

Comprehensive Study of Antiretroviral Drug Permeability at the Cervicovaginal Mucosa via an In Vitro Model



 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines, Growth Media, Drugs and Cell Stimulants
2.2. Transwells®
2.3. Measurement of Transepithelial Electrical Resistance (TEER)
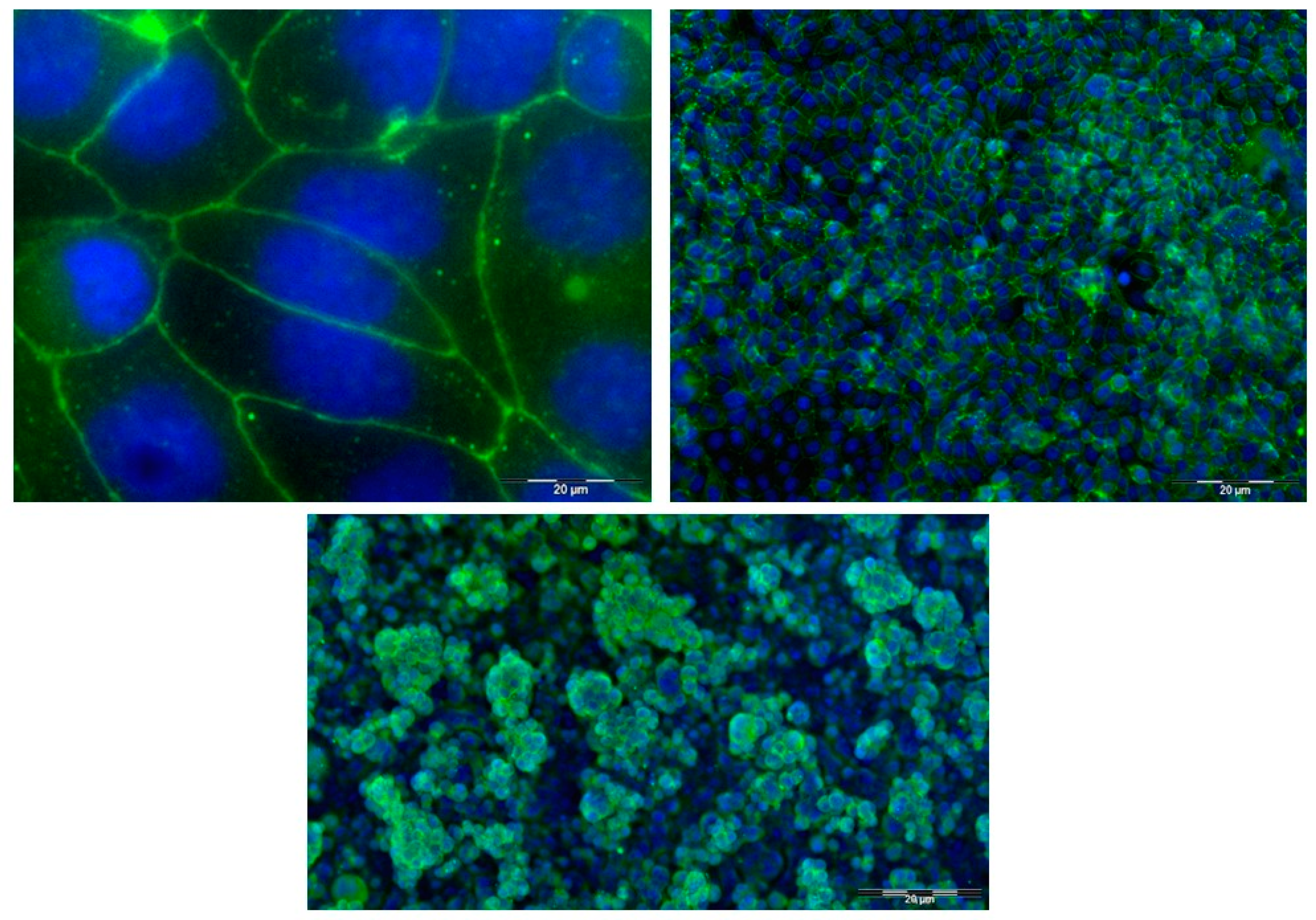
2.4. Immunofluorescence
2.5. Drug Permeability Studies
2.6. Intracellular Accumulation Studies
2.7. LC-MS/MS
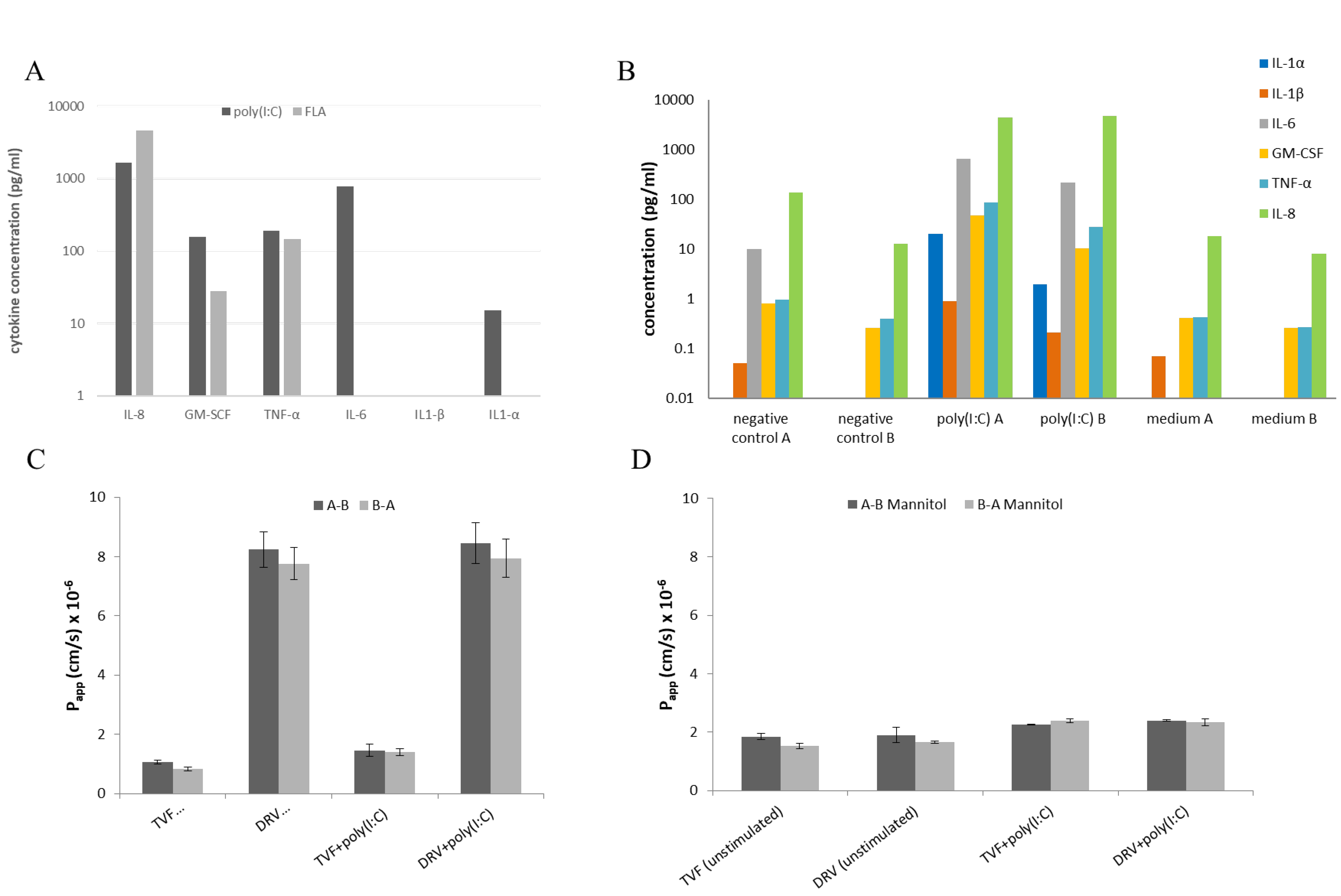
2.8. Stimulation with Pathogen-Associated Molecular Patterns (PAMP) and Cytokine Detection
2.9. Cell line Stimulation with Drugs, Cytokines, Hormones and Microbes
2.10. Extraction of RNA, cDNA Synthesis and Real-Time Quantitative PCR
2.11. Statistical Analysis
3. Results
3.1. Dual Chamber Cell Culture Model of Barrier Epithelium
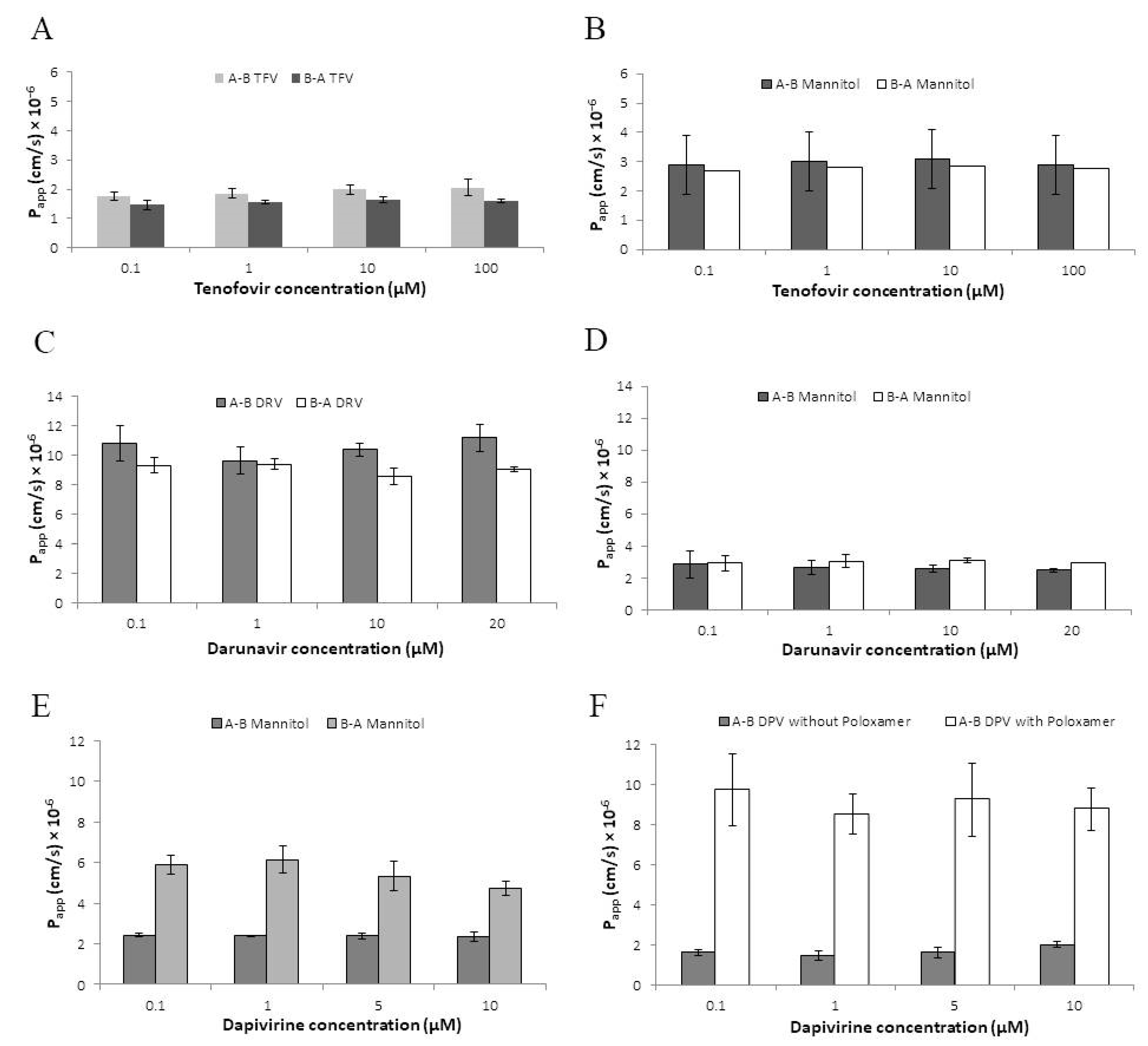
3.2. Permeability of Antiretroviral Drugs across the HEC-1A Model Epithelium
Coadministrations of ARV Drugs
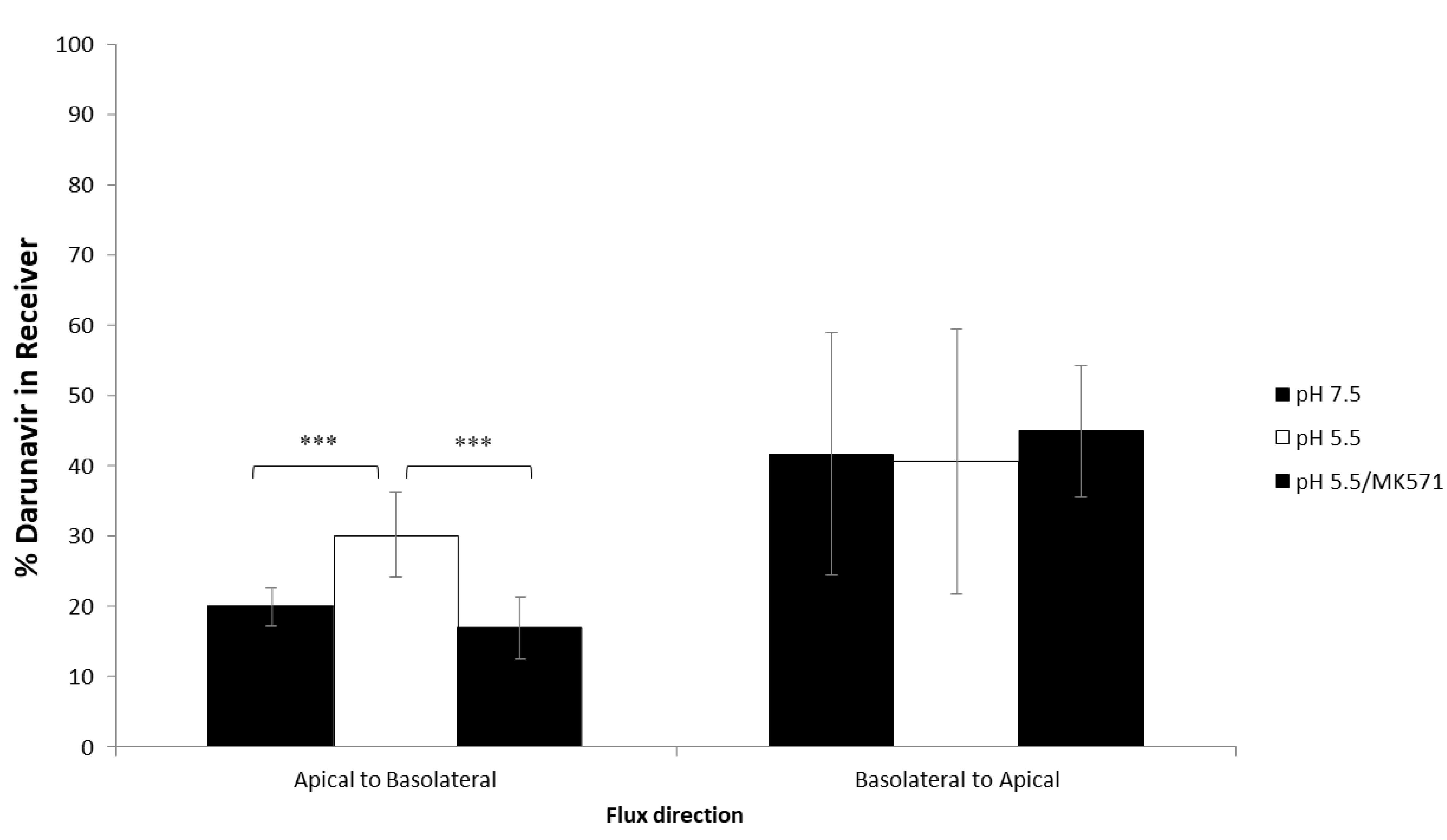
3.3. Effect of pH on Darunavir Permeability
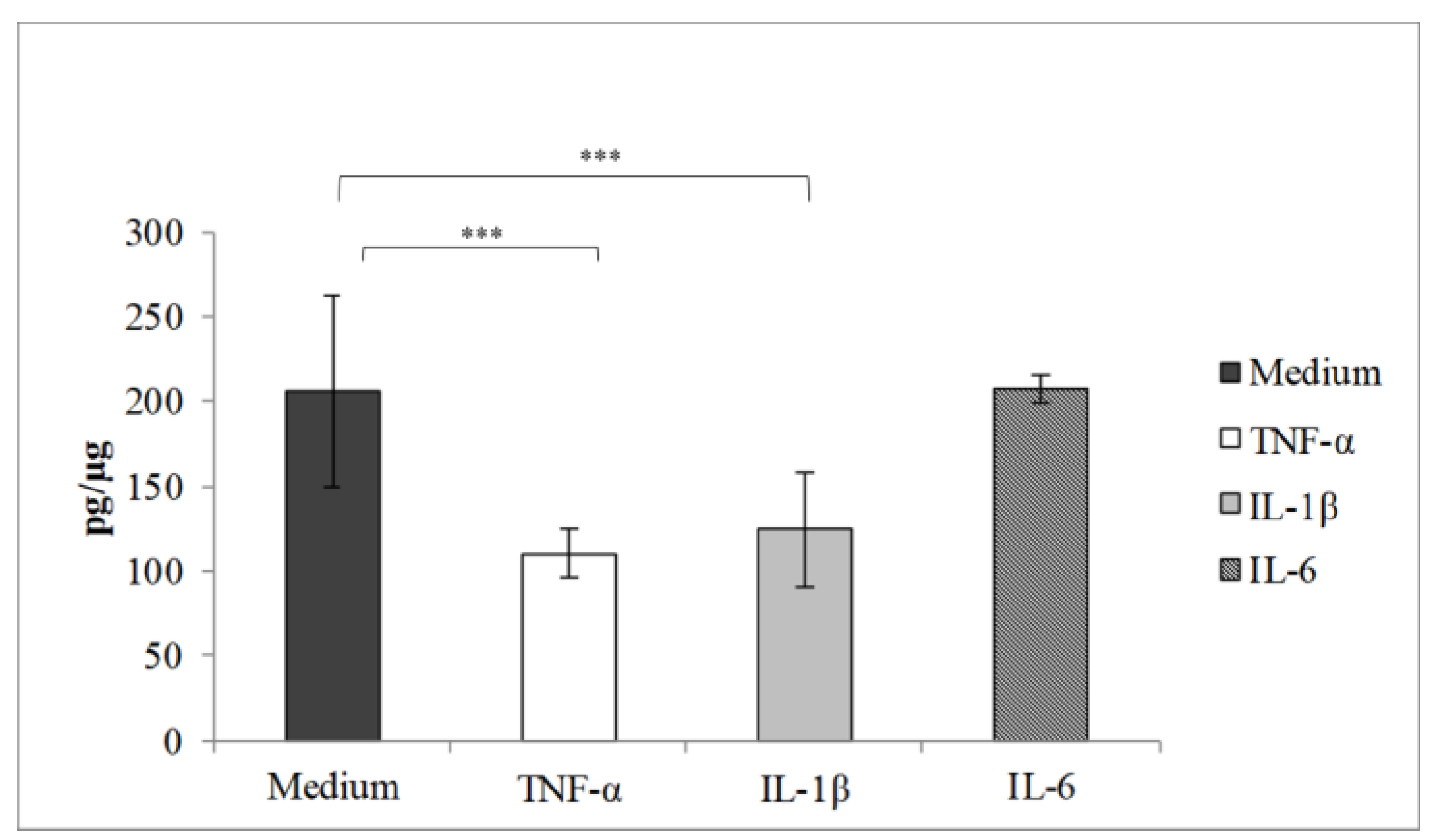
3.4. Effect of Pro-Inflammatory Cytokines on Drug Permeability
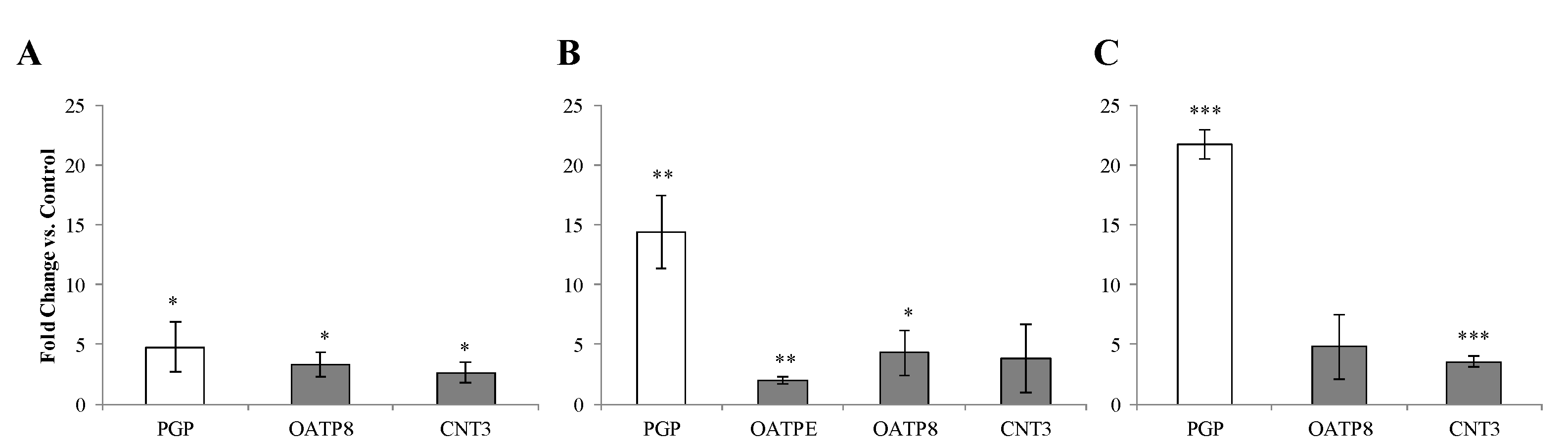
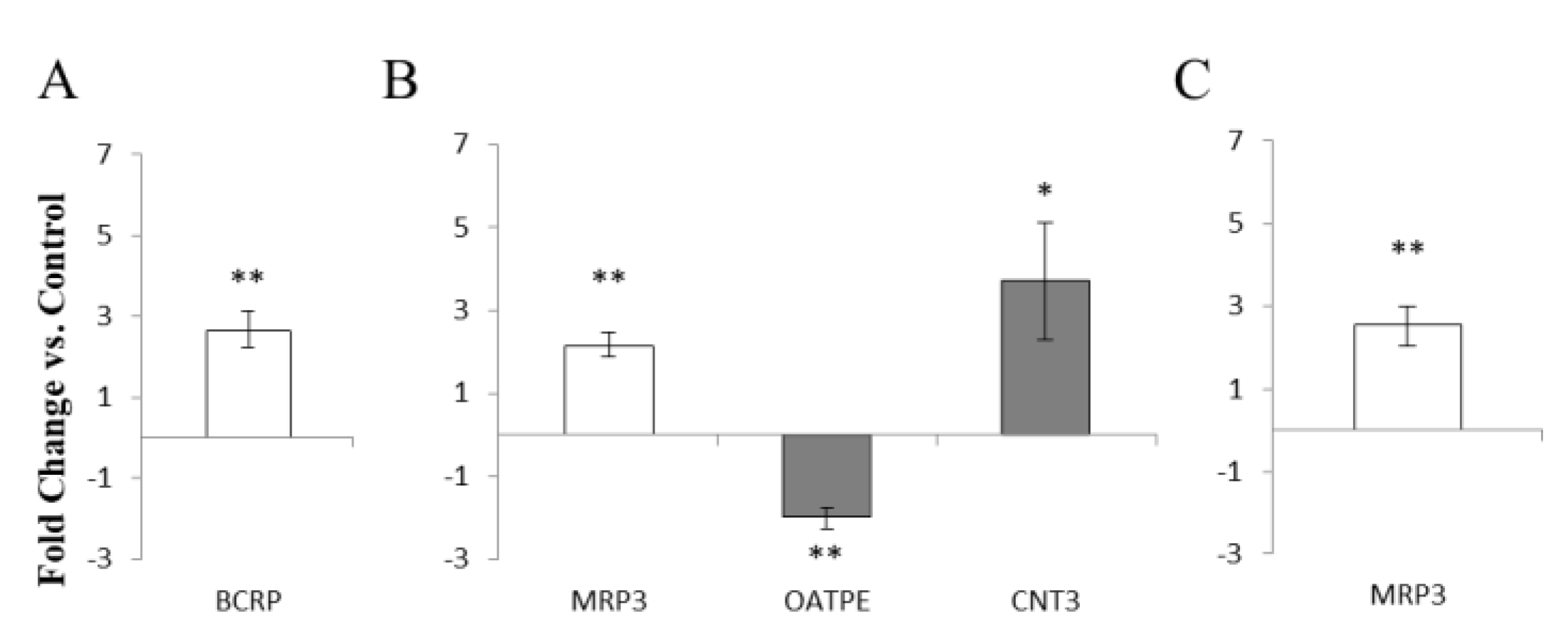
3.5. Environmental Effects on Vaginal Drug Transporter Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Klasse, P.J.; Shattock, R.; Moore, J.P. Antiretroviral drug-based microbicides to prevent HIV-1 sexual transmission. Annu. Rev. Med. 2008, 59, 455–471. [Google Scholar] [CrossRef]
- Hijazi, K.; Cuppone, A.M.; Smith, K.; Stincarelli, M.A.; Ekeruche-Makinde, J.; De Falco, G.; Hold, G.L.; Shattock, R.; Kelly, C.G.; Pozzi, G.; et al. Expression of Genes for Drug Transporters in the Human Female Genital Tract and Modulatory Effect of Antiretroviral Drugs. PLoS ONE 2015, 10, e0131405. [Google Scholar] [CrossRef]
- Hijazi, K.; Iannelli, F.; Cuppone, A.M.; Desjardins, D.; Caldwell, A.; Dereuddre-Bosquet, N.; Scala, C.; Smith, K.A.; Mukhopadya, I.; Frank, B.; et al. In Vivo Modulation of Cervicovaginal Drug Transporters and Tissue Distribution by Film-Released Tenofovir and Darunavir for Topical Prevention of HIV-1. Mol. Pharm. 2020, 17, 852–864. [Google Scholar] [CrossRef]
- Kis, O.; Robillard, K.; Chan, G.N.; Bendayan, R. The complexities of antiretroviral drug-drug interactions: Role of ABC and SLC transporters. Trends. Pharmacol. Sci. 2010, 31, 22–35. [Google Scholar] [CrossRef]
- Nicol, M.R.; Fedoriw, Y.; Mathews, M.; Prince, H.M.; Patterson, K.B.; Geller, E.; Mollan, K.; Mathews, S.; Kroetz, D.L.; Kashuba, A.D. Expression of six drug transporters in vaginal, cervical, and colorectal tissues: Implications for drug disposition in HIV prevention. J. Clin. Pharmacol. 2014, 54, 574–583. [Google Scholar] [CrossRef]
- Grammen, C.; Baes, M.; Haenen, S.; Verguts, J.; Augustyns, K.; Zydowsky, T.; La Colla, P.; Augustijns, P.; Brouwers, J. Vaginal expression of efflux transporters and the potential impact on the disposition of microbicides in vitro and in rabbits. Mol. Pharm. 2014, 11, 4405–4414. [Google Scholar] [CrossRef]
- Zhou, T.; Hu, M.; Pearlman, A.; Patton, D.; Rohan, L. Expression and localization of p-glycoprotein, multidrug resistance protein 4, and breast cancer resistance protein in the female lower genital tract of human and pigtailed macaque. AIDS Res. Hum. Retroviruses 2014, 30, 1106–1116. [Google Scholar] [CrossRef]
- Swedrowska, M.; Jamshidi, S.; Kumar, A.; Kelly, C.; Rahman, K.M.; Forbes, B. In Silico and in Vitro Screening for P-Glycoprotein Interaction with Tenofovir, Darunavir, and Dapivirine: An Antiretroviral Drug Combination for Topical Prevention of Colorectal HIV Transmission. Mol. Pharm. 2017, 14, 2660–2669. [Google Scholar] [CrossRef]
- Grammen, C.; Augustijns, P.; Brouwers, J. In vitro profiling of the vaginal permeation potential of anti-HIV microbicides and the influence of formulation excipients. Antiviral. Res. 2012, 96, 226–233. [Google Scholar] [CrossRef]
- Thews, O.; Gassner, B.; Kelleher, D.K.; Schwerdt, G.; Gekle, M. Impact of extracellular acidity on the activity of P-glycoprotein and the cytotoxicity of chemotherapeutic drugs. Neoplasia 2006, 8, 143–152. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Hirokawa, N.; Ohshiro, N.; Sekine, T.; Sasaki, T.; Tokuyama, S.; Endou, H.; Yamamoto, T. Differential gene expression of organic anion transporters in male and female rats. Biochem. Biophys. Res. Commun. 2002, 290, 482–487. [Google Scholar] [CrossRef]
- Boris, S.; Suarez, J.E.; Vazquez, F.; Barbes, C. Adherence of human vaginal lactobacilli to vaginal epithelial cells and interaction with uropathogens. Infect. Immun. 1998, 66, 1985–1989. [Google Scholar] [CrossRef]
- Lieberman, J.A.; Moscicki, A.B.; Sumerel, J.L.; Ma, Y.; Scott, M.E. Determination of cytokine protein levels in cervical mucus samples from young women by a multiplex immunoassay method and assessment of correlates. Clin. Vaccine Immunol. 2008, 15, 49–54. [Google Scholar] [CrossRef]
- Belec, L.; Gherardi, R.; Payan, C.; Prazuck, T.; Malkin, J.E.; Tevi-Benissan, C.; Pillot, J. Proinflammatory cytokine expression in cervicovaginal secretions of normal and HIV-infected women. Cytokine 1995, 7, 568–574. [Google Scholar] [CrossRef]
- Anderson, B.L.; Ghosh, M.; Raker, C.; Fahey, J.; Song, Y.; Rouse, D.J.; Wira, C.R.; Cu-Uvin, S. In vitro anti-HIV-1 activity in cervicovaginal secretions from pregnant and nonpregnant women. Am. J. Obstet. Gynecol. 2012, 207, 65.e1–65.e10. [Google Scholar] [CrossRef]
- Liptrott, N.J.; Penny, M.; Bray, P.G.; Sathish, J.; Khoo, S.H.; Back, D.J.; Owen, A. The impact of cytokines on the expression of drug transporters, cytochrome P450 enzymes and chemokine receptors in human PBMC. Br. J. Pharmacol. 2009, 156, 497–508. [Google Scholar] [CrossRef]
- Evseenko, D.A.; Paxton, J.W.; Keelan, J.A. Independent regulation of apical and basolateral drug transporter expression and function in placental trophoblasts by cytokines, steroids, and growth factors. Drug. Metab. Dispos. 2007, 35, 595–601. [Google Scholar] [CrossRef]
- Le Vee, M.; Gripon, P.; Stieger, B.; Fardel, O. Down-regulation of organic anion transporter expression in human hepatocytes exposed to the proinflammatory cytokine interleukin 1beta. Drug. Metab. Dispos. 2008, 36, 217–222. [Google Scholar] [CrossRef]
- Le Vee, M.; Lecureur, V.; Stieger, B.; Fardel, O. Regulation of drug transporter expression in human hepatocytes exposed to the proinflammatory cytokines tumor necrosis factor-alpha or interleukin-6. Drug Metab. Dispos. 2009, 37, 685–693. [Google Scholar] [CrossRef]
- Kim, W.Y.; Benet, L.Z. P-glycoprotein (P-gp/MDR1)-mediated efflux of sex-steroid hormones and modulation of P-gp expression in vitro. Pharm. Res. 2004, 21, 1284–1293. [Google Scholar] [CrossRef]
- Marcotte, H.; Ferrari, S.; Cesena, C.; Hammarstrom, L.; Morelli, L.; Pozzi, G.; Oggioni, M.R. The aggregation-promoting factor of Lactobacillus crispatus M247 and its genetic locus. J. Appl. Microbiol. 2004, 97, 749–756. [Google Scholar] [CrossRef]
- Hubatsch, I.; Ragnarsson, E.G.; Artursson, P. Determination of drug permeability and prediction of drug absorption in Caco-2 monolayers. Nat. Protoc. 2007, 2, 2111–2119. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Gali, Y.; Delezay, O.; Brouwers, J.; Addad, N.; Augustijns, P.; Bourlet, T.; Hamzeh-Cognasse, H.; Arien, K.K.; Pozzetto, B.; Vanham, G. In vitro evaluation of viability, integrity, and inflammation in genital epithelia upon exposure to pharmaceutical excipients and candidate microbicides. Antimicrob. Agents Chemother. 2010, 54, 5105–5114. [Google Scholar] [CrossRef]
- Caillouette, J.C.; Sharp, C.F., Jr.; Zimmerman, G.J.; Roy, S. Vaginal pH as a marker for bacterial pathogens and menopausal status. Am. J. Obstet. Gynecol. 1997, 176, 1270–1275, discussion 1275-1277. [Google Scholar] [CrossRef]
- Ravel, J.; Gajer, P.; Abdo, Z.; Schneider, G.M.; Koenig, S.S.; McCulle, S.L.; Karlebach, S.; Gorle, R.; Russell, J.; Tacket, C.O.; et al. Vaginal microbiome of reproductive-age women. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. 1), 4680–4687. [Google Scholar] [CrossRef]
- Schaefer, T.M.; Fahey, J.V.; Wright, J.A.; Wira, C.R. Innate immunity in the human female reproductive tract: Antiviral response of uterine epithelial cells to the TLR3 agonist poly(I:C). J. Immunol. 2005, 174, 992–1002. [Google Scholar] [CrossRef]
- Liu, H.; Chen, F.; Wu, W.; Cao, A.T.; Xue, X.; Yao, S.; Evans-Marin, H.L.; Li, Y.Q.; Cong, Y. TLR5 mediates CD172alpha(+) intestinal lamina propria dendritic cell induction of Th17 cells. Sci. Rep. 2016, 6, 22040. [Google Scholar] [CrossRef]
- Short, C.S.; Quinlan, R.; Bennett, P.; Shattock, R.J.; Taylor, G.P. Optimising the collection of female genital tract fluid for cytokine analysis in pregnant women. J. Immunol. Methods 2018, 458, 15–20. [Google Scholar] [CrossRef]
- Comar, M.; Monasta, L.; Zanotta, N.; Vecchi Brumatti, L.; Ricci, G.; Zauli, G. Human papillomavirus infection is associated with decreased levels of GM-CSF in cervico-vaginal fluid of infected women. J. Clin. Virol. 2013, 58, 479–481. [Google Scholar] [CrossRef]
- Bajaj, R.; Chong, L.B.; Zou, L.; Tsakalozou, E.; Ni, Z.; Giacomini, K.M.; Kroetz, D.L. Interaction of Commonly Used Oral Molecular Excipients with P-glycoprotein. AAPS J. 2021, 23, 106. [Google Scholar] [CrossRef] [PubMed]
- Gudnadottir, U.; Debelius, J.W.; Du, J.; Hugerth, L.W.; Danielsson, H.; Schuppe-Koistinen, I.; Fransson, E.; Brusselaers, N. The vaginal microbiome and the risk of preterm birth: A systematic review and network meta-analysis. Sci. Rep. 2022, 12, 7926. [Google Scholar] [CrossRef] [PubMed]
- Thurman, A.R.; Schwartz, J.L.; Brache, V.; Chen, B.A.; Chandra, N.; Kashuba, A.D.M.; Weiner, D.H.; Mauck, C.; Doncel, G.F. Effect of Hormonal Contraception on Pharmacokinetics of Vaginal Tenofovir in Healthy Women: Increased Tenofovir Diphosphate in Injectable Depot Medroxyprogesterone Acetate Users. J. Acquir. Immune Defic. Syndr. 2019, 80, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhya, I.; Murray, G.I.; Berry, S.; Thomson, J.; Frank, B.; Gwozdz, G.; Ekeruche-Makinde, J.; Shattock, R.; Kelly, C.; Iannelli, F.; et al. Drug transporter gene expression in human colorectal tissue and cell lines: Modulation with antiretrovirals for microbicide optimization. J. Antimicrob. Chemother. 2016, 71, 372–386. [Google Scholar] [CrossRef]
- Mukhopadhya, I.; Murray, G.I.; Duncan, L.; Yuecel, R.; Shattock, R.; Kelly, C.; Iannelli, F.; Pozzi, G.; El-Omar, E.M.; Hold, G.L.; et al. Transporters for Antiretroviral Drugs in Colorectal CD4+ T Cells and Circulating alpha4beta7 Integrin CD4+ T Cells: Implications for HIV Microbicides. Mol. Pharm. 2016, 13, 3334–3340. [Google Scholar] [CrossRef]
- Kwan, W.S.; Janneh, O.; Hartkoorn, R.; Chandler, B.; Khoo, S.; Back, D.; Owen, A. Intracellular ’boosting’ of darunavir using known transport inhibitors in primary PBMC. Br. J. Clin. Pharmacol. 2009, 68, 375–380. [Google Scholar] [CrossRef][Green Version]
- Cuevas, M.E.; Gaska, J.M.; Gist, A.C.; King, J.M.; Sheller, R.A.; Todd, M.C. Estrogen-dependent expression and subcellular localization of the tight junction protein claudin-4 in HEC-1A endometrial cancer cells. Int. J. Oncol. 2015, 47, 650–656. [Google Scholar] [CrossRef]
- Bashir, T.; Reddy, K. HbAHP-25 attenuates HIV-1 gp120 mediated inflammation and barrier dysfunction. HIV Med. 2018, 19, 206–215. [Google Scholar] [CrossRef]
- Le Ferrec, E.; Chesne, C.; Artusson, P.; Brayden, D.; Fabre, G.; Gires, P.; Guillou, F.; Rousset, M.; Rubas, W.; Scarino, M.L. In vitro models of the intestinal barrier. The report and recommendations of ECVAM Workshop 46. European Centre for the Validation of Alternative methods. Altern. Lab. Anim. 2001, 29, 649–668. [Google Scholar] [CrossRef]
- Vachon, P.H.; Durand, J.; Beaulieu, J.F. Basement membrane formation and re-distribution of the beta 1 integrins in a human intestinal co-culture system. Anat. Rec. 1993, 235, 567–576. [Google Scholar] [CrossRef]
- Dezzutti, C.S.; Rohan, L.C.; Wang, L.; Uranker, K.; Shetler, C.; Cost, M.; Lynam, J.D.; Friend, D. Reformulated tenofovir gel for use as a dual compartment microbicide. J. Antimicrob. Chemother. 2012, 67, 2139–2142. [Google Scholar] [CrossRef] [PubMed]
- Akil, A.; Devlin, B.; Cost, M.; Rohan, L.C. Increased Dapivirine tissue accumulation through vaginal film codelivery of dapivirine and Tenofovir. Mol. Pharm. 2014, 11, 1533–1541. [Google Scholar] [CrossRef] [PubMed]
- Errasti-Murugarren, E.; Pastor-Anglada, M.; Casado, F.J. Role of CNT3 in the transepithelial flux of nucleosides and nucleoside-derived drugs. J. Physiol. 2007, 582, 1249–1260. [Google Scholar] [CrossRef] [PubMed]
- Hau, R.K.; Tash, J.S.; Georg, G.I.; Wright, S.H.; Cherrington, N.J. Physiological Characterization of the Transporter-Mediated Uptake of the Reversible Male Contraceptive H2-Gamendazole Across the Blood-Testis Barrier. J. Pharmacol. Exp. Ther. 2022, 382, 299–312. [Google Scholar] [CrossRef]
- Capaldo, C.T.; Nusrat, A. Cytokine regulation of tight junctions. Biochim. Biophys. Acta 2009, 1788, 864–871. [Google Scholar] [CrossRef]
- Ma, T.Y.; Iwamoto, G.K.; Hoa, N.T.; Akotia, V.; Pedram, A.; Boivin, M.A.; Said, H.M. TNF-alpha-induced increase in intestinal epithelial tight junction permeability requires NF-kappa B activation. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 286, G367–G376. [Google Scholar] [CrossRef]
- Chen, M.L.; Ge, Z.; Fox, J.G.; Schauer, D.B. Disruption of tight junctions and induction of proinflammatory cytokine responses in colonic epithelial cells by Campylobacter jejuni. Infect. Immun. 2006, 74, 6581–6589. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drug Transporter | NCBI Ref Sequence | Forward Primer 5′ > 3′ | Reverse Primer 5′ > 3′ |
|---|---|---|---|
| HPRT1 | NG_012329.2 | TGACCTTGATTTATTTTGCATACC | CGAGCAAGACGTTCAGTCCT |
| RPLP0 | NC_000012.12 | TCCAGGCTTTAGGTATCACCAC | TTGATCAGCTGCACATCACTC |
| P-gp | NG_011513.1 | CTTCATCGAGTCACTGCCTAATAA | GCTATGGCAATGCGTTGTT |
| BCRP | NG_032067.2 | TGGCTTAGACTCAAGCACAGC | TCGTCCCTGCTTAGACATCC |
| MRP1 | NG_028268.2 | CCTGTTCAACGTCATTGGTG | AGGAAGCCACGTAGAACCTCT |
| MRP2 | NG_011798.2 | CTGGATCACCTCCAACAGGT | AGAAGACAGTCAGGTTCCCAAC |
| MRP3 | NC_000017.11 | AATGTCGACCCTAACCCCTAC | GGGTCAGGGTTTAGCAGCTT |
| MRP4 | NG_050651.2 | GCGCCTGGAATCTACAACTC | CTTTGTATGCCCGGATGGT |
| MRP5 | NG_047115.1 | CAGCGACCTGACGGAGAT | GGGCAAGGCTGATCCTCT |
| MRP6 | NG_007558.3 | CGGGAAGGATTGCATCAC | GGCACCGTGAGGTTTATTCT |
| MRP7 | NC_003074.8 | GCCCTCAATGATGACCTCAG | CCTCCGCTAAGGGTGACA |
| ENT1 | NG_042893.1 | TCACCAGCCTCAGGACAGAT | GAGTGGCCGTCATGAAAAA |
| ENT2 | NC_000011.10 | GCTGAGCTCCTCCAGTCTGA | GGCTCATCTGGCTCTGATTC |
| CNT3 | NC_000009.12 | CTGAACTCCACGCCATCA | CAAGTGGGAGGATGGAACC |
| OCT1 | NC_000006.12 | CTCGCCCAACTACATGTCC | CGAGCCAACAAATTCTGTGAT |
| OCT3 | NC_000006.12 | CACCATCGTCAGCGAGTTT | CAGGATGGCTTGGGTGAG |
| OATPD | NC_000015.10 | TGTTCCCTATGGAAACAGCA | CAGGTAGGTGATGCCATCTG |
| OATPE | NC_000020.11 | AGCTGCCACCTTGTTTGG | GCCTGAGCTTGTTCACAAAGA |
| OATP8 | NG_032071.1 | TCAAGTGGTATTAAAAAGCATACAGTG | TTCACCCAAGTGTGCTGAGT |
| Drug Name (Concentration) | % Drug Distribution * | ||
|---|---|---|---|
| Apical | Basolateral | Cell-Associated | |
| TFV (0.1 μM) | 97.03 ± 0.28 | 2.63 ± 0.25 | 0.34 ± 0.06 |
| TFV (1 μM) | 96.85 ± 0.29 | 2.73 ± 0.24 | 0.42 ± 0.08 |
| TFV (10 μM) | 96.91 ± 0.27 | 2.79 ± 0.25 | 0.3 ± 0.06 |
| TFV (100 μM) | 96.59 ± 0.41 | 2.99 ± 0.36 | 0.42 ± 0.17 |
| DRV (1 μM) | 82.90 ± 1.76 | 15.13 ± 1.44 | 1.97 ± 1.97 |
| DRV (5 μM) | 83.15 ± 2.61 | 14.55 ± 1.88 | 2.55 ± 1.22 |
| DRV (10 μM) | 83.36 ± 2.12 | 13.95 ± 1.75 | 2.70 ± 1.31 |
| DRV (20 μM) | 81.72 ± 1.61 | 15.31 ± 1.22 | 2.97 ± 1.27 |
| DPV (0.1 μM) | 15.04 ± 0.55 | 2.87 ± 0.26 | 82.10 ± 0.30 |
| DPV (1 μM) | 14.72 ± 0.10 | 2.13 ± 0.33 | 83.15 ± 0.35 |
| DPV (5 μM) | 14.98 ± 0.76 | 1.86 ± 0.41 | 83.16 ± 1.15 |
| DPV (10 μM) | 14.52 ± 0.45 | 2.09 ± 0.18 | 83.39 ± 0.55 |
| DPV (0.1 μM) + Poloxamer | 16.06 ± 3.26 | 10.56 ± 1.26 | 73.37 ± 3.37 |
| DPV (1 μM) + Poloxamer | 16.47 ± 2.86 | 11.01 ± 1.41 | 72.51 ± 2.86 |
| DPV (5 μM) + Poloxamer | 17.47± 2.76 | 10.78 ± 1.18 | 71.75 ± 2.33 |
| DPV (10 μM) + Poloxamer | 17.59 ± 3.41 | 11.72 ± 1.34 | 70.69 ± 2.76 |
| Lactobacillus crispatus | Candida albicans | |||
|---|---|---|---|---|
| HEC-1A | VK2/E6E7 | HEC-1A | VK2/E6E7 | |
| P-GP | - | - | - | - |
| BCRP | ↑ | - | - | - |
| MRP1 | - | - | - | - |
| MRP2 | - | - | - | - |
| MRP3 | - | - | - | ↑ |
| MRP4 | - | - | ↓ | - |
| MRP5 | - | - | - | - |
| MRP6 | - | - | - | - |
| MRP7 | - | - | - | - |
| OATP8 | - | ↑↑ | - | - |
| OATPD | - | - | - | - |
| OATPE | - | ↑↑ | - | ↑↑↑ |
| OCT1 | - | ↑ | - | - |
| OCT3 | - | - | - | - |
| CNT3 | - | ↑ | ↑ | - |
| ENT1 | - | - | - | - |
| ENT2 | - | - | ↓ | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carserides, C.; Smith, K.; Zinicola, M.; Kumar, A.; Swedrowska, M.; Scala, C.; Cameron, G.; Riches, Z.; Iannelli, F.; Pozzi, G.; et al. Comprehensive Study of Antiretroviral Drug Permeability at the Cervicovaginal Mucosa via an In Vitro Model. Pharmaceutics 2022, 14, 1938. https://doi.org/10.3390/pharmaceutics14091938
Carserides C, Smith K, Zinicola M, Kumar A, Swedrowska M, Scala C, Cameron G, Riches Z, Iannelli F, Pozzi G, et al. Comprehensive Study of Antiretroviral Drug Permeability at the Cervicovaginal Mucosa via an In Vitro Model. Pharmaceutics. 2022; 14(9):1938. https://doi.org/10.3390/pharmaceutics14091938
Chicago/Turabian StyleCarserides, Constandinos, Kieron Smith, Marta Zinicola, Abhinav Kumar, Magda Swedrowska, Carlo Scala, Gary Cameron, Zoe Riches, Francesco Iannelli, Gianni Pozzi, and et al. 2022. "Comprehensive Study of Antiretroviral Drug Permeability at the Cervicovaginal Mucosa via an In Vitro Model" Pharmaceutics 14, no. 9: 1938. https://doi.org/10.3390/pharmaceutics14091938
APA StyleCarserides, C., Smith, K., Zinicola, M., Kumar, A., Swedrowska, M., Scala, C., Cameron, G., Riches, Z., Iannelli, F., Pozzi, G., Hold, G. L., Forbes, B., Kelly, C., & Hijazi, K. (2022). Comprehensive Study of Antiretroviral Drug Permeability at the Cervicovaginal Mucosa via an In Vitro Model. Pharmaceutics, 14(9), 1938. https://doi.org/10.3390/pharmaceutics14091938