansari, M.A.; Fatima, Z.; Hameed, S. Sesamol: A natural phenolic compound with promising anticandidal potential. J. Pathog. 2014, 2014, 895193. [Google Scholar] [CrossRef]
- Saldanha, L.L.; Delgado, A.Q.; Marcourt, L.; Camaforte, N.A.P.; Vareda, P.M.P.; Ebrahimi, S.N.; Vilegas, W.; Dokkedal, A.L.; Queiroz, E.F.; Wolfender, J.; et al. Hypoglycemic active principles from the leaves of Bauhinia holophylla: Comprehensive phytochemical characterization and in vivo activity profile. PLoS ONE. 2021, 16, e0258016. [Google Scholar] [CrossRef]
- Niksic, H.; Becic, F.; Koric, E.; Gúsic, I.; Omeragic, E.; Muratovic, S.; Miladinovic, B.; Duric, K. Cytotoxicity screening of Thymus vulgaris L. essential oil in brine shrimp nauplii and cancer cell lines. Sci. Rep. 2021, 11, 13178. [Google Scholar] [CrossRef] [PubMed]
- Indrayanto, G.; Putra, G.S.; Suhud, F. Validation of In-Vitro Bioassay Methods: Application in Herbal Drug Research. Profiles Drug Subst. Excip. Relat. Methodol. 2021, 46, 273–307. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhang, D.; Li, C.; Zheng, Z.; Fu, M.; Ni, F.; Liu, Y.; Du, T.; Wang, H.; Griffin, G.E.; et al. Characterization of Zika virus endocytic pathways in human glioblastoma cells. Front. Microb. 2020, 11, 242. [Google Scholar] [CrossRef] [PubMed]
- Bernardo-Menezes, L.C.; Agrelli, A.; Oliveira, A.S.L.E.; Azevedo, E.A.N.; Morais, C.N.L. Zika virus: Critical crosstalk between pathogenesis, cytopathic effects, and macroautophagy. J. Cell. Biochem. 2023. [Google Scholar] [CrossRef] [PubMed]
- Nothias, L.-F.; Petras, D.; Schmid, R.; Dührkop, K.; Rainer, J.; Sarvepalli, A.; Protsyuk, I.; Ernst, M.; Tsugawa, H.; Fleischauer, M.; et al. Feature-based molecular networking in the GNPS analysis environment. Nat. Methods 2020, 17, 905–908. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.M.; Li, Y.Y.; Shi, Y.W.; Fang, Y.W.; Chao, R.; Gu, Y.C.; Wang, C.Y.; Shao, C.L. Integrating molecular networking and 1H NMR to target the isolation of Chrysogeamides from a library of marine derived Penicillium fungi. J. Org. Chem. 2019, 84, 1228–1237. [Google Scholar] [CrossRef] [PubMed]
- Olivon, F.; Allard, P.M.; Koval, A.; Righi, D.; Genta-Jouve, G.; Neyts, J.; Apel, C.; Pannecouque, C.; Nothias, L.F.; Cachet, X.; et al. Bioactive natural products prioritization using massive multi-informational molecular networks. ACS Chem. Biol. 2017, 12, 2644–2651. [Google Scholar] [CrossRef] [PubMed]
- Qin, G.F.; Zhang, X.; Zhu, F.; Huo, Z.Q.; Yao, Q.Q.; Feng, Q.; Liu, Z.; Zhang, G.M.; Yao, J.C.; Liang, H.B. MS/MS-Based molecular networking: An efficient approach for natural products dereplication. Molecules 2022, 28, 157. [Google Scholar] [CrossRef]
- Ramabulana, A.T.; Petras, D.; Madala, N.E.; Tugizimana, F. Metabolomics and molecular networking to characterize the chemical space of four Momordica plant species. Metabolites 2021, 11, 763. [Google Scholar] [CrossRef]
- Ramatapa, T.; Msobo, A.; Maphari, P.W.; Ncube, E.N.; Nogemane, N.; Mhlongo, M.I. Identification of plant-derived bioactive compounds using affinity mass spectrometry and molecular networking. Metabolites 2022, 12, 863. [Google Scholar] [CrossRef]
- Wong, G.; He, S.; Siragam, V.; Bi, Y.; Mbikay, M.; Chretien, M.; Qiu, X. Antiviral activity of quercetin-3-β-O-D-glucoside against Zika virus infection. Virol. Sin. 2017, 32, 545–547. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, A.E.; Kuster, R.M.; Yamamoto, K.A.; Salles, T.S.; Campos, R.; De Meneses, M.D.F.; Soares, M.R.; Ferreira, D. Quercetin and quercetin 3-O-glycosides from Bauhinia longifolia (Bong.) Steud. show anti-Mayaro virus activity. Parasit. Vectors 2014, 7, 130. [Google Scholar] [CrossRef] [PubMed]
- Loaiza-Cano, V.; Monsalve-Escudero, L.M.; Filho, C.S.B.; Martinez-Gutierrez, M.; Sousa, D.P. Antiviral role of phenolic compounds against Dengue virus: A review. Biomolecules 2020, 11, 11. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, V.; Brognaro, H.; Prabhu, P.R.; de Souza, E.E.; Günther, S.; Reinke, P.Y.A.; Lane, T.J.; Ginn, H.; Han, H.; Ewert, W.; et al. Antiviral activity of natural phenolic compounds in complex at an allosteric site of SARS-CoV-2 papain-like protease. Commun. Biol. 2022, 5, 805. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.S.; Santos, F.C.P.; Campana, P.R.V.; Costa, V.V.; de Pádua, R.M.; Souza, D.G.; Teixeira, M.M.; Braga, F.C. Natural products, and derivatives as potential Zika virus inhibitors: A comprehensive review. Viruses 2023, 15, 1211. [Google Scholar] [CrossRef]
- Santos, F.R.S.; Lima, W.G.; Maia, E.H.B.; Assis, L.C.; Davyt, D.; Taranto, A.G.; Ferreira, J.M.S. Identification of a potential Zika Virus inhibitor targeting NS5 methyltransferase using virtual screening and molecular dynamics simulations. J. Chem. Inf. Model. 2020, 24, 562–568. [Google Scholar] [CrossRef] [PubMed]
- Santos, F.R.S.; Nunes, D.A.F.; Lima, W.G.; Davyt, D.; Santos, L.; Taranto, A.G.; Ferreira, J.M.S. Identification of Zika virus NS2B-NS3 protease inhibitors by structure-based virtual screening and drug repurposing approaches. J. Chem. Inf. Model. 2019, 24, 731–737. [Google Scholar] [CrossRef] [PubMed]
- Zou, M.; Liu, H.; Li, J.; Yao, X.; Chen, Y.; Ke, C.; Liu, S. Structure-activity relationship of flavonoid bifunctional inhibitors against Zika virus infection. Biochem. Pharmacol. 2020, 177, 113962. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Choi, H.; Kim, Y.B. Therapeutic targets, and biological mechanisms of action of curcumin against Zika virus: In silico and in vitro analyses. Eur. J. Pharmacol. 2021, 904, 174144. [Google Scholar] [CrossRef]
- Lim, H.-J.; Nguyen, T.T.H.; Kim, N.M.; Park, J.S.; Jang, T.S.; Kim, D. Inhibitory effect of flavonoids against NS2B-NS3 protease of Zika virus and their structure activity relationship. Biotechnol. Lett. 2017, 39, 415. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | CC50 f (µg/mL) | EC50 g (µg/mL) | SI h | ||
|---|---|---|---|---|---|
| MOI 1.0 | MOI 5.0 | MOI 1.0 | MOI 5.0 | ||
| CHE a | 159.60 ± 3.52 | 11.93 ± 4.74 | 28.59 ± 16.21 | 13.38 | 5.58 |
| EtOAc b | 106.31 ± 4.71 | 25.49 ± 10.40 | NE e | 4.17 | NE e |
| DCM c | 92.40 ± 3.12 | 31.37 ± 11.32 | NE e | 2.94 | NE e |
| HE d | 191.10 ± 4.15 | 35.65 ± 21.62 | NE e | 5.36 | NE e |
| ID | Rt * (min) | Molecular Formula | [M + H]+ (m/z) Theoretical | [M + H]+ (m/z) Experimental | Error (ppm) | MS/MS (MS2) a | Proposed Compound | Area CHE b | Area DCM c | Area EtOA d | Area HE e |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 12.23 | C20H18O12 | 451.0865 | 451.0814 | 11.3 | 319 (-Pen) | Myricetin-O-pentoside | - | - | 8.0E3 | 4.1E4 |
| 2 | 13.00 | C21H20O12 | 465.1022 | 465.0927 | 20.4 | 303 (-Hex), 153, 125, 121 | Dihydroxy-dimethoxyflavan-O-hexoside | 2.9E3 | 8.0E5 | - | - |
| 3 | 13.19 | C21H20O12 | 465.1022 | 465.0972 | 10.8 | 303 (-Hex), 153, 165, 137 | Quercetin-O-hexoside | - | - | 4.2E5 | 3.5E5 |
| 4 | 16.36 | C22H22O12 | 479.1178 | 479.1124 | 11.3 | 317 (-Hex) | Isorhamnetin-3-O-hexoside | - | 8.3E3 | 3.2E4 | 3.8E3 |
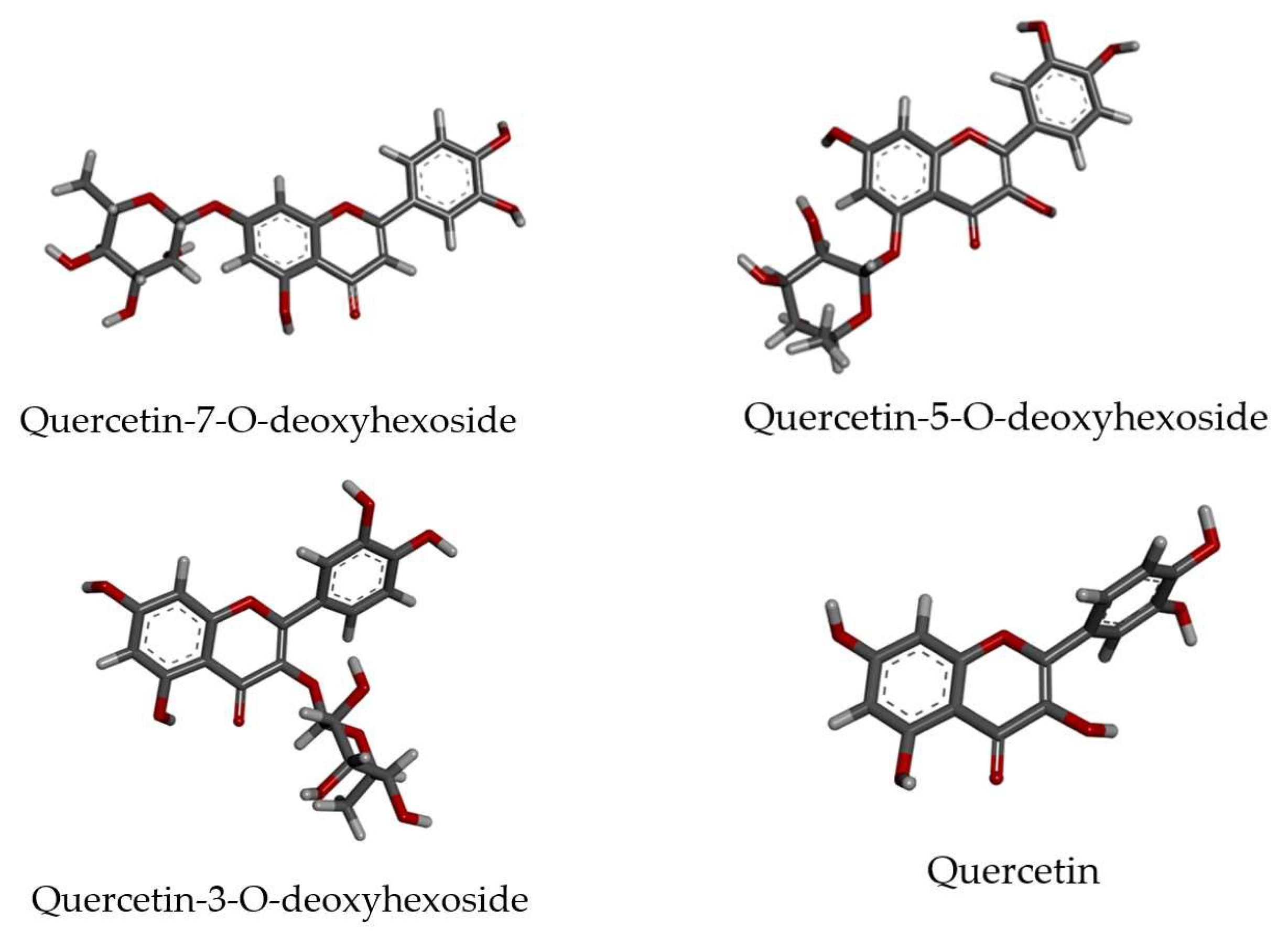
| 5 | 16.45 | C21H20O11 | 449.1073 | 449.0982 | 20.3 | 303 (-dHex), 153, 165, 137 | Quercetin-O-deoxyhexoside | 1.7E4 | - | - | - |
| 6 | 16.72 | C21H20O11 | 449.1073 | 449.1040 | 7.3 | 317 (-Pen) | Isorhamnetin-O-pentoside | - | - | - | 3.0E5 |
| 7 | 18.13 | C20H18O10 | 419.0967 | 419.0902 | 15.5 | 287 (-Pen) | Kaempferol-O-pentoside | - | 5.6E4 | - | - |
| 8 | 18.54 | C22H22O12 | 479.1178 | 479.1109 | 14.4 | 317 (-Hex) | Isorhamnetin-3-O-hexoside | - | 2.2E5 | 3.1E4 | 3.3E4 |
| 9 | 18.75 | C21H20O10 | 433.1123 | 433.1047 | 17.5 | 287 (-dHex) | Kaempferol-O-deoxyhexoside | - | 1.1E5 | - | - |
| Target | Ligand | Interaction Energy (Kcal/mol) |
|---|---|---|
| NS2B-NS3 protease a | Quercetin-5-O-deoxyhexoside | −33.06 |
| Quercetin-3-O-deoxyhexoside | −30.23 | |
| Quercetin-7-O-deoxyhexoside | −29.94 | |
| Quercetin | −26.88 | |
| Benzimidazol-1ylmethanol * | −21.63 | |
| NS3 helicase b | 5′-guanosine-diphosphate-monothiophosphate * | −60.42 |
| Quercetin-5-O-deoxyhexoside | −39.02 | |
| Quercetin | −35.44 | |
| Quercetin-7-O-deoxyhexoside | −33.62 | |
| Quercetin-3-O-deoxyhexoside | −7.50 | |
| NS5 methyltransferase c | S-adenosylmethionine * | −71.34 |
| Quercetin-7-O-deoxyhexoside | −50.08 | |
| Quercetin-5-O-deoxyhexoside | −49.72 | |
| Quercetin-3-O-deoxyhexoside | −46.25 | |
| Quercetin | −42.22 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thomasi, R.M.d.O.; Teixeira, T.R.; Lopes, G.F.M.; Mendonça, S.C.; Gomes, B.A.; Leitão, S.G.; Oliveira, T.A.d.; Fonseca, S.T.D.d.; Taranto, A.G.; Ferreira, J.M.S.; et al. Antiviral Activity of Flavonoids from Bauhinia holophylla Leaves against Zika virus. Microbiol. Res. 2024, 15, 582-597. https://doi.org/10.3390/microbiolres15020038
Thomasi RMdO, Teixeira TR, Lopes GFM, Mendonça SC, Gomes BA, Leitão SG, Oliveira TAd, Fonseca STDd, Taranto AG, Ferreira JMS, et al. Antiviral Activity of Flavonoids from Bauhinia holophylla Leaves against Zika virus. Microbiology Research. 2024; 15(2):582-597. https://doi.org/10.3390/microbiolres15020038
Chicago/Turabian StyleThomasi, Rodrigo Michelini de Oliveira, Thaiz Rodrigues Teixeira, Gabriela Francine Martins Lopes, Simony Carvalho Mendonça, Brendo Araujo Gomes, Suzana Guimarães Leitão, Tiago Alves de Oliveira, Sara Thamires Dias da Fonseca, Alex Gutterres Taranto, Jaqueline Maria Siqueira Ferreira, and et al. 2024. "Antiviral Activity of Flavonoids from Bauhinia holophylla Leaves against Zika virus" Microbiology Research 15, no. 2: 582-597. https://doi.org/10.3390/microbiolres15020038
APA StyleThomasi, R. M. d. O., Teixeira, T. R., Lopes, G. F. M., Mendonça, S. C., Gomes, B. A., Leitão, S. G., Oliveira, T. A. d., Fonseca, S. T. D. d., Taranto, A. G., Ferreira, J. M. S., dos Santos Lima, L. A. R., & Castro, A. H. F. (2024). Antiviral Activity of Flavonoids from Bauhinia holophylla Leaves against Zika virus. Microbiology Research, 15(2), 582-597. https://doi.org/10.3390/microbiolres15020038