
Oxyresveratrol Attenuates Inflammation in Human Keratinocyte via Regulating NF-kB Signaling and Ameliorates Eczematous Lesion in DNCB-Induced Dermatitis Mice

 , ,
, , 
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Culture of HaCaT Cells and Primary Keratinocyte HEKa Cells
2.2. Reagents
2.2.1. Oxyresveratrol (ORV)
2.2.2. Peptidoglycan (PGN) and Lipopolysaccharides (LPS)
2.3. Cell Viability Assay
2.4. Apoptosis Assay
2.5. Cell Cycle Analysis
2.6. RNA Isolation, Complementary DNA Synthesis, PCR and Quantitative Real Time-PCR Analysis
2.7. IL-6 and IL-8 Cytokine Measurement
2.8. Western Blot Analysis
2.9. Dermatitis Animal Model
2.10. Evaluation of Severity Using Clinical Skin Score
2.11. Pathology Observation and Immunohistochemistry
2.12. Statistical Analysis
3. Results
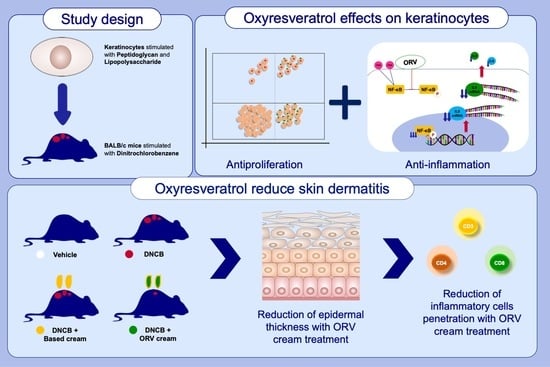
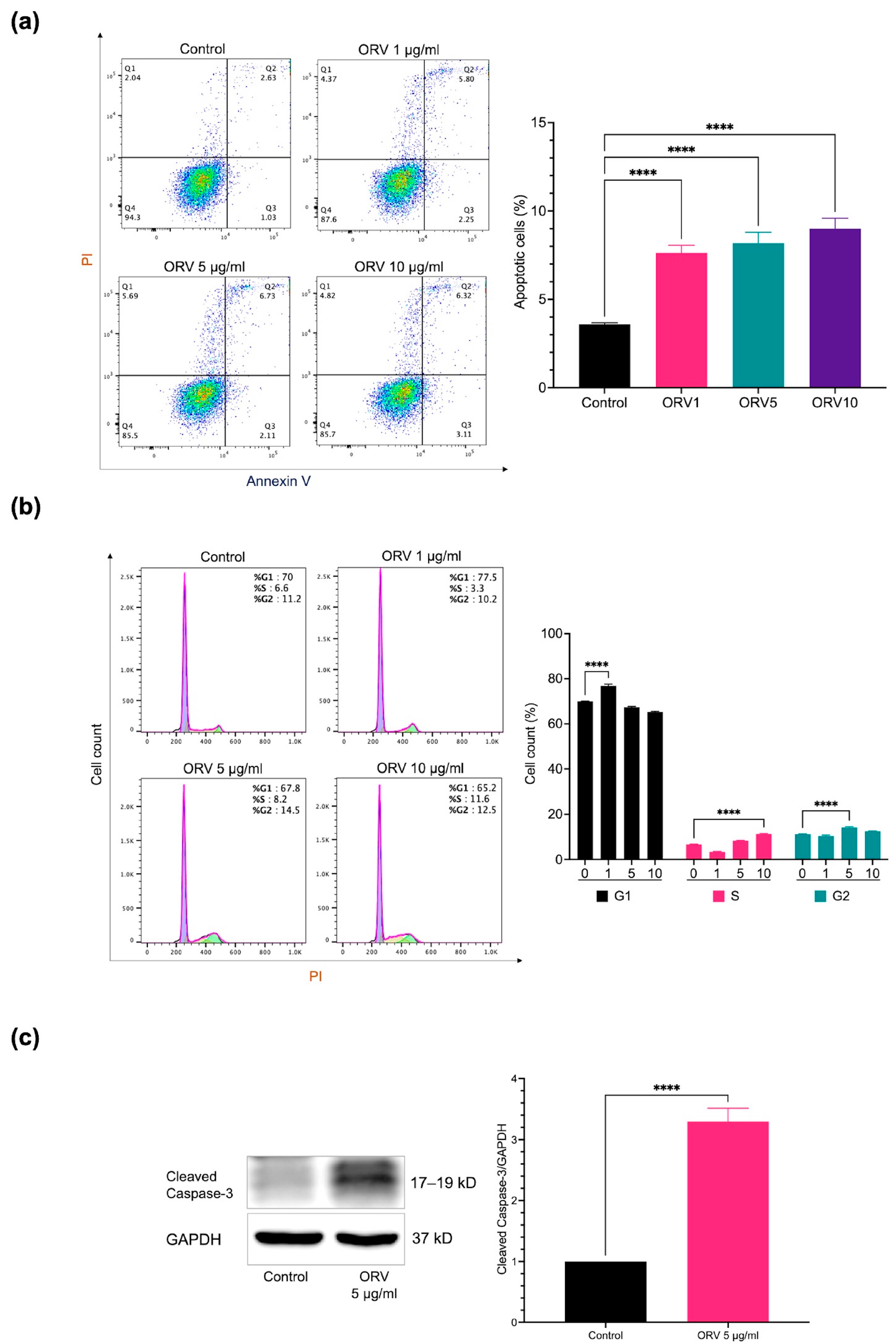
3.1. Effects of Oxyresveratrol on Human Immortalized (HaCaT) and Primary (HEKa) Keratinocytes
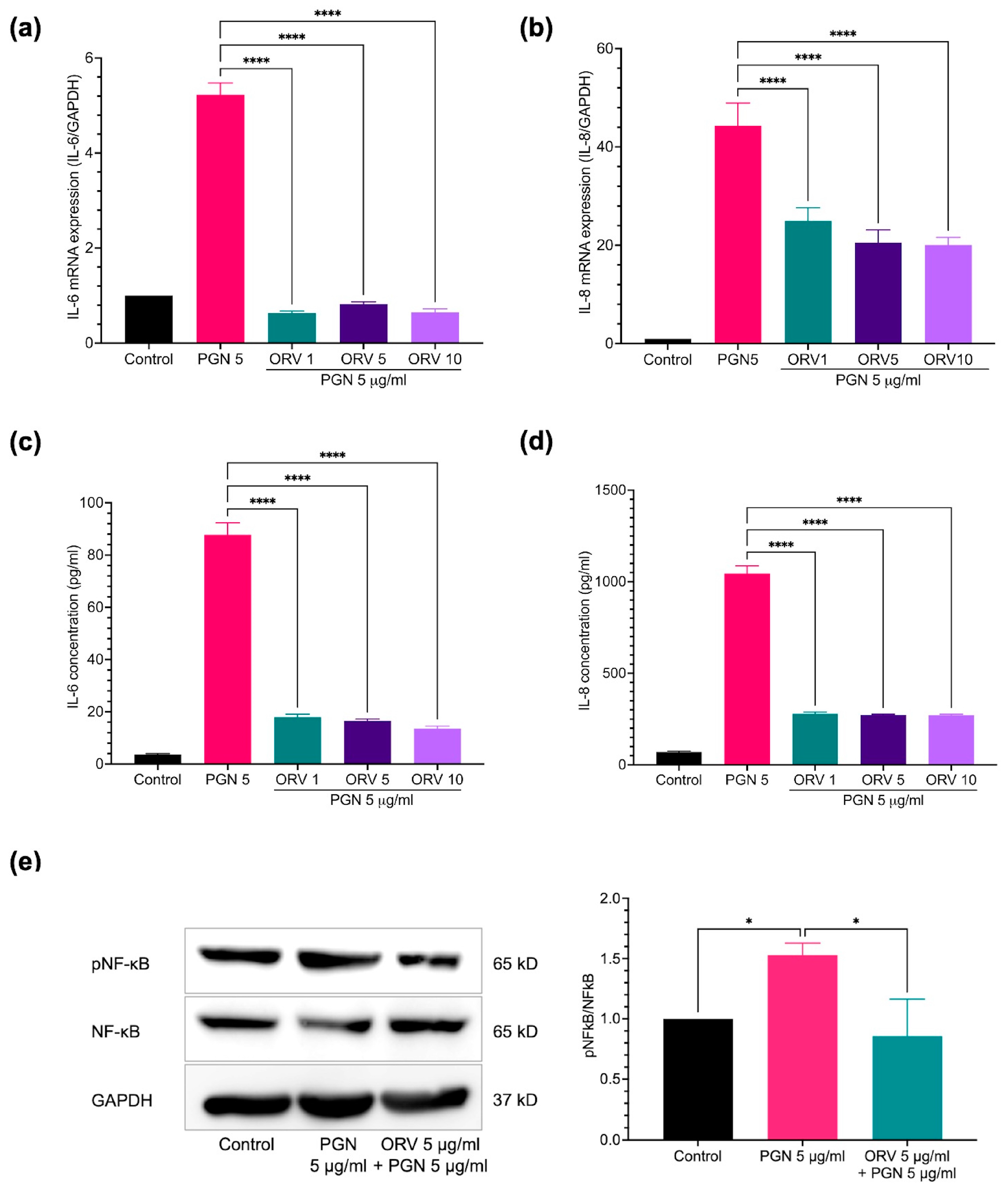
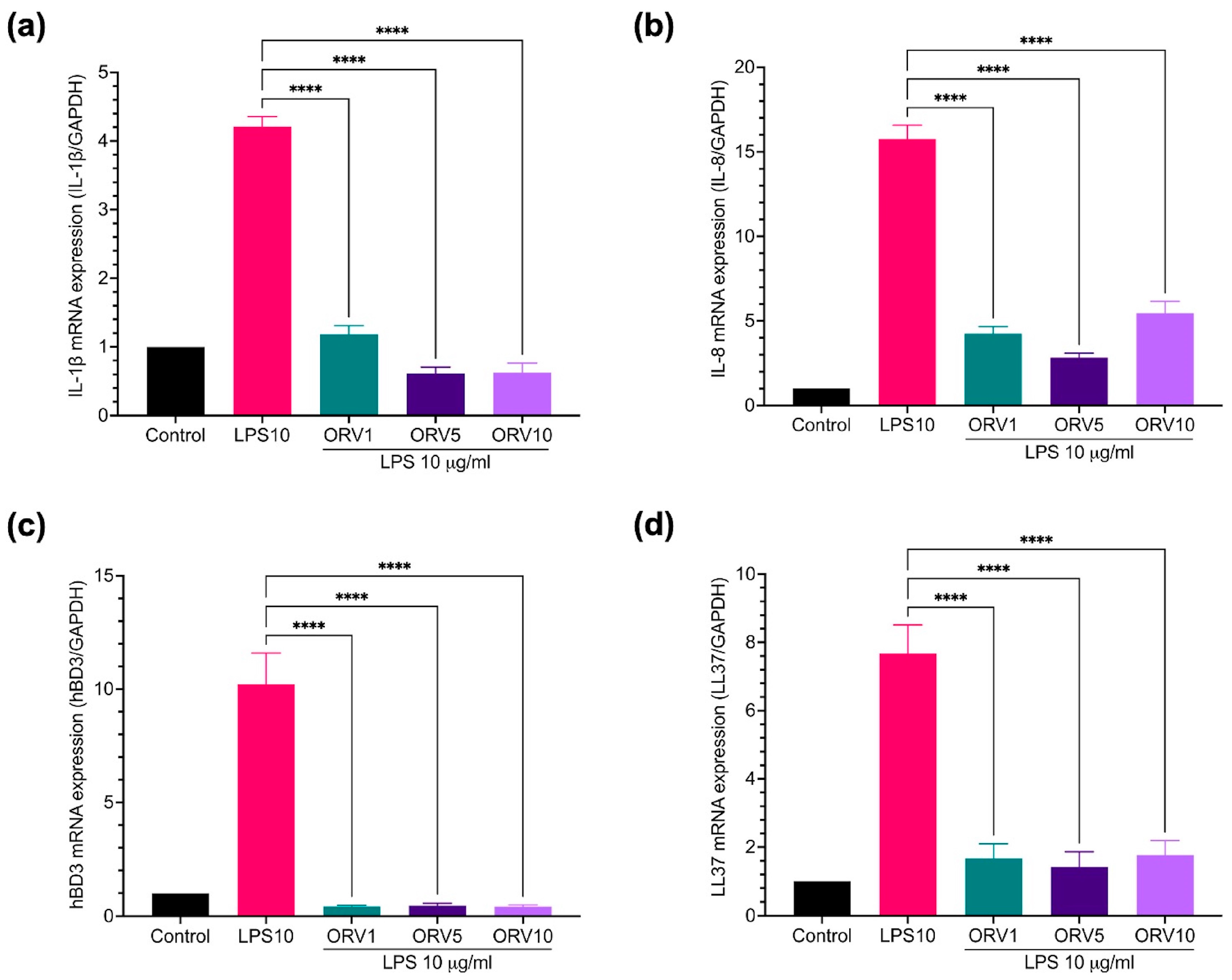
3.2. Anti-Inflammatory Effect of Oxyresveratrol on HaCaT Cells
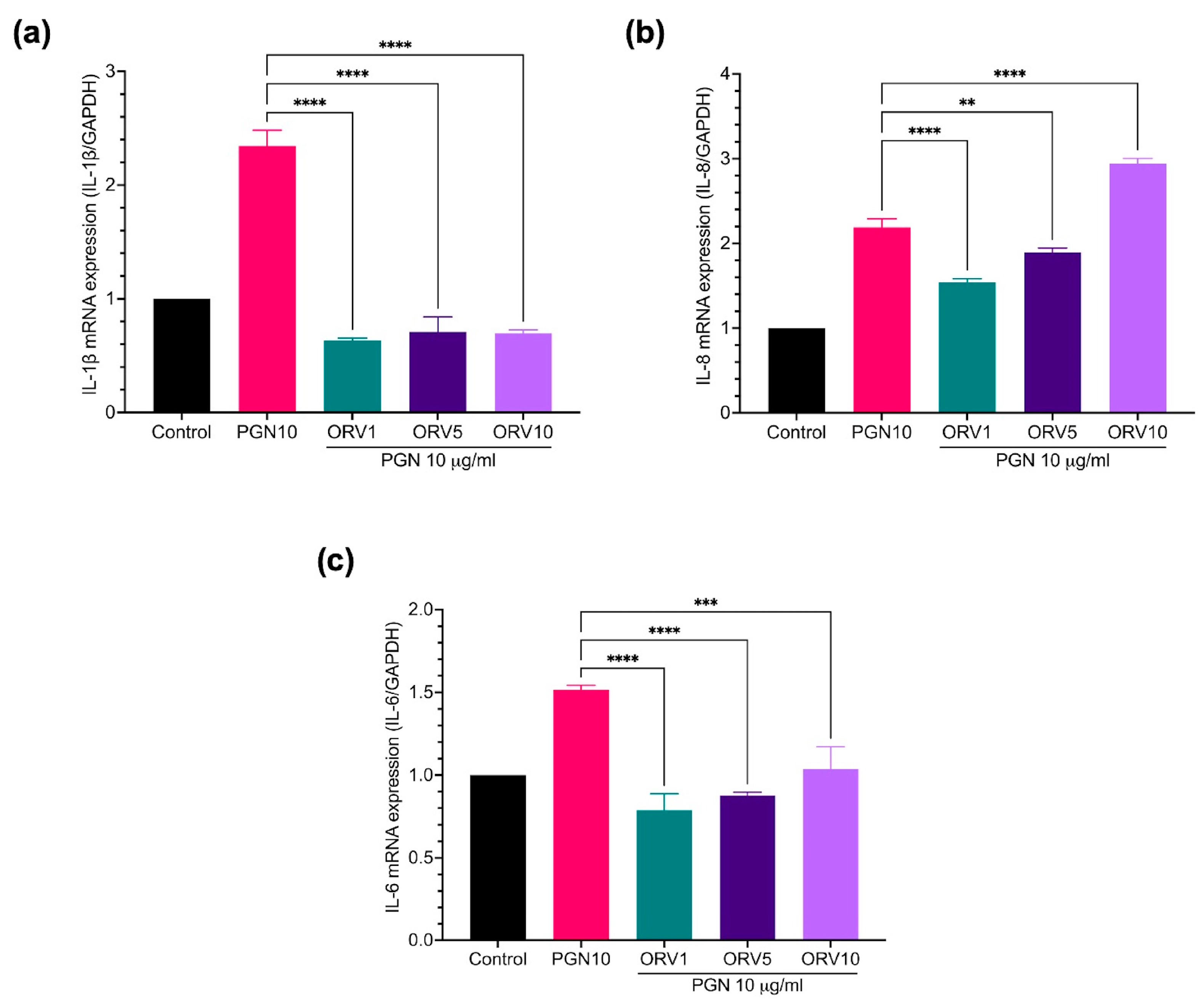
3.3. Anti-Inflammatory Effect of Oxyresveratrol on Primary Epidermal Keratinocytes
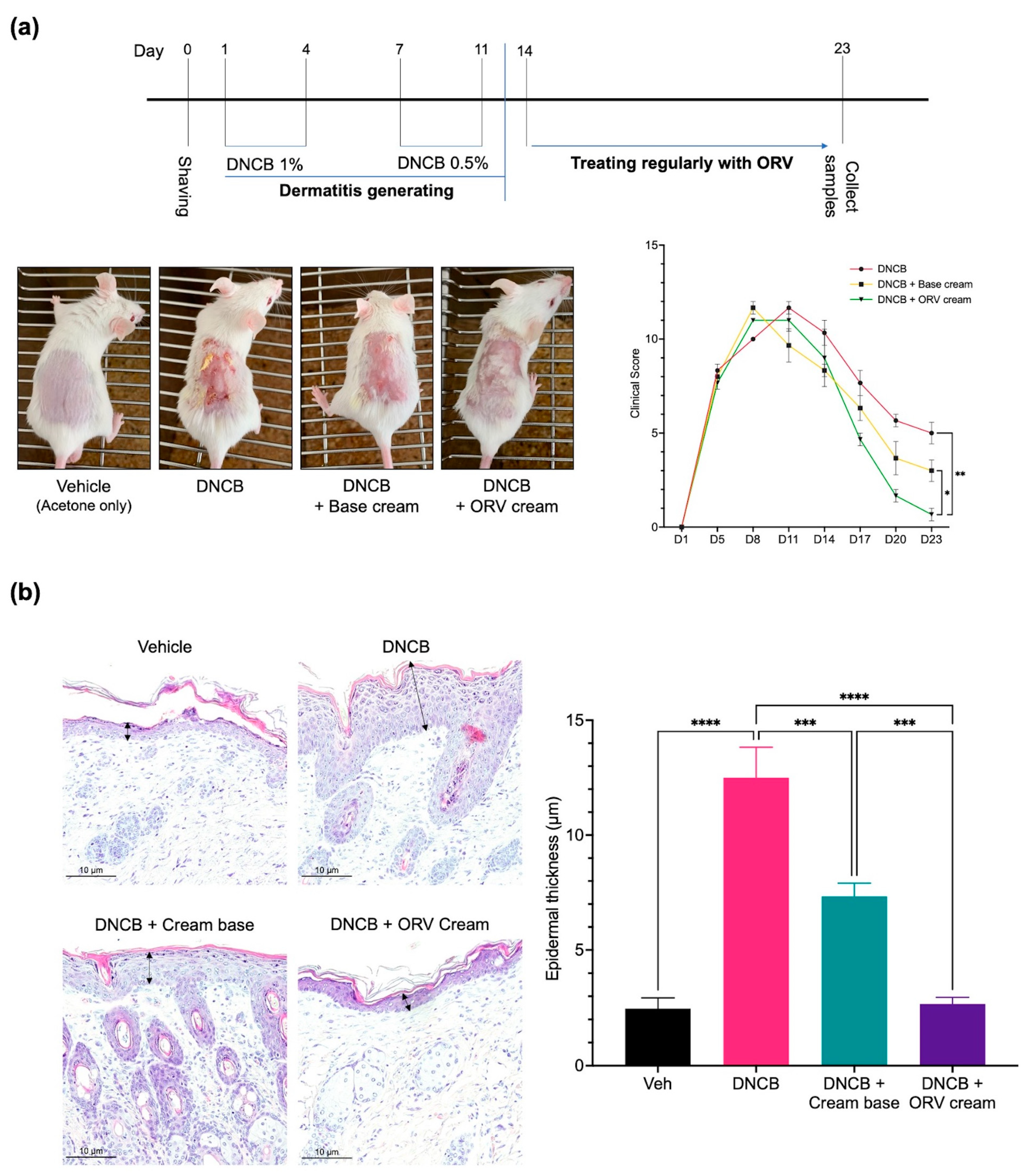
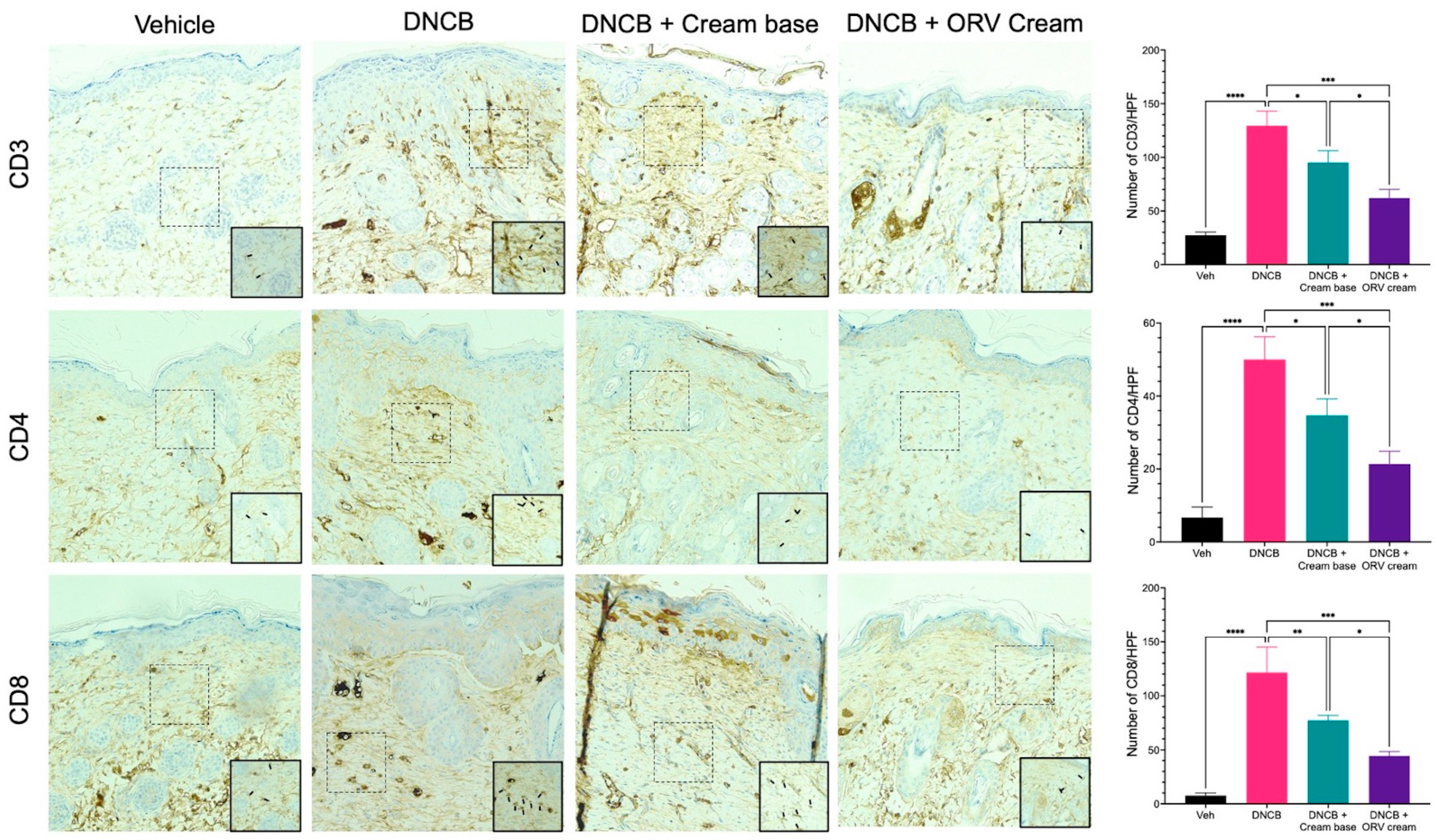
3.4. Oxyresveratrol Treatment Alleviates Skin Inflammatory Manifestation on Mice
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dhakar, N.K.; Matencio, A.; Caldera, F.; Argenziano, M.; Cavalli, R.; Dianzani, C.; Zanetti, M.; López-Nicolás, J.M.; Trotta, F. Comparative Evaluation of Solubility, Cytotoxicity and Photostability Studies of Resveratrol and Oxyresveratrol Loaded Nanosponges. Pharmaceutics 2019, 11, 545. [Google Scholar] [CrossRef]
- Mei, M.; Ruan, J.-Q.; Wu, W.-J.; Zhou, R.-N.; Lei, J.P.-C.; Zhao, H.-Y.; Yan, R.; Wang, Y.-T. In Vitro Pharmacokinetic Characterization of Mulberroside A, the Main Polyhydroxylated Stilbene in Mulberry (Morus alba L.), and Its Bacterial Metabolite Oxyresveratrol in Traditional Oral Use. J. Agric. Food Chem. 2012, 60, 2299–2308. [Google Scholar] [CrossRef] [PubMed]
- Chatsumpun, N.; Chuanasa, T.; Sritularak, B.; Lipipun, V.; Jongbunprasert, V.; Ruchirawat, S.; Ploypradith, P.; Likhitwitayawuid, K. Oxyresveratrol: Structural Modification and Evaluation of Biological Activities. Molecules 2016, 21, 489. [Google Scholar] [CrossRef]
- Chung, K.O.; Kim, B.Y.; Lee, M.H.; Kim, Y.R.; Chung, H.Y.; Park, J.H.; Moon, J.O. In-vitro and in-vivo anti-inflammatory effect of oxyresveratrol from Morus alba L. J. Pharm. Pharm. 2003, 55, 1695–1700. [Google Scholar] [CrossRef]
- Du, H.; Ma, L.; Chen, G.; Li, S. The effects of oxyresveratrol abrogates inflammation and oxidative stress in rat model of spinal cord injury. Mol. Med. Rep. 2018, 17, 4067–4073. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Kim, D.H.; Hong, J.E.; Lee, J.Y.; Kim, E.J. Oxyresveratrol suppresses lipopolysaccharide-induced inflammatory responses in murine macrophages. Hum. Exp. Toxicol. 2015, 34, 808–818. [Google Scholar] [CrossRef] [PubMed]
- Hankittichai, P.; Lou, H.J.; Wikan, N.; Smith, D.R.; Potikanond, S.; Nimlamool, W. Oxyresveratrol Inhibits IL-1β-Induced Inflammation via Suppressing AKT and ERK1/2 Activation in Human Microglia, HMC3. Int. J. Mol. Sci. 2020, 21, 6054. [Google Scholar] [CrossRef]
- Abdel Bar, F.M.; Abbas, G.M.; Gohar, A.A.; Lahloub, M.-F.I. Antiproliferative activity of stilbene derivatives and other constituents from the stem bark of Morus nigra L. Nat. Prod. Res. 2020, 34, 3506–3513. [Google Scholar] [CrossRef]
- Sintuyanon, N.; Phoolcharoen, W.; Pavasant, P.; Sooampon, S. Resveratrol Demonstrated Higher Antiproliferative and Antiangiogenic Efficacy Compared with Oxyresveratrol on Head and Neck Squamous Cell Carcinoma Cell Lines. Nat. Prod. Commun. 2017, 12, 1934578X1701201134. [Google Scholar] [CrossRef]
- Li, H.; Wu, W.K.K.; Zheng, Z.; Che, C.T.; Li, Z.J.; Xu, D.D.; Wong, C.C.M.; Ye, C.G.; Sung, J.J.Y.; Cho, C.H.; et al. 3,3′,4,5,5′-pentahydroxy-trans-stilbene, a resveratrol derivative, induces apoptosis in colorectal carcinoma cells via oxidative stress. Eur. J. Pharmacol. 2010, 637, 55–61. [Google Scholar] [CrossRef]
- Simpson, E.L.; Leung, D.Y.; Eichenfield, L.F.; Boguniewicz, M. Atopic Dermatitis. In Fitzpatrick’s Dermatology, 9th ed.; McGraw-Hill Education: New York, NY, USA, 2019; Volume 1, pp. 363–384. [Google Scholar]
- Turrentine, J.E.; Sheehan, M.P.; Cruz, P.D. Allergic Contact Dermatitis. In Fitzpatric’s Dermatology, 9th ed.; McGraw-Hill Education: New York, NY, USA, 2019; Volume 1, pp. 395–413. [Google Scholar]
- Karimkhani, C.; Dellavalle, R.P.; Coffeng, L.E.; Flohr, C.; Hay, R.J.; Langan, S.M.; Nsoesie, E.O.; Ferrari, A.J.; Erskine, H.E.; Silverberg, J.I.; et al. Global Skin Disease Morbidity and Mortality: An Update from the Global Burden of Disease Study 2013. JAMA Dermatol. 2017, 153, 406–412. [Google Scholar] [CrossRef] [PubMed]
- Hanifin, J.M.; Reed, M.L.; Prevalence, E.; Group, I.W. A Population-Based Survey of Eczema Prevalence in the United States. Dermatitis 2007, 18, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Danso, M.O.; van Drongelen, V.; Mulder, A.; van Esch, J.; Scott, H.; van Smeden, J.; El Ghalbzouri, A.; Bouwstra, J.A. TNF-α and Th2 cytokines induce atopic dermatitis-like features on epidermal differentiation proteins and stratum corneum lipids in human skin equivalents. J. Investig. Derm. 2014, 134, 1941–1950. [Google Scholar] [CrossRef] [PubMed]
- Vocanson, M.; Hennino, A.; Rozières, A.; Poyet, G.; Nicolas, J.F. Effector and regulatory mechanisms in allergic contact dermatitis. Allergy 2009, 64, 1699–1714. [Google Scholar] [CrossRef]
- Lee, H.Y.; Stieger, M.; Yawalkar, N.; Kakeda, M. Cytokines and Chemokines in Irritant Contact Dermatitis. Mediat. Inflamm. 2013, 2013, 916497. [Google Scholar] [CrossRef]
- Gabros, S.; Nessel, T.A.; Zito, P.M. Topical Corticosteroids. In StatPearls; StatPearls Publishing LLC.: Treasure Island, FL, USA, 2022. [Google Scholar]
- Gutfreund, K.; Bienias, W.; Szewczyk, A.; Kaszuba, A. Topical calcineurin inhibitors in dermatology. Part I: Properties, method and effectiveness of drug use. Postep. Derm. Alergol. 2013, 30, 165–169. [Google Scholar] [CrossRef]
- Wongpiyabovorn, J.; Soonthornchai, W.; Wilantho, A.; Palasuk, M.; Payungporn, S.; Sodsai, P.; Poomipak, W.; Weschawalit, S.; Ruchusatsawat, K.; Baillie, G.S.; et al. Effect of tacrolimus on skin microbiome in atopic dermatitis. Allergy 2019, 74, 1400–1406. [Google Scholar] [CrossRef]
- Russell, G.; Graveley, R.; Seid, J.; al-Humidan, A.K.; Skjodt, H. Mechanisms of action of cyclosporine and effects on connective tissues. Semin. Arthritis Rheum 1992, 21, 16–22. [Google Scholar] [CrossRef]
- Schram, M.E.; Roekevisch, E.; Leeflang, M.M.; Bos, J.D.; Schmitt, J.; Spuls, P.I. A randomized trial of methotrexate versus azathioprine for severe atopic eczema. J. Allergy Clin. Immunol. 2011, 128, 353–359. [Google Scholar] [CrossRef]
- Thaçi, D.; Simpson, E.L.; Beck, L.A.; Bieber, T.; Blauvelt, A.; Papp, K.; Soong, W.; Worm, M.; Szepietowski, J.C.; Sofen, H.; et al. Efficacy and safety of dupilumab in adults with moderate-to-severe atopic dermatitis inadequately controlled by topical treatments: A randomised, placebo-controlled, dose-ranging phase 2b trial. Lancet 2016, 387, 40–52. [Google Scholar] [CrossRef]
- Kim, J.; Kim, B.E.; Leung, D.Y.M. Pathophysiology of atopic dermatitis: Clinical implications. Allergy Asthma Proc. 2019, 40, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Bissonnette, R.; Papp, K.A.; Poulin, Y.; Gooderham, M.; Raman, M.; Mallbris, L.; Wang, C.; Purohit, V.; Mamolo, C.; Papacharalambous, J.; et al. Topical tofacitinib for atopic dermatitis: A phase IIa randomized trial. Br. J. Derm. 2016, 175, 902–911. [Google Scholar] [CrossRef] [PubMed]
- Guttman-Yassky, E.; Silverberg, J.I.; Nemoto, O.; Forman, S.B.; Wilke, A.; Prescilla, R.; de la Peña, A.; Nunes, F.P.; Janes, J.; Gamalo, M.; et al. Baricitinib in adult patients with moderate-to-severe atopic dermatitis: A phase 2 parallel, double-blinded, randomized placebo-controlled multiple-dose study. J. Am. Acad. Derm. 2019, 80, 913–921.e919. [Google Scholar] [CrossRef]
- Hong, S.H.; Ku, J.M.; Kim, H.I.; Kim, T.Y.; Seo, H.S.; Shin, Y.C.; Ko, S.-G. Topical Application of KAJD Attenuates 2,4-Dinitrochlorobenzene-Induced Atopic Dermatitis Symptoms Through Regulation of IgE and MAPK Pathways in BALB/C Mice and Several Immune Cell Types. Front. Pharmacol. 2019, 10, 1097. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.C.; Hung, Y.L.; Ko, W.C.; Tsai, Y.J.; Chang, J.F.; Liang, C.W.; Chang, D.C.; Hung, C.F. Effect of Neferine on DNCB-Induced Atopic Dermatitis in HaCaT Cells and BALB/c Mice. Int. J. Mol. Sci. 2021, 22, 8237. [Google Scholar] [CrossRef] [PubMed]
- Proksch, E.; Fölster-Holst, R.; Jensen, J.M. Skin barrier function, epidermal proliferation and differentiation in eczema. J. Derm. Sci. 2006, 43, 159–169. [Google Scholar] [CrossRef]
- Kerdmuanglek, F.; Chomtong, T.; Boonsith, S.; Chutimasakul, T.; Iemsam-Arng, J.; Thepwatee, S. Non-Ionic surfactant-assisted controlled release of oxyresveratrol on dendritic fibrous silica for topical applications. J. Colloid Interface Sci. 2023, 646, 342–353. [Google Scholar] [CrossRef]
- Wikan, N.; Hankittichai, P.; Thaklaewphan, P.; Potikanond, S.; Nimlamool, W. Oxyresveratrol Inhibits TNF-α-Stimulated Cell Proliferation in Human Immortalized Keratinocytes (HaCaT) by Suppressing AKT Activation. Pharmaceutics 2022, 14, 63. [Google Scholar]
- Barnum, K.J.; O’Connell, M.J. Cell cycle regulation by checkpoints. Methods Mol. Biol. 2014, 1170, 29–40. [Google Scholar] [CrossRef]
- King, K.L.; Cidlowski, J.A. CELL CYCLE REGULATION AND APOPTOSIS. Annu. Rev. Physiol. 1998, 60, 601–617. [Google Scholar] [CrossRef]
- Seo, M.D.; Kang, T.J.; Lee, C.H.; Lee, A.Y.; Noh, M. HaCaT Keratinocytes and Primary Epidermal Keratinocytes Have Different Transcriptional Profiles of Cornified Envelope-Associated Genes to T Helper Cell Cytokines. Biomol. Ther. (Seoul) 2012, 20, 171–176. [Google Scholar] [CrossRef]
- Moran, M.C.; Pandya, R.P.; Leffler, K.A.; Yoshida, T.; Beck, L.A.; Brewer, M.G. Characterization of Human Keratinocyte Cell Lines for Barrier Studies. JID Innov. 2021, 1, 100018. [Google Scholar] [CrossRef] [PubMed]
- Juráňová, J.; Franková, J.; Ulrichová, J. The role of keratinocytes in inflammation. J. Appl. Biomed. 2017, 15, 169–179. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-κB signaling in inflammation. Signal Transduct. Target Ther. 2017, 2, 17023. [Google Scholar] [CrossRef]
- Yamasaki, K.; Gallo, R.L. Antimicrobial peptides in human skin disease. Eur. J. Derm. 2008, 18, 11–21. [Google Scholar] [CrossRef]
- Herman, A.; Herman, A.P. Antimicrobial peptides activity in the skin. Ski. Res. Technol. 2019, 25, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Frohm, M.; Agerberth, B.; Ahangari, G.; Stâhle-Bäckdahl, M.; Lidén, S.; Wigzell, H.; Gudmundsson, G.H. The expression of the gene coding for the antibacterial peptide LL-37 is induced in human keratinocytes during inflammatory disorders. J. Biol. Chem. 1997, 272, 15258–15263. [Google Scholar] [CrossRef]
- Niyonsaba, F.; Ushio, H.; Nakano, N.; Ng, W.; Sayama, K.; Hashimoto, K.; Nagaoka, I.; Okumura, K.; Ogawa, H. Antimicrobial peptides human beta-defensins stimulate epidermal keratinocyte migration, proliferation and production of proinflammatory cytokines and chemokines. J. Investig. Derm. 2007, 127, 594–604. [Google Scholar] [CrossRef]
- Johnston, G.A.; Exton, L.S.; Mohd Mustapa, M.F.; Slack, J.A.; Coulson, I.H.; English, J.S.C.; Bourke, J.F. British Association of Dermatologists’ guidelines for the management of contact dermatitis 2017. Br. J. Dermatol. 2017, 176, 317–329. [Google Scholar] [CrossRef]
- Yang, H.; Parkhouse, R.M.; Wileman, T. Monoclonal antibodies that identify the CD3 molecules expressed specifically at the surface of porcine gammadelta-T cells. Immunology 2005, 115, 189–196. [Google Scholar] [CrossRef]
- Hijnen, D.; Knol, E.F.; Gent, Y.Y.; Giovannone, B.; Beijn, S.J.; Kupper, T.S.; Bruijnzeel-Koomen, C.A.; Clark, R.A. CD8(+) T cells in the lesional skin of atopic dermatitis and psoriasis patients are an important source of IFN-γ, IL-13, IL-17, and IL-22. J. Investig. Derm. 2013, 133, 973–979. [Google Scholar] [CrossRef] [PubMed]
- James, E.A.; Kwok, W.W. Autoreactive CD4(+) T cells in patients with atopic dermatitis. J. Allergy Clin. Immunol. 2011, 128, 100–101. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tran, H.G.; Shuayprom, A.; Kueanjinda, P.; Leelahavanichkul, A.; Wongsinkongman, P.; Chaisomboonpan, S.; Tawatsin, A.; Ruchusatsawat, K.; Wongpiyabovorn, J. Oxyresveratrol Attenuates Inflammation in Human Keratinocyte via Regulating NF-kB Signaling and Ameliorates Eczematous Lesion in DNCB-Induced Dermatitis Mice. Pharmaceutics 2023, 15, 1709. https://doi.org/10.3390/pharmaceutics15061709
Tran HG, Shuayprom A, Kueanjinda P, Leelahavanichkul A, Wongsinkongman P, Chaisomboonpan S, Tawatsin A, Ruchusatsawat K, Wongpiyabovorn J. Oxyresveratrol Attenuates Inflammation in Human Keratinocyte via Regulating NF-kB Signaling and Ameliorates Eczematous Lesion in DNCB-Induced Dermatitis Mice. Pharmaceutics. 2023; 15(6):1709. https://doi.org/10.3390/pharmaceutics15061709
Chicago/Turabian StyleTran, Hung Gia, Aussavashai Shuayprom, Patipark Kueanjinda, Asada Leelahavanichkul, Prapai Wongsinkongman, Siriwan Chaisomboonpan, Apiwat Tawatsin, Kriangsak Ruchusatsawat, and Jongkonnee Wongpiyabovorn. 2023. "Oxyresveratrol Attenuates Inflammation in Human Keratinocyte via Regulating NF-kB Signaling and Ameliorates Eczematous Lesion in DNCB-Induced Dermatitis Mice" Pharmaceutics 15, no. 6: 1709. https://doi.org/10.3390/pharmaceutics15061709
APA StyleTran, H. G., Shuayprom, A., Kueanjinda, P., Leelahavanichkul, A., Wongsinkongman, P., Chaisomboonpan, S., Tawatsin, A., Ruchusatsawat, K., & Wongpiyabovorn, J. (2023). Oxyresveratrol Attenuates Inflammation in Human Keratinocyte via Regulating NF-kB Signaling and Ameliorates Eczematous Lesion in DNCB-Induced Dermatitis Mice. Pharmaceutics, 15(6), 1709. https://doi.org/10.3390/pharmaceutics15061709