

The Secretome of Human Deciduous Tooth-Derived Mesenchymal Stem Cells Enhances In Vitro Wound Healing and Modulates Inflammation


 , , ,
, , ,  ,
,  and
and
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
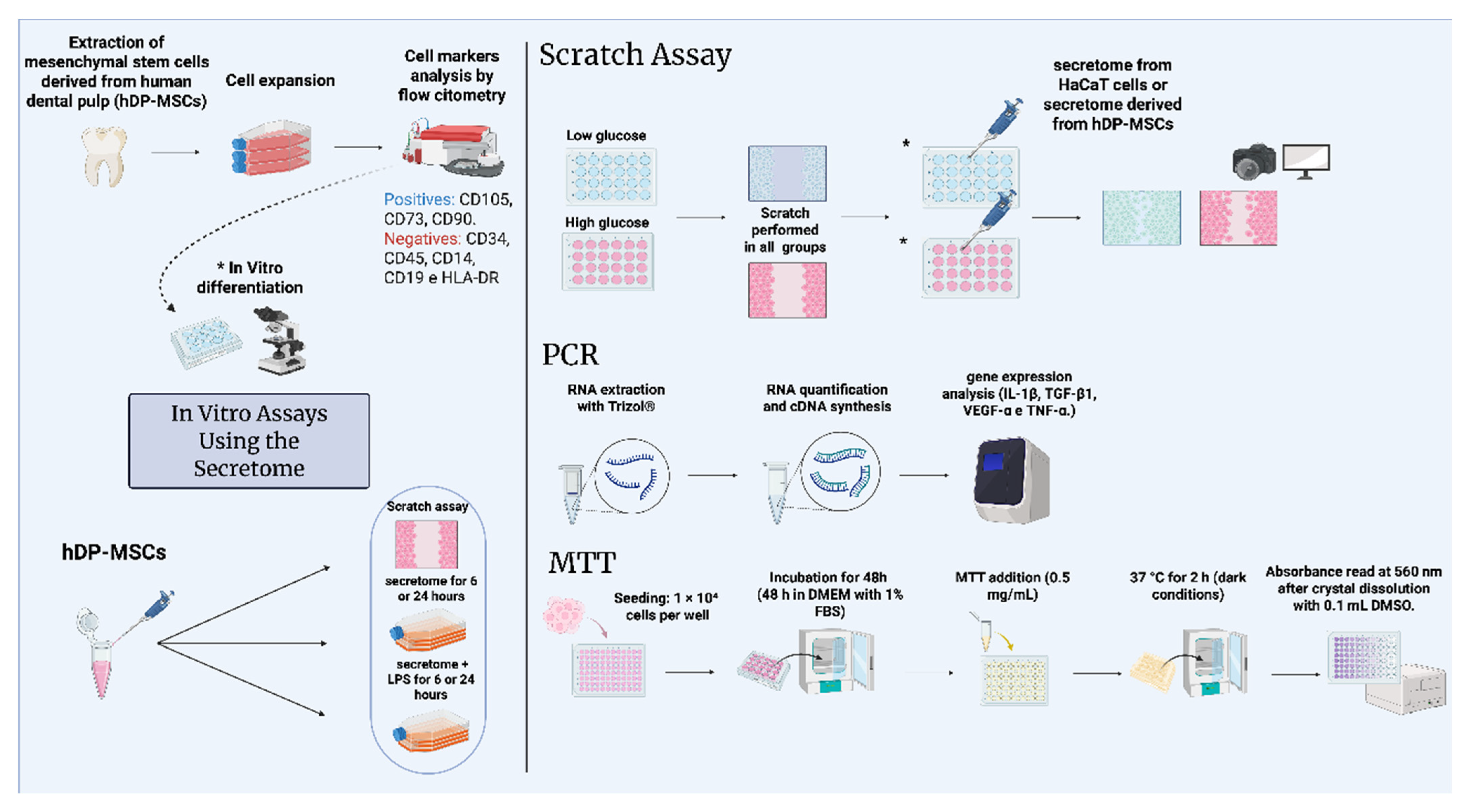
2. Materials and Methods
2.1. Sampling and Isolation of Mesenchymal Stem Cells Derived from Deciduous Dental Pulp
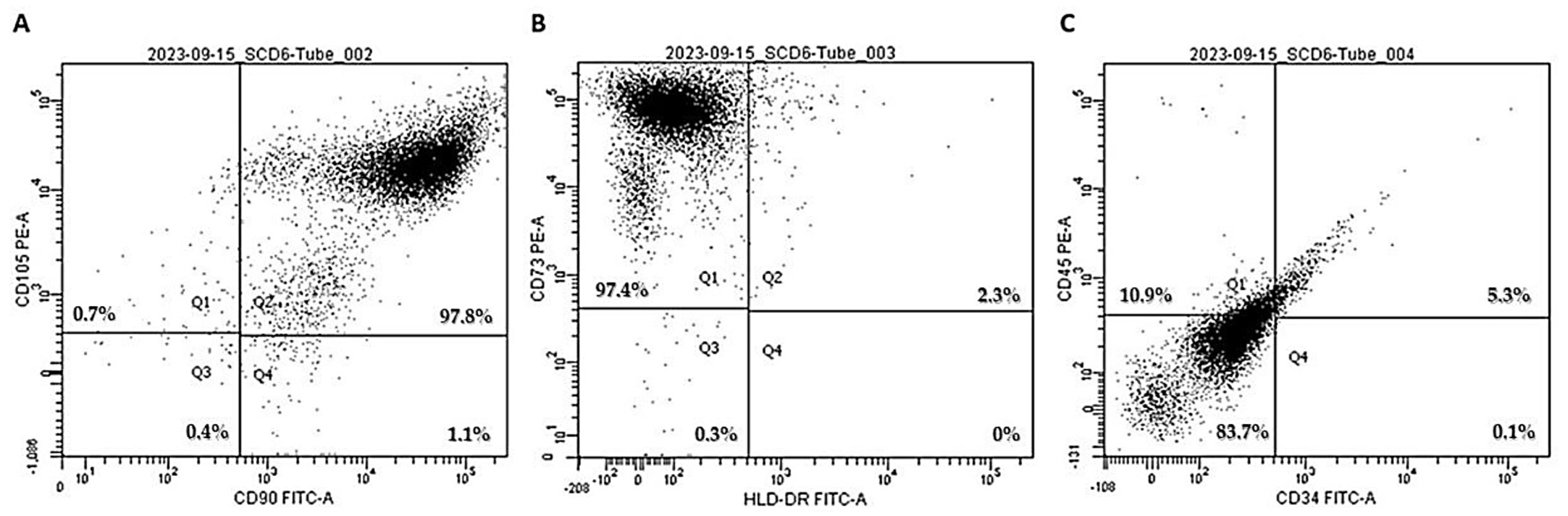
2.1.1. Phenotyping of Human Mesenchymal Stem Cells Isolated from Deciduous Dental Pulp
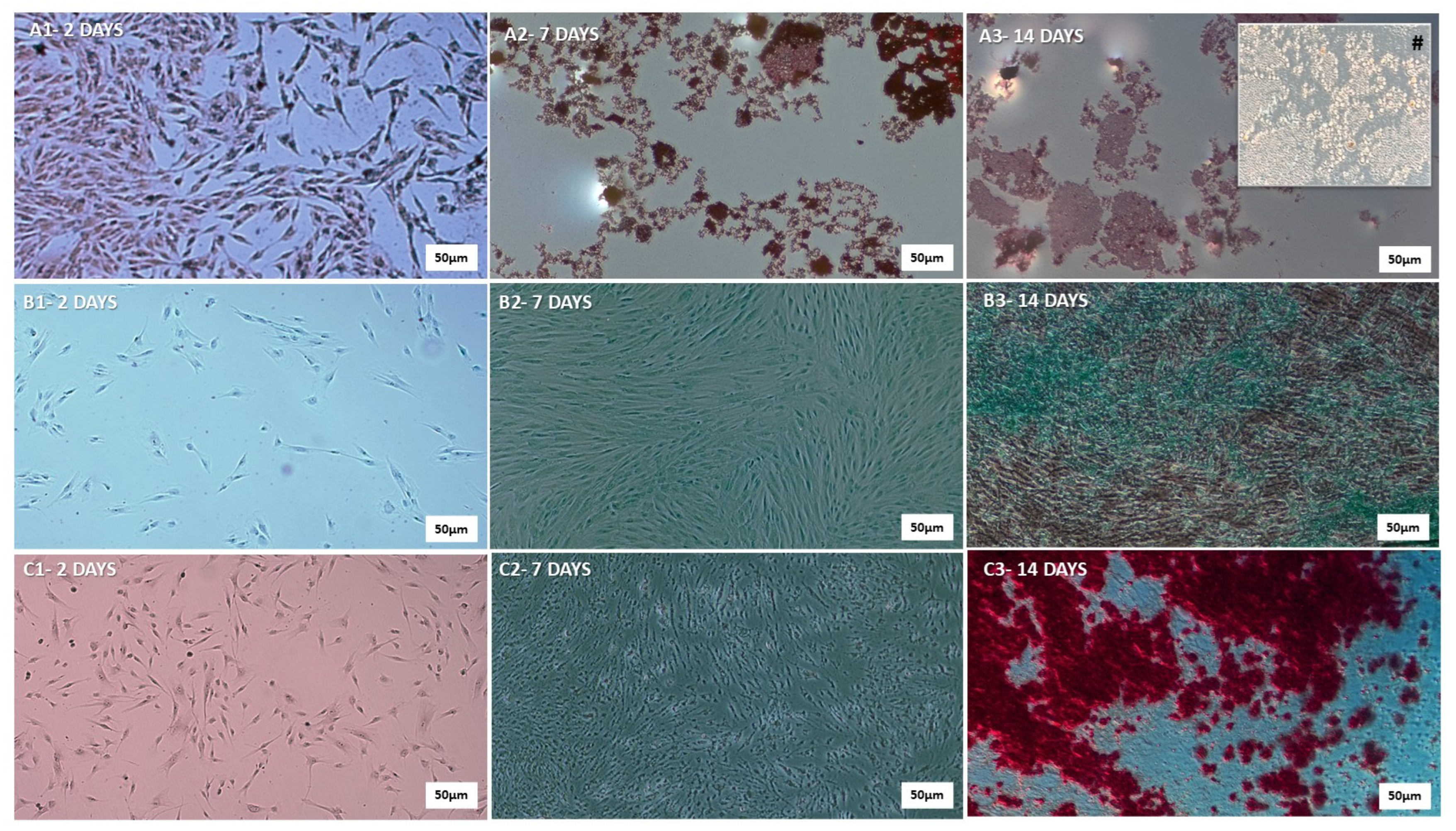
2.1.2. Confirmation of Human Mesenchymal Stem Cell Differentiation Potential
2.1.3. Cell Culture and Collection of Conditioned Medium (Secretome)
2.1.4. HaCaT Cell Culture, Secretome Treatment, and Inflammation Induction
2.1.5. Scratch Assay and Quantification of Scratch Closure in HaCaT Cells
2.1.6. RT-qPCR Analysis
2.1.7. Quantitative PCR
2.1.8. Cell Viability Analysis Using an MTT Assay
2.1.9. Statistical Analysis of Results
3. Results
3.1. Confirmation of hDP-MSC Identity as Human Mesenchymal Stem Cells
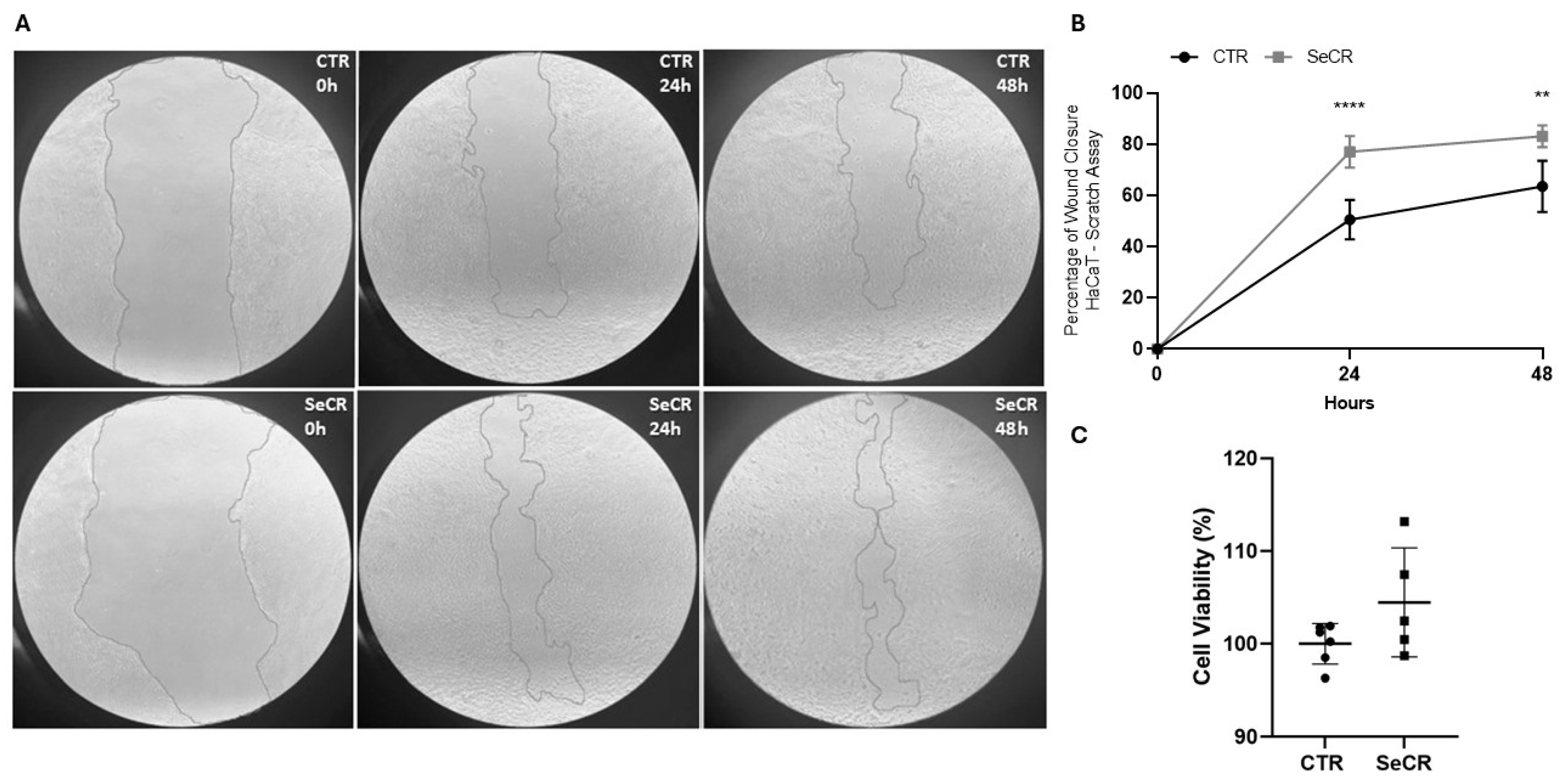
3.2. Secretome Stimulate Cell Migration and Proliferation in Scratch Assay with HaCaT Keratinocytes
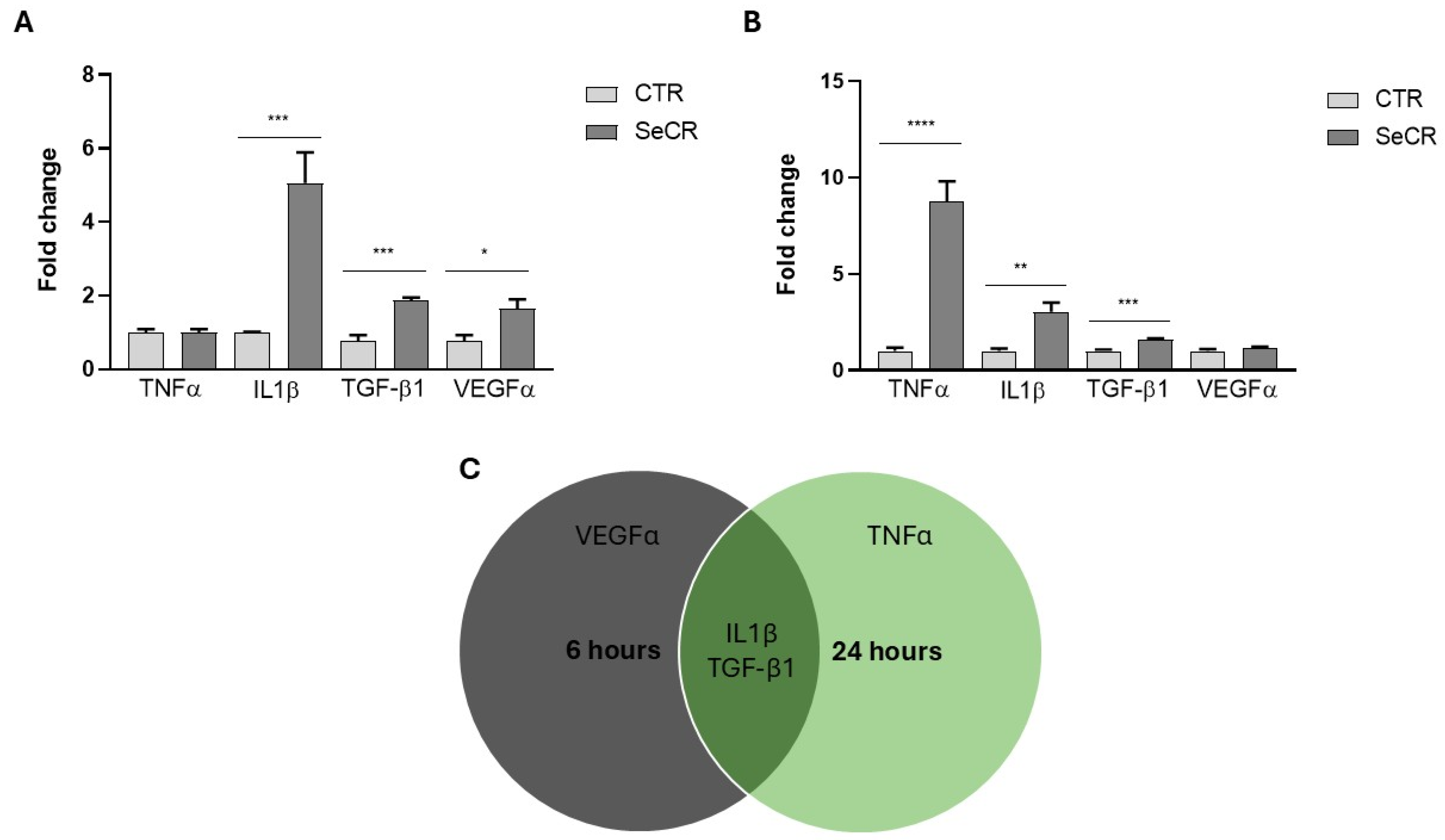
3.3. Expression Profile of Key Inflammatory and Regenerative Genes Induced by the hDP-MSC-Derived Secretome
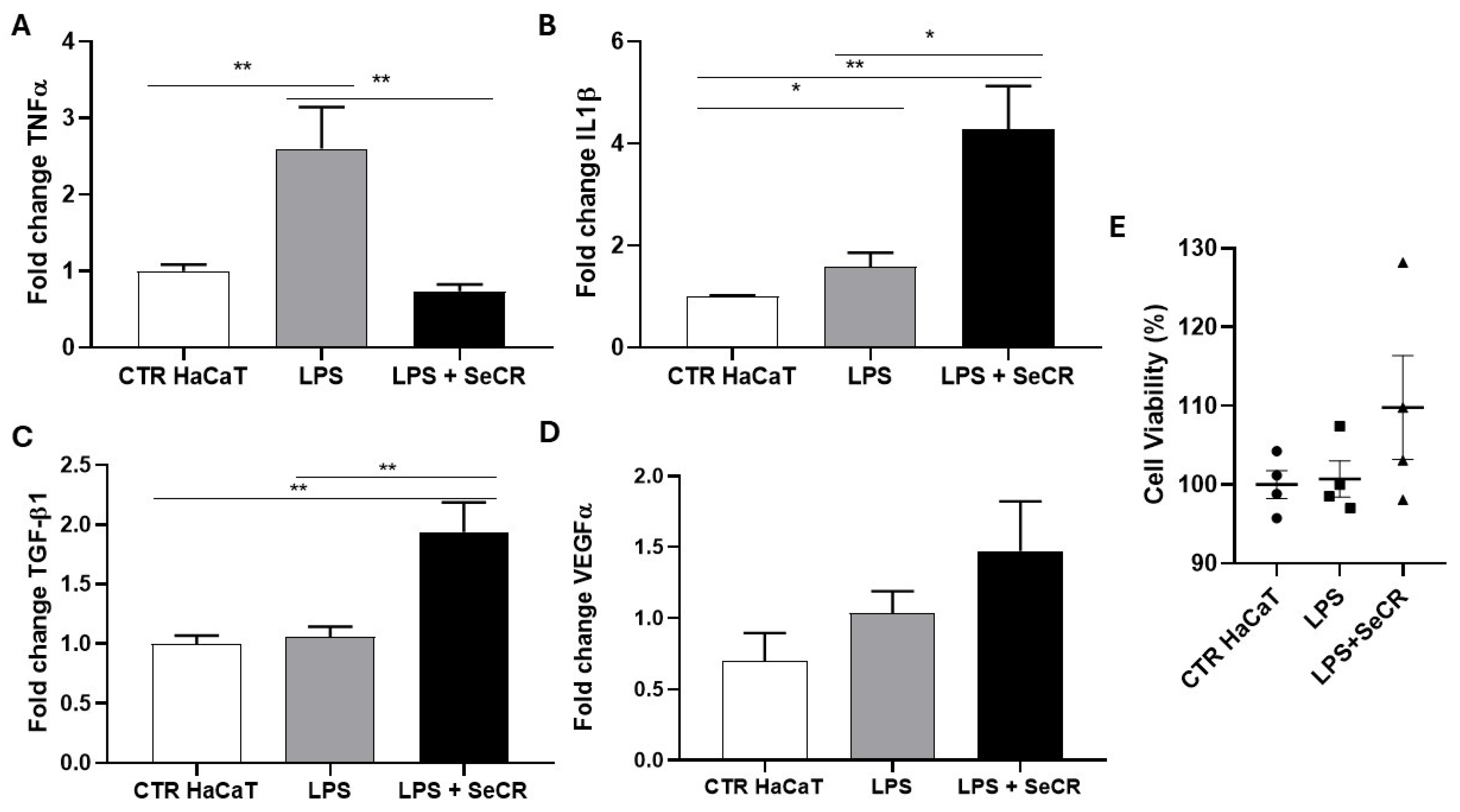
3.4. Effect of the Secretome on the Modulation of the Inflammatory Response in LPS-Stimulated Keratinocytes
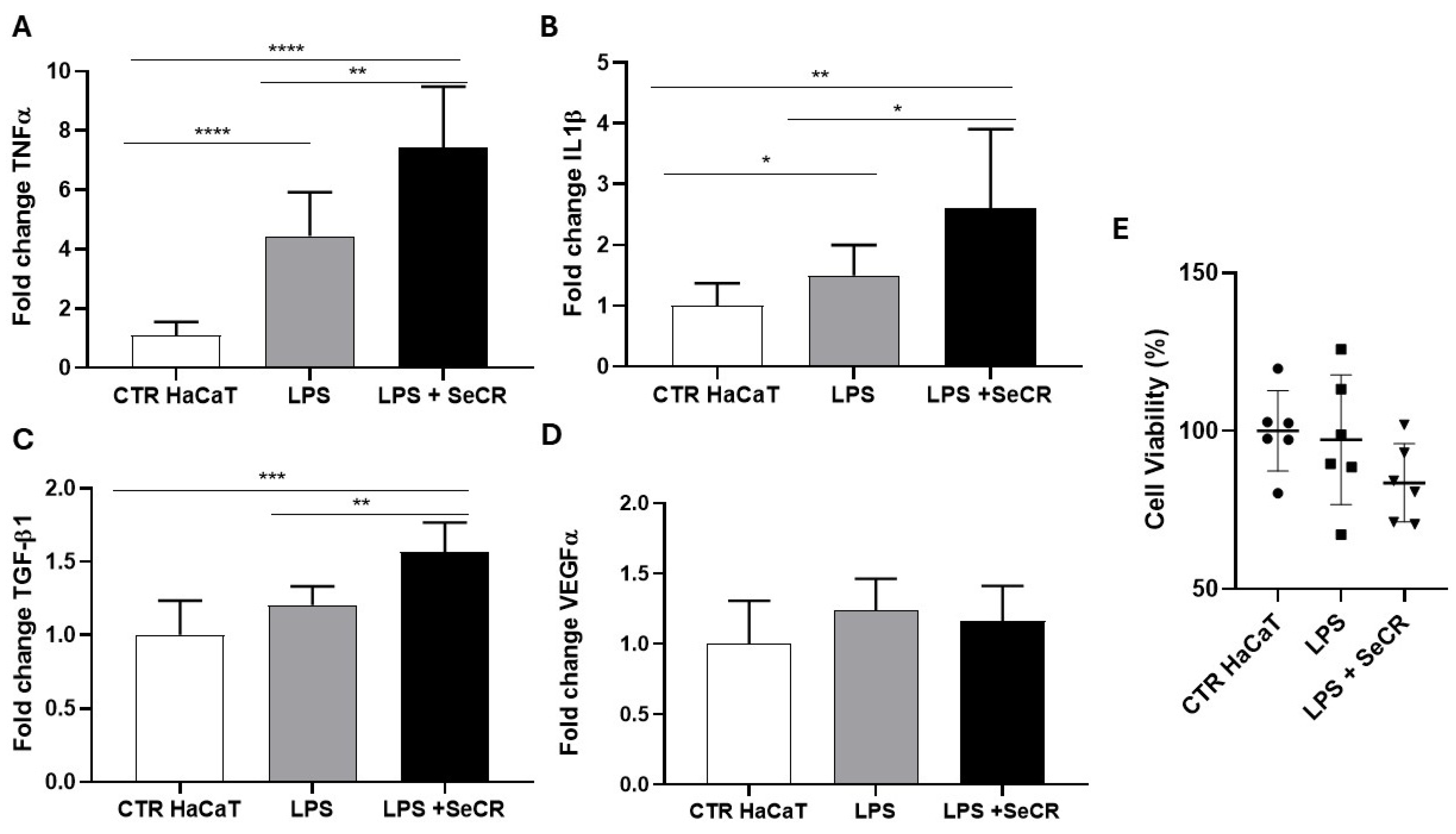
3.5. Secretome-Mediated Modulation After 24 h LPS Exposure
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sorg, H.; Sorg, C.G.G. Skin wound healing: Of players, patterns, and processes. Eur. Surg. Res. 2023, 64, 141–157. [Google Scholar] [CrossRef] [PubMed]
- Martin, P.; Nunan, R. Cellular and molecular mechanisms of repair in acute and chronic wound healing. Br. J. Dermatol. 2015, 173, 370–378. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Burgess, J.L.; Wyant, W.A.; Abujamra, B.A.; Kirsner, R.S.; Jozic, I. Diabetic wound-healing science. Medicina 2021, 57, 1072. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Okonkwo, U.A.; DiPietro, L.A. Diabetes and wound angiogenesis. Int. J. Mol. Sci. 2017, 18, 1419. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Yang, S.; Li, Y.; Liu, C.; Wu, Y.; Wan, Z.; Shen, D. Pathogenesis and treatment of wound healing in patients with diabetes after tooth extraction. Front. Endocrinol. 2022, 13, 949535. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Eming, S.A.; Martin, P.; Tomic-Canic, M. Wound repair and regeneration: Mechanisms, signaling, and translation. Sci. Transl. Med. 2014, 6, 265sr6. [Google Scholar] [CrossRef] [PubMed]
- Landén, N.X.; Li, D.; Ståhle, M. Transition from inflammation to proliferation: A critical step during wound healing. Cell Mol. Life Sci. 2016, 73, 3861–3885. [Google Scholar] [CrossRef] [PubMed]
- Oualla-Bachiri, W.; Fernández-González, A.; Quiñones-Vico, M.I.; Arias-Santiago, S. From grafts to human bioengineered vascularized skin substitutes. Int. J. Mol. Sci. 2020, 21, 8197. [Google Scholar] [CrossRef] [PubMed]
- Editors of Nursing. Pressure ulcers get new terminology and staging definitions. Nursing 2017, 47, 68–69. [Google Scholar]
- Huang, Y.; Wu, Q.; Tam, P.K.H. Immunomodulatory mechanisms of mesenchymal stem cells and their potential clinical applications. Int. J. Mol. Sci. 2022, 23, 10023. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Shi, Y.; Wang, Y.; Li, Q.; Liu, K.; Hou, J.; Shao, C.; Wang, Y. Immunoregulatory mechanisms of mesenchymal stem and stromal cells in inflammatory diseases. Nat. Rev. Nephrol. 2018, 14, 493–507. [Google Scholar] [CrossRef] [PubMed]
- Özdemir, R.B.; Özdemir, A.T.; Kırmaz, C.; Sarıboyacı, A.E.; Karaöz, E.; Erman, G.; Vatansever, H.S.; Gökmen, N.M. Age-related changes in the immunomodulatory effects of human dental pulp derived mesenchymal stem cells on the CD4+ T cell subsets. Cytokine 2021, 138, 155367. [Google Scholar] [CrossRef] [PubMed]
- Yasui, T.; Mabuchi, Y.; Toriumi, H.; Ebine, T.; Niibe, K.; Houlihan, D.D.; Morikawa, S.; Onizawa, K.; Kawana, H.; Akazawa, C.; et al. Purified human dental pulp stem cells promote osteogenic regeneration. J. Dent. Res. 2016, 95, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, T.L.; Cortez de SantAnna, J.P.; Frisene, I.; Gazarini, J.P.; Gomes Pinheiro, C.C.; Gomoll, A.H.; Lattermann, C.; Hernandez, A.J.; Franco Bueno, D. Systematic review of human dental pulp stem cells for cartilage regeneration. Tissue Eng. Part B Rev. 2020, 26, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Thapaswini, Y.; Nikitha Phanindra, S.; Ramavarapu, A.; Kudala, V.R.; Cherukuri, S.A. Evaluation of the toxicity of human dental pulp-derived mesenchymal stem cells on animal models: An animal study. J. Pharm. Bioallied Sci. 2022, 14 (Suppl. S1), S683–S687. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Caseiro, A.R.; Santos Pedrosa, S.; Ivanova, G.; Vieira Branquinho, M.; Almeida, A.; Faria, F.; Amorim, I.; Pereira, T.; Maurício, A.C. Mesenchymal stem/stromal cells metabolomic and bioactive factors profiles: A comparative analysis on the umbilical cord and dental pulp derived stem/stromal cells secretome. PLoS ONE 2019, 14, e0221378. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ivica, A.; Zehnder, M.; Weber, F.E. Therapeutic potential of mesenchymal stem cell-derived extracellular vesicles in regenerative endodontics. Eur. Cell Mater. 2021, 41, 233–244. [Google Scholar] [CrossRef] [PubMed]
- Salkin, H.; Acar, M.B.; Korkmaz, S.; Gunaydin, Z.; Gonen, Z.B.; Basaran, K.E.; Ozcan, S. Transforming growth factor β1-enriched secretome up-regulate osteogenic differentiation of dental pulp stem cells, and a potential therapeutic for gingival wound healing: A comparative proteomics study. J. Dent. 2022, 124, 104224. [Google Scholar] [CrossRef] [PubMed]
- Ranjbar, E.; Tavakol Afshari, J.; KhajaviRad, A.; Ebrahimzadeh-Bideskan, A.E.; Shafieian, R. Insights into the protective capacity of human dental pulp stem cells and its secretome in cisplatin-induced nephrotoxicity: Effects on oxidative stress and histological changes. J. Basic Clin. Physiol. Pharmacol. 2022, 34, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Shekatkar, M.; Kheur, S.; Deshpande, S.; Sanap, A.; Kharat, A.; Navalakha, S.; Gupta, A.; Kheur, M.; Bhonde, R.; Merchant, Y.P. Angiogenic potential of various oral cavity-derived mesenchymal stem cells and cell-derived secretome: A systematic review and meta-analysis. Eur. J. Dent. 2024, 18, 712–742. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Sultan, N.; Amin, A.; Mohammed, F.A. Human exfoliated deciduous teeth stem cells (SHED) display neural crest markers and differentiate into neural-like cells. Saudi Dent. J. 2019, 31, 251–258. [Google Scholar] [CrossRef]
- Naz, S.; Khan, F.R.; Zohra, R.R.; Lakhundi, S.S.; Khan, M.S.; Mohammed, N.; Ahmad, T. Isolation and culture of dental pulp stem cells from permanent and deciduous teeth. Pak. J. Med. Sci. 2019, 35, 997–1002. [Google Scholar] [CrossRef] [PubMed]
- Dominici, M.L.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Krause, D.S.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Mushahary, D.; Spittler, A.; Kasper, C.; Weber, V.; Charwat, V. Isolation, cultivation, and characterization of human mesenchymal stem cells. Cytom. A 2018, 93, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Zanchetta, F.C.; De Wever, P.; Morari, J.; Gaspar, R.C.; Prado, T.P.; De Maeseneer, T.; Cardinaels, R.; Araújo, E.P.; Lima, M.H.; Fardim, P. In vitro and in vivo evaluation of chitosan/HPMC/insulin hydrogel for wound healing applications. Bioengineering 2024, 11, 168. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Nagarajan, A.; Uchil, P.D. Analysis of cell viability by the MTT assay. Cold Spring Harb. Protoc. 2018, 6. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Mai, Z.; Cui, L.; Zhao, X. Engineering exosomes and biomaterial-assisted exosomes as therapeutic carriers for bone regeneration. Stem Cell Res Ther. 2023, 14, 55. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Rodríguez-Fuentes, D.E.; Fernández-Garza, L.E.; Samia-Meza, J.A.; Barrera-Barrera, S.A.; Caplan, A.I.; Barrera-Saldaña, H.A. Mesenchymal stem cells current clinical applications: A systematic review. Arch. Med. Res. 2021, 52, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wei, Q.; Lu, L.; Cui, S.; Ma, K.; Zhang, W.; Ma, F.; Li, H.; Fu, X.; Zhang, C. Immunomodulatory potential of mesenchymal stem cell-derived extracellular vesicles: Targeting immune cells. Front. Immunol. 2023, 14, 1094685. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Meiliana, A.; Dewi, N.M.; Wijaya, A. Mesenchymal stem cell secretome: Cell-free therapeutic strategy in regenerative medicine. Indones Biomed J. 2019, 11, 113–122. [Google Scholar] [CrossRef]
- Marof, F.; Alexandrovna, K.I.; Margiana, R.; Bahramali, M.; Suksatan, W.; Abdelbasset, W.K.; Chupradit, S.; Nasimi, M.; Maashi, M.S. MSCs and their exosomes: A rapidly evolving approach in the context of cutaneous wounds therapy. Stem Cell Res. Ther. 2021, 12, 597. [Google Scholar] [CrossRef] [PubMed]
- Al-Masawa, M.E.; Alshawsh, M.A.; Ng, C.Y.; Ng, A.M.H.; Foo, J.B.; Vijakumaran, U.; Subramaniam, R.; Abdul Ghani, N.A.; Witwer, K.W.; Law, J.X. Efficacy and safety of small extracellular vesicle interventions in wound healing and skin regeneration: A systematic review and meta-analysis of animal studies. Theranostics 2022, 12, 6455–6508. [Google Scholar] [CrossRef] [PubMed]
- Hermann, M.; Peddi, A.; Gerhards, A.; Schmid, R.; Schmitz, D.; Arkudas, A.; Weisbach, V.; Horch, R.E.; Kengelbach-Weigand, A. Secretome of adipose-derived stem cells cultured in platelet lysate improves migration and viability of keratinocytes. Int. J. Mol. Sci. 2023, 24, 3522. [Google Scholar] [CrossRef] [PubMed]
- Spravchikov, N.; Sizyakov, G.; Gartsbein, M.; Accili, D.; Tennenbaum, T.; Wertheimer, E. Glucose effects on skin keratinocytes: Implications for diabetes skin complications. Diabetes 2001, 50, 1627–1635. [Google Scholar] [CrossRef] [PubMed]
- Terashi, H.; Izumi, K.; Deveci, M.; Rhodes, L.M.; Marcelo, C.L. High glucose inhibits human epidermal keratinocyte proliferation for cellular studies on diabetes mellitus. Int. Wound J. 2005, 2, 298–304. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yang, P.; Liu, D.; Gao, M.; Wang, J.; Yu, T.; Zhang, X.; Liu, Y. Inhibiting hyper-O-GlcNAcylation of c-Myc accelerates diabetic wound healing by alleviating keratinocyte dysfunction. Burns Trauma 2021, 9, tkab031. [Google Scholar] [CrossRef] [PubMed]
- Fang, P.H.; Lai, Y.Y.; Chen, C.L.; Wang, H.Y.; Chang, Y.N.; Lin, Y.C.; Yan, Y.T.; Lai, C.H.; Cheng, B. Cobalt protoporphyrin promotes human keratinocyte migration under hyperglycemic conditions. Mol. Med. 2022, 28, 71. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, N.N.; Hamad, K.; Al Shibitini, A.; Juma, S.; Sharifi, S.; Gould, L.; Mahmoudi, M. Investigating inflammatory markers in wound healing: Understanding implications and identifying artifacts. ACS Pharmacol. Transl. Sci. 2024, 7, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Wynn, T.A.; Ramalingam, T.R. Mechanisms of fibrosis: Therapeutic translation for fibrotic disease. Nat. Med. 2012, 18, 1028–1040. [Google Scholar] [CrossRef] [PubMed]
- An, Y.H.; Kim, D.H.; Lee, E.J.; Lee, D.; Park, M.J.; Ko, J.; Kim, D.W.; Koh, J.; Hong, H.S.; Son, Y.; et al. High-efficient production of adipose-derived stem cell (ADSC) secretome through maturation process and its non-scarring wound healing applications. Front. Bioeng. Biotechnol. 2021, 9, 681501. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Yi, X.; Liang, T.; Jiang, S.; He, R.; Hu, Y.; Bai, L.; Wang, C.; Wang, K.; Zhu, L. Autograft microskin combined with adipose-derived stem cell enhances wound healing in a full-thickness skin defect mouse model. Stem Cell Res. Ther. 2019, 10, 279. [Google Scholar] [CrossRef] [PubMed]
- Cuesta-Gomez, N.; Medina-Ruiz, L.; Graham, G.J.; Campbell, J.D. IL-6 and TGF-β-secreting adoptively-transferred murine mesenchymal stromal cells accelerate healing of psoriasis-like skin inflammation and upregulate IL-17A and TGF-β. Int. J. Mol. Sci. 2023, 24, 10132. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Payão, T.S.; Pellegrini, V.; Morari, J.; Gonçalves, G.M.S.; de Godoy, M.C.X.; Gambero, A.; Reis, L.O.; Velloso, L.A.; Araújo, E.P.; Pascoal, L.B. The Secretome of Human Deciduous Tooth-Derived Mesenchymal Stem Cells Enhances In Vitro Wound Healing and Modulates Inflammation. Pharmaceutics 2025, 17, 961. https://doi.org/10.3390/pharmaceutics17080961
Payão TS, Pellegrini V, Morari J, Gonçalves GMS, de Godoy MCX, Gambero A, Reis LO, Velloso LA, Araújo EP, Pascoal LB. The Secretome of Human Deciduous Tooth-Derived Mesenchymal Stem Cells Enhances In Vitro Wound Healing and Modulates Inflammation. Pharmaceutics. 2025; 17(8):961. https://doi.org/10.3390/pharmaceutics17080961
Chicago/Turabian StylePayão, Thais Simião, Vanessa Pellegrini, Joseane Morari, Gisele Mara Silva Gonçalves, Maria Carolina Ximenes de Godoy, Alessandra Gambero, Leonardo O. Reis, Lício Augusto Velloso, Eliana Pereira Araújo, and Lívia Bitencourt Pascoal. 2025. "The Secretome of Human Deciduous Tooth-Derived Mesenchymal Stem Cells Enhances In Vitro Wound Healing and Modulates Inflammation" Pharmaceutics 17, no. 8: 961. https://doi.org/10.3390/pharmaceutics17080961
APA StylePayão, T. S., Pellegrini, V., Morari, J., Gonçalves, G. M. S., de Godoy, M. C. X., Gambero, A., Reis, L. O., Velloso, L. A., Araújo, E. P., & Pascoal, L. B. (2025). The Secretome of Human Deciduous Tooth-Derived Mesenchymal Stem Cells Enhances In Vitro Wound Healing and Modulates Inflammation. Pharmaceutics, 17(8), 961. https://doi.org/10.3390/pharmaceutics17080961