
Towards Selective Binding to the GLUT5 Transporter: Synthesis, Molecular Dynamics and In Vitro Evaluation of Novel C-3-Modified 2,5-Anhydro-D-mannitol Analogs



 , ,
, , 
Abstract
:
1. Introduction
2. Materials and Methods
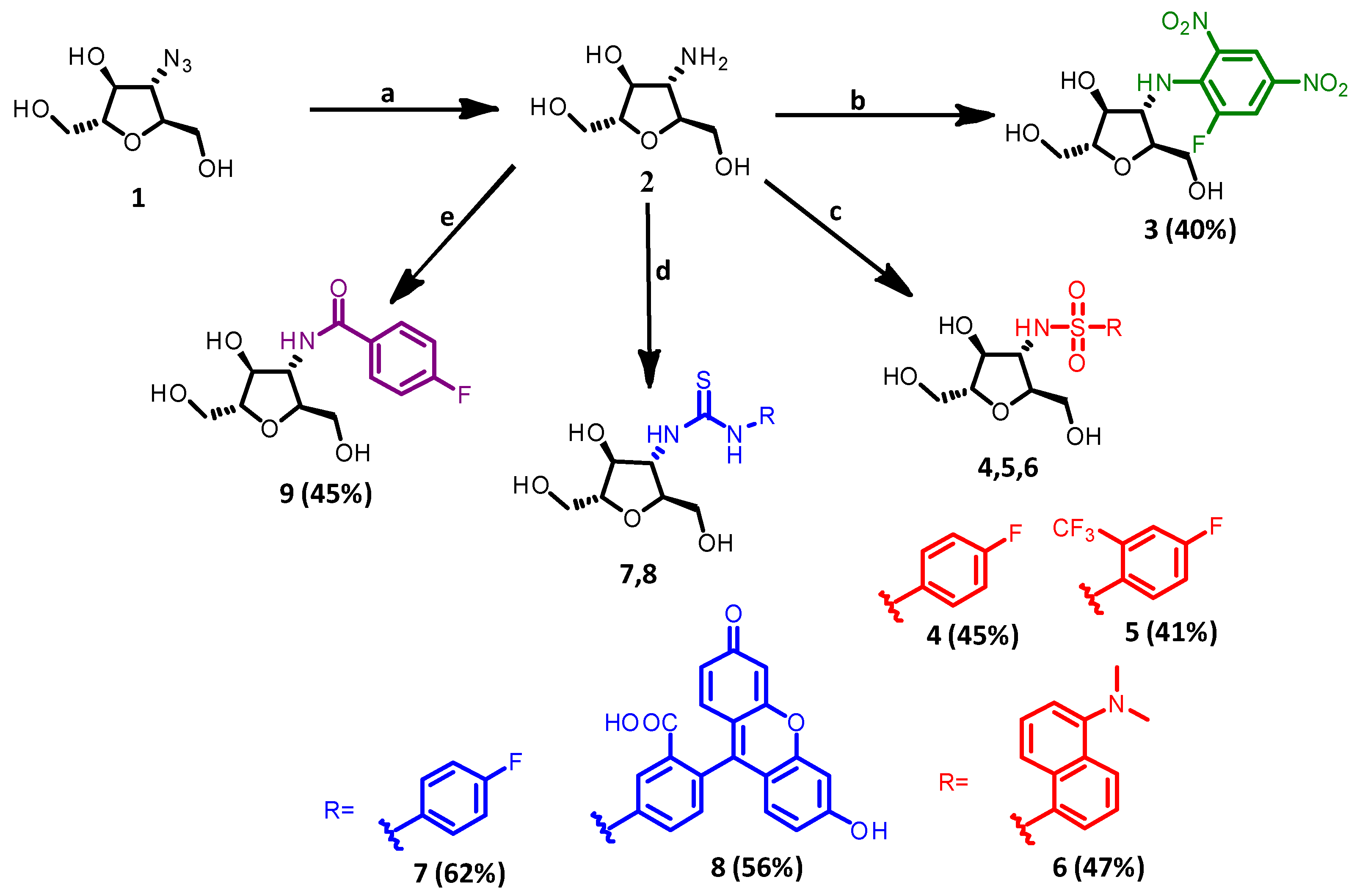
2.1. Synthesis
- 3-deoxy-3-amino-2,5-anhydro-D-mannitol (2)
- 3-deoxy-3-[N-(5-fluoro-2,4-dinitro-phenyl)amino]-2,5-anhydro-D-mannitol (3)
2.1.1. General Procedure to Synthesize 4, 5 and 6
- 3-deoxy-3-[N-(4-fluorobenzenesulfonamide)amino]-2,5-anhydro-D-mannitol (4)
- 3-deoxy-3-[N-(4-fluoro-2-(trifluoromethyl)benzenesulfonamide)amino]-2,5-anhydro-D-mannitol (5)
- 3-deoxy-3-[N-(5-(dimethylamino)naphthalene-1-sulfonamide)amino]-2,5-anhydro-D-mannitol (6)
2.1.2. General Procedure to Synthesize 7 and 8
- 3-deoxy-3-[N-(1-(4-fluorophenyl)thiourea)amino]-2,5-anhydro-D-mannitol (7)
- 3-deoxy-3-[N-(3-(fluorescein)-5-yl)thiourea)amino]-2,5-anhydro-D-mannitol (8)
- 3-deoxy-3-[N-(4-fluorobenzamide)amino]-2,5-anhydro-D-mannitol (9)
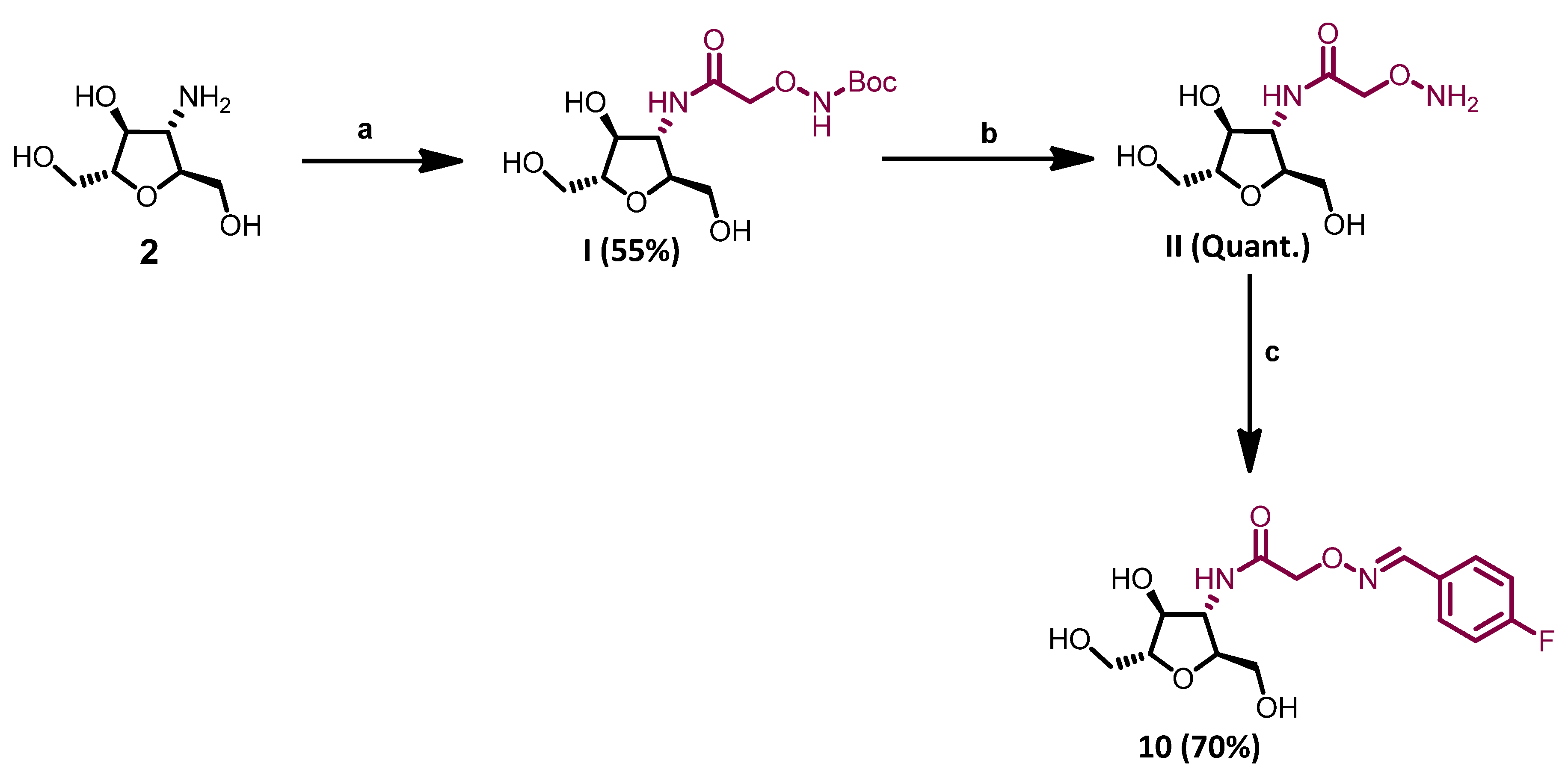
- 3-deoxy-3-[N-(tert-butyl-2-amino-2-oxoethoxycarbamate)]-2,5-anhydro-D-mannitol (I)
- 3-deoxy-3-[N-(2-(((4-fluorobenzylidene)amino)oxy)acetamide)amino]-2,5-anhydro-D-mannitol (10)
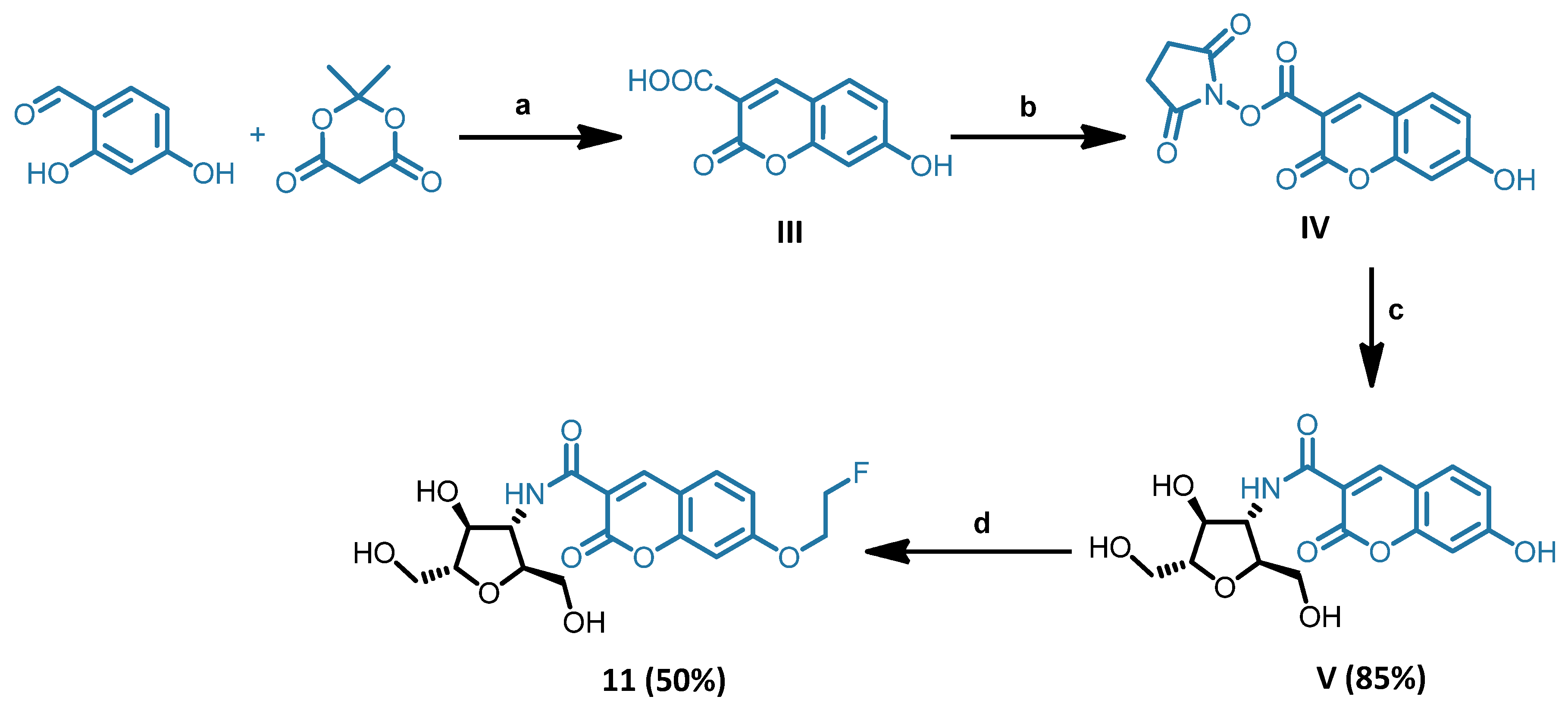
- 3-deoxy-3-[N-(7-hydroxy-2-oxo-2H-chromene-3-carboxamide)amino]-2,5-anhydro-D-mannitol (V)
- 3-deoxy-3-[N-(7-(2-fluoroethoxy)-2-oxo-2H-chromene-3-carboxamide)amino]-2,5-anhydro-D-mannitol (11)
2.2. In Vitro Cell Experiments
2.2.1. Instruments
2.2.2. Buffer Solutions
2.2.3. Radiotracer Synthesis
2.2.4. Cell Culture
2.2.5. General Procedure for In Vitro Inhibition of 6-[18F]FDF Cell Uptake
- Fructose—freely soluble in Krebs–Ringer buffer;
- 2,5-AM derivatives—all the samples were first dissolved in ≤0.1% DMSO and were further diluted using Krebs–Ringer buffer according to the desired concentration maintaining ≤0.1% DMSO;
- Blank—0.1% DMSO.
2.3. Molecular Docking
Molecular Dynamics Simulations
3. Results and Discussion
3.1. Synthesis of C-3-Modified 2,5-AM Compounds
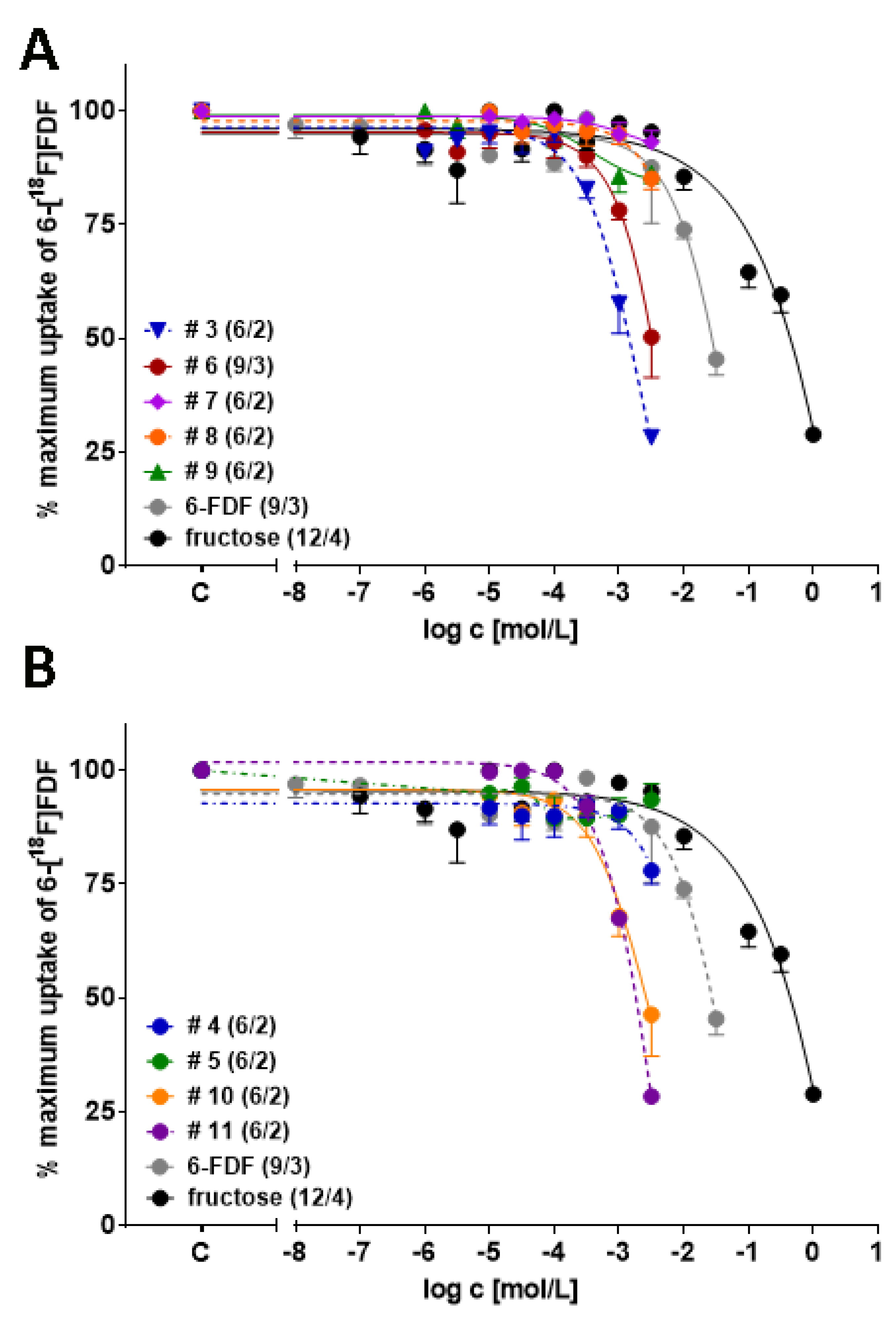
3.2. In Vitro Cell Experiments
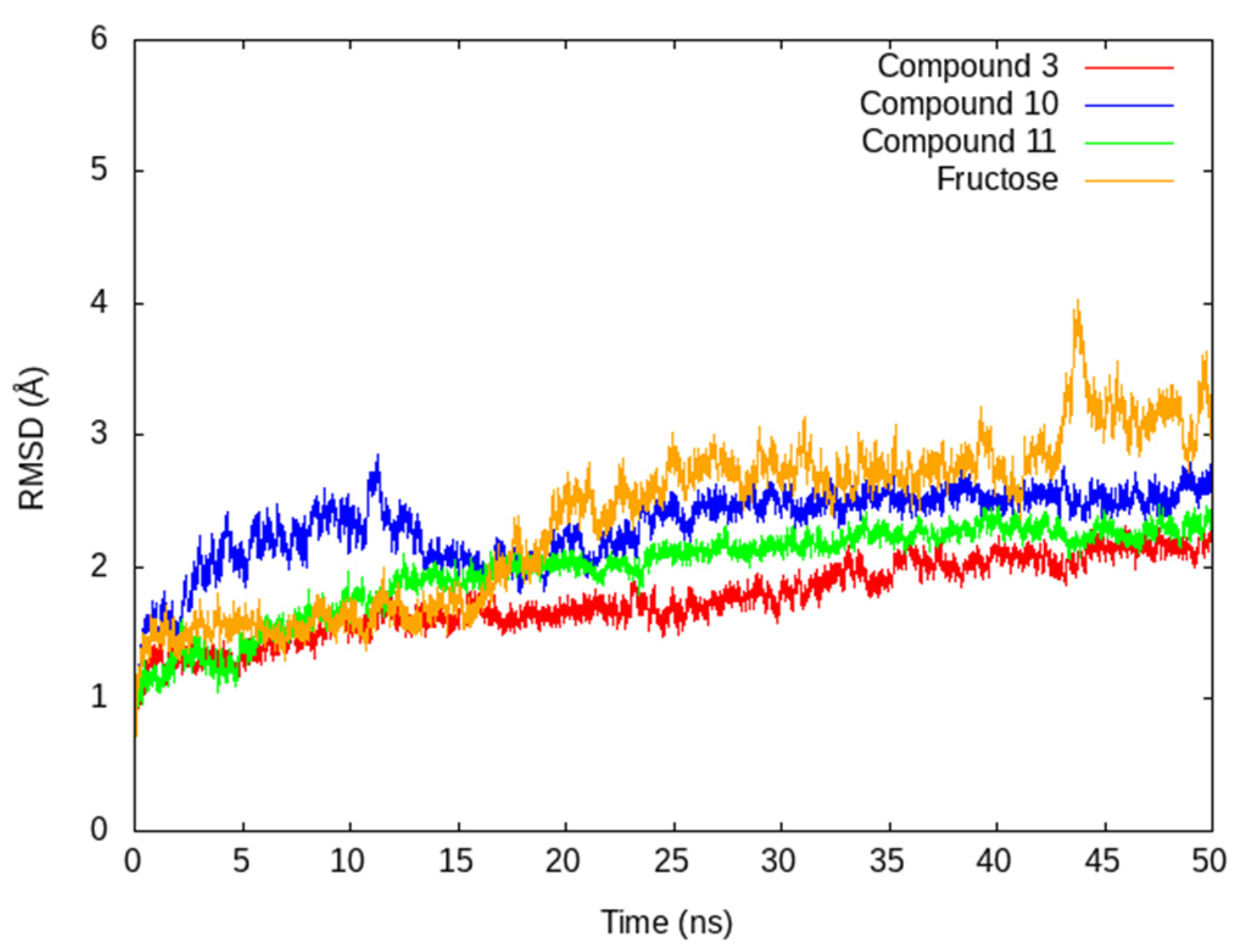
3.3. In Silico Studies
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer Statistics. CA Cancer J. Clin. 2018, 67, 7–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamora-León, S.P.; Golde, D.W.; Concha, I.I.; Rivas, C.I.; Delgado-López, F.; Baselga, J.; Nualart, F.; Vera, J.C. Expression of the fructose transporter GLUT5 in human breast cancer. Proc. Natl. Acad. Sci. USA 1996, 93, 1847–1852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marger, M.D.; Saier, M.H., Jr. A major superfamily of transmembrane facilitators that catalyse uniport, symport and antiport. Trends Biochem. Sci. 1993, 18, 13–20. [Google Scholar] [CrossRef]
- Manolescu, A.R.; Witkowska, K.; Kinnaird, A.; Cessford, T.; Cheeseman, C. Facilitated hexose transporters: New perspectives on form and function. Physiology 2007, 22, 234–240. [Google Scholar] [CrossRef]
- García-Álvarez, I.; Garrido, L.; Fernández-Mayoralas, A. Studies on the uptake of glucose derivatives by red blood cells. ChemMedChem 2007, 2, 496–504. [Google Scholar] [CrossRef]
- McQuade, D.T.; Plutschack, M.B.; Seeberger, P.H. Passive fructose transporters in disease: A molecular overview of their structural specificity. Org. Biomol. Chem. 2013, 11, 4909–4920. [Google Scholar] [CrossRef]
- Mueckler, M.; Thorens, B. The SLC2 (GLUT) family of membrane transporters. Mol. Asp. Med. 2013, 34, 121–138. [Google Scholar] [CrossRef] [Green Version]
- Tappy, L.; Lê, K.A. Metabolic effects of fructose and the worldwide increase in obesity. Physiol. Rev. 2010, 90, 23–46. [Google Scholar] [CrossRef] [Green Version]
- Gaby, A.R. Adverse effects of dietary fructose. Altern. Med. Rev. 2005, 10, 294–306. [Google Scholar]
- Camps, M.; Castelló, A.; Muñoz, P.; Monfar, M.; Testar, X.; Palacín, M.; Zorzano, A. Effect of diabetes and fasting on GLUT-4 (muscle/fat) glucose-transporter expression in insulin-sensitive tissues. Heterogeneous response in heart, red and white muscle. Biochem. J. 1992, 282, 765–772. [Google Scholar] [CrossRef] [Green Version]
- Medina, R.A.; Owen, G.I. Glucose transporters: Expression, regulation, and cancer. Biol. Res. 2002, 35, 9–26. [Google Scholar] [CrossRef] [PubMed]
- Adekola, K.; Rosen, S.T.; Shanmugam, M. Glucose transporters in cancer metabolism. Curr. Opin. Oncol. 2012, 24, 650–654. [Google Scholar] [CrossRef]
- Lin, L.; Yee, S.W.; Kim, R.B.; Giacomini, K.M. SLC transporters as therapeutic targets: Emerging opportunities. Nat. Rev. Drug Discov. 2015, 14, 543–560. [Google Scholar] [CrossRef] [Green Version]
- Colas, C.; Ung, P.M.; Schlessinger, A. SLC transporters: Structure, function, and drug discovery. Med. Chem. Comm. 2016, 7, 1069–1081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotsampasakou, E.; Ecker, G.F.; Sitte, H.H.; Mannhold, R.; Buschmann, H.; Clausen, R.P. Organic Anion Transporting Polypeptides as Drug Targets, in Transporters as Drug Targets. In Transporters as Drug Targets; Wiley—VCH Verlag GmbH & Co. KGa: Hoboken, NJ, USA, 2017; pp. 271–324. [Google Scholar]
- Kostakoglu, L.; Agress, H., Jr.; Goldsmith, S.J. Clinical role of FDG PET in evaluation of cancer patients. RadioGraphics 2003, 23, 315–340. [Google Scholar] [CrossRef] [PubMed]
- Joost, H.G.; Bell, G.I.; Best, J.D.; Birnbaum, M.J.; Charron, M.J.; Chen, Y.T.; Doege, H.; James, D.E.; Lodish, H.F.; Moley, K.H.; et al. Nomenclature of the GLUT/SLC2A family of sugar/polyol transport facilitators. Am. J. Physiol. Metab. 2002, 282, E974–E976. [Google Scholar] [CrossRef] [Green Version]
- Fan, X.; Liu, H.; Liu, M.; Wang, Y.; Qiu, L.; Cui, Y. Increased utilization of fructose has a positive effect on the development of breast cancer. PeerJ 2017, 5, e3804. [Google Scholar] [CrossRef]
- Chen, W.L.; Wang, Y.Y.; Zhao, A.; Xia, L.; Xie, G.; Su, M.; Zhao, L.; Liu, J.; Qu, C.; Wei, R.; et al. Enhanced fructose utilization mediated by SLC2A5 is a unique metabolic feature of acute myeloid leukemia with therapeutic potential. Cancer Cell 2016, 30, 779–791. [Google Scholar] [CrossRef] [Green Version]
- Weng, Y.; Zhu, J.; Chen, Z.; Fu, J.; Zhang, F. Fructose fuels lung adenocarcinoma through GLUT5. Cell Death Dis. 2018, 9, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Villaamil, V.M.; Gallego, G.A.; Rubira, L.V.; Campelo, R.G.; Valladares-Ayersbes, M.; Pulido, E.G.; Bolós, M.V.; Caínzos, I.S.; Aparicio, L.M.A. Fructose transporter GLUT5 expression in clear renal cell carcinoma. Oncol. Rep. 2011, 25, 315–323. [Google Scholar]
- Hamann, I.; Krys, D.; Glubrecht, D.; Bouvet, V.; Marshall, A.; Vos, L.; Mackey, J.R.; Wuest, M.; Wuest, F. Expression and function of hexose transporters GLUT1, GLUT2, and GLUT5 in breast cancer—effects of hypoxia. FASEB J. 2018, 32, 5104–5118. [Google Scholar] [CrossRef]
- Douard, V.; Ferraris, R.P. Regulation of the fructose transporter GLUT5 in health and disease. Am. J. Physiol. Endocrinol. Metab. 2008, 238, E227–E237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kane, S.; Seatter, M.J.; Gould, G.W. Functional studies of human GLUT5: Effect of pH on substrate selection and an analysis of substrate interactions. Biochem. Biophys. Res. Commun. 1997, 238, 503–505. [Google Scholar] [CrossRef] [PubMed]
- Mate, A.; Barfull, A.; Hermosa, A.M.; Planas, J.M.; Vazquez, C.M. Regulation of D-fructose transporter GLUT5 in the ileum of spontaneously hypertensive rats. J. Membr. Biol. 2004, 199, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Gorovits, N.; Charron, M.J. What we know about facilitative glucose transporters: Lessons from cultured cells, animal models, and human studies. Biochem. Mol. Biol. Educ. 2008, 31, 163–172. [Google Scholar] [CrossRef]
- Nomura, N.; Verdon, G.; Kang, H.J.; Shimamura, T.; Nomura, Y.; Sonoda, Y.; Hussien, S.A.; Qureshi, A.A.; Coincon, M.; Sato, Y.; et al. Structure and mechanism of the mammalian fructose transporter GLUT5. Nature 2015, 526, 397–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanasova, M.; Fedie, J.R. Molecular tools for facilitative carbohydrate transporters (Gluts). ChemBioChem 2017, 18, 1774–1788. [Google Scholar] [CrossRef] [PubMed]
- Girniene, J.; Tatibouët, A.; Sackus, A.; Yang, J.; Holman, G.D.; Rollin, P. Inhibition of the D-fructose transporter protein GLUT5 by fused-ring glyco-1, 3-oxazolidin-2-thiones and-oxazolidin-2-ones. Carbohydr. Res. 2003, 338, 711–719. [Google Scholar] [CrossRef]
- Barnett, J.E.G.; Holman, G.D.; Munday, K.A. Structural requirements for binding to the sugar-transport system of the human erythrocyte. Biochem. J. 1973, 131, 211–221. [Google Scholar] [CrossRef] [Green Version]
- Barnett, J.E.; Holman, G.D.; Chalkley, R.A.; Munday, K.A. Evidence for two asymmetric conformational states in the human erythrocyte sugar-transport system. Biochem. J. 1975, 145, 417–429. [Google Scholar] [CrossRef] [Green Version]
- Rees, W.D.; Holman, G.D. Hydrogen bonding requirements for the insulin-sensitive sugar transport system of rat adipocytes. Biochim. Biophys. Acta- Biomembr. 1981, 646, 251–260. [Google Scholar] [CrossRef]
- Holman, G.D.; Rees, W.D. Side-specific analogues for the rat adipocyte sugar transport system. Biochim. Biophys. Acta- Biomembr. 1982, 685, 78–86. [Google Scholar] [CrossRef]
- Holman, G.D.; Midgley, P.J.W. Synthesis of novel bis (D-mannose) compounds. Carbohydr. Res. 1985, 135, 337–341. [Google Scholar] [CrossRef]
- Abbadi, M.; Holman, G.D.; Morin, C.; Rees, W.D.; Yang, J. Synthesis of symmetrical 4, 4′-and 6, 6′-bis (d-glucose)-based probes as tools for the study of d-glucose transport proteins. Tetrahedron Lett. 1999, 40, 5861–5864. [Google Scholar] [CrossRef]
- Bouvet, V.; Jans, H.S.; Wuest, M.; Soueidan, O.M.; Mercer, J.; McEwan, A.J.B.; West, F.G.; Cheeseman, C.I.; Wuest, F. Automated synthesis and dosimetry of 6-deoxy-6-[18F] fluoro-D-fructose (6-[18F] FDF): A radiotracer for imaging of GLUT5 in breast cancer. Am. J. Nucl. Med. Mol. Imaging 2014, 4, 248–259. [Google Scholar]
- Soueidan, O.M.; Scully, T.W.; Kaur, J.; Panigrahi, R.; Belovodskiy, A.; Do, V.; Matier, C.D.; Lemieux, M.J.; Wuest, F.; Cheeseman, C.I.; et al. Fluorescent hexose conjugates establish stringent stereochemical requirement by glut5 for recognition and transport of monosaccharides. ACS Chem. Biol. 2017, 12, 1087–1094. [Google Scholar] [CrossRef]
- Wuest, M.; Hamann, I.; Bouvet, V.; Glubrecht, D.; Marshall, A.; Trayner, B.; Soueidan, O.M.; Krys, D.; Wagner, M.; Cheeseman, C.; et al. Molecular imaging of GLUT1 and GLUT5 in breast cancer: A multitracer positron emission tomography imaging study in mice. Mol. Pharmacol. 2018, 93, 79–89. [Google Scholar] [CrossRef]
- Tatibouët, A.; Yang, J.; Morin, C.; Holman, G.D. Synthesis, and evaluation of fructose analogues as inhibitors of the D-fructose transporter GLUT5. Bioorg. Med. Chem. 2000, 8, 1825–1833. [Google Scholar] [CrossRef]
- Inukai, K.; Katagiri, H.; Takata, K.; Asano, T.; Anai, M.; Ishihara, H.; Nakazaki, M.; Kikuchi, M.; Yazaki, Y.; Oka, Y. Characterization of rat GLUT5 and functional analysis of chimeric proteins of GLUT1 glucose transporter and GLUT5 fructose transporter. Endocrinology 1995, 136, 4850–4857. [Google Scholar] [CrossRef]
- Soueidan, O.M.; Trayner, B.J.; Grant, T.N.; Henderson, J.R.; Wuest, F.; West, F.G.; Cheeseman, C.I. New fluorinated fructose analogs as selective probes of the hexose transporter protein GLUT5. Org. Biomol. Chem. 2015, 13, 6511–6521. [Google Scholar] [CrossRef]
- Tanasova, M.; Plutschack, M.; Muroski, M.E.; Sturla, S.J.; Strouse, G.F.; McQuade, D.T. Fluorescent THF-based fructose analogue exhibits fructose-dependent uptake. ChemBioChem 2013, 14, 1263–1270. [Google Scholar] [CrossRef] [PubMed]
- Kumar Kondapi, V.P.; Soueidan, O.M.; Hosseini, S.N.; Jabari, N.; West, F.G. Efficient and Easy Access to Optically Pure Tetrasubstituted Tetrahydrofurans via Stereoselective Opening of C2-Symmetric Epoxide and Aziridine Rings. Eur. J. Org. Chem. 2016, 7, 1367–1379. [Google Scholar] [CrossRef]
- Horton, D.; Philips, K.D. The nitrous acid deamination of glycosides and acetates of 2-amino-2-deoxy-D-glucose. Carbohydr. Res. 1973, 30, 367–374. [Google Scholar] [CrossRef]
- Brennauer, A. Acylguanidines as Bioisosteric Groups in Argininamide-Type Neuropeptide Y Y1 and Y2 Receptor Antagonists: Synthesis, Stability and Pharmacological Activity. Ph.D. Thesis, University of Regensburg, Regensburg, Germany, 2006. [Google Scholar]
- Tang, F.; Yang, Y.; Tang, Y.; Tang, S.; Yang, L.; Sun, B.; Jiang, B.; Dong, J.; Liu, H.; Huang, M.; et al. One-pot N-glycosylation remodeling of IgG with non-natural sialylglycopeptides enables glycosite-specific and dual-payload antibody–drug conjugates. Org. Biomol. Chem. 2016, 14, 9501–9518. [Google Scholar] [CrossRef] [PubMed]
- Gnaccarini, C.; Ben-Tahar, W.; Mulani, A.; Roy, I.; Lubell, W.D.; Pelletier, J.N.; Keillor, J.W. Site-specific protein propargylation using tissue transglutaminase. Org. Biomol. Chem. 2012, 10, 5258–5265. [Google Scholar] [CrossRef]
- Shiota, S.; Yamamoto, S.; Shimomura, A.; Ojida, A.; Nishino, T.; Maruyama, T. Quantification of amino groups on solid surfaces using cleavable fluorescent compounds. Langmuir 2015, 31, 8824–8829. [Google Scholar] [CrossRef]
- Neumann, K.T.; Lindhardt, A.T.; Bang-Andersen, B.; Skrydstrup, T. Synthesis and selective 2H-, 13C-, and 15N-labeling of the Tau protein binder THK-523. J. Label. Compd. Radiopharm. 2017, 60, 30–35. [Google Scholar] [CrossRef]
- Wuest, M.; Trayner, B.J.; Grant, T.N.; Jans, H.S.; Mercer, J.R.; Murray, D.; West, F.G.; McEwan, A.J.B.; Wuest, F.; Cheeseman, C.I. Radiopharmacological evaluation of 6-deoxy-6-[18F] fluoro-D-fructose as a radiotracer for PET imaging of GLUT5 in breast cancer. Nucl. Med. Biol. 2011, 38, 461–475. [Google Scholar] [CrossRef]
- Halgren, T.A.; Murphy, R.B.; Friesner, R.A.; Beard, H.S.; Frye, L.L.; Pollard, W.T.; Banks, J.L. Glide: A new approach for rapid, accurate docking and scoring. 2. Enrichment factors in database screening. J. Med. Chem. 2004, 47, 1750–1759. [Google Scholar] [CrossRef]
- Jo, S.; Kim, T.; Iyer, V.G.; Im, W. CHARMM-GUI: A web-based graphical user interface for CHARMM. J. Comput. Chem. 2008, 29, 1859–1865. [Google Scholar] [CrossRef]
- Roe, D.R.; Cheatham, T.E., III. PTRAJ and CPPTRAJ: Software for processing and analysis of molecular dynamics trajectory data. J. Chem. Theory Comput. 2013, 9, 3084–3095. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 27–38. [Google Scholar] [CrossRef]
- Miller, B.R.; McGee, T.D.; Swails, J.M.; Homeyer, N.; Gohlke, H.; Roitberg, A.E. MMPBSA. py: An efficient program for end-state free energy calculations. J. Chem. Theory Comput. 2012, 8, 3314–3321. [Google Scholar] [CrossRef] [PubMed]
- Kumar Kondapi, V.P.; Soueidan, O.M.; Cheeseman, C.I.; West, F.G. Tunable GLUT–Hexose Binding and Transport via Modulation of Hexose C-3 Hydrogen-Bonding Capabilities. Chem. Eur. J. 2017, 23, 8073–8081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Begoyan, V.V.; Weseliński, Ł.J.; Xia, S.; Fedie, J.; Kannan, S.; Ferrier, A.; Rao, S.; Tanasova, M. Multicolor GLUT5-permeable fluorescent probes for fructose transport analysis. Chem. Commun. 2018, 54, 3855–3858. [Google Scholar] [CrossRef] [PubMed]
- Trayner, B.J.; Grant, T.N.; West, F.G.; Cheeseman, C.I. Synthesis and characterization of 6-deoxy-6-fluoro-D-fructose as a potential compound for imaging breast cancer with PET. Bioorg. Med. Chem. 2009, 17, 5488–5495. [Google Scholar] [CrossRef]
- Thompson, A.M.G.; Iancu, C.V.; Nguyen, T.T.H.; Kim, D.; Choe, J.Y. Inhibition of human GLUT1 and GLUT5 by plant carbohydrate products; insights into transport specificity. Sci. Rep. 2015, 5, 1–10. [Google Scholar] [CrossRef] [Green Version]
- George Thompson, A.M.; Ursu, O.; Babkin, P.; Iancu, C.V.; Whang, A.; Oprea, T.I.; Choe, J.Y. Discovery of a specific inhibitor of human GLUT5 by virtual screening and in vitro transport evaluation. Sci. Rep. 2016, 6, 24240. [Google Scholar] [CrossRef] [Green Version]
- Ainsley, J.; Chaturvedi, S.S.; Karabencheva-Christova, T.G.; Tanasova, M.; Christov, C.Z. Integrating molecular probes and molecular dynamics to reveal binding modes of GLUT5 activatory and inhibitory ligands. Chem. Commun. 2018, 54, 9917–9920. [Google Scholar] [CrossRef]
- Ferreira, R.S.; Pons, J.-L.; Labesse, G. Insights into Substrate and Inhibitor Selectivity among Human GLUT Transporters through Comparative Modeling and Molecular Docking. ACS Omega 2019, 4, 4748–4760. [Google Scholar] [CrossRef]
- Tian, C.; Kasavajhala, K.; Belfon, K.A.A.; Raguette, L.; Huang, H.; Migues, A.N.; Bickel, J.; Wang, Y.; Pincay, J.; Wu, Q.; et al. ff19SB: Amino-acid-specific protein backbone parameters trained against quantum mechanics energy surfaces in solution. J. Chem. Theory Comput. 2019, 16, 528–552. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, W.; Kollman, P.A.; Case, D.A. Automatic atom type and bond type perception in molecular mechanical calculations. J. Mol. Graph. Model. 2006, 25, 247–260. [Google Scholar] [CrossRef]
- Bai, Q.; Zhang, Y.; Li, X.; Chen, W.; Liu, H.; Yao, X. Computational study on the interaction between CCR5 and HIV-1 entry inhibitor maraviroc: Insight from accelerated molecular dynamics simulation and free energy calculation. Phys. Chem. Chem. Phys. 2014, 16, 24332–24338. [Google Scholar] [CrossRef] [PubMed]
- Genheden, S.; Ryde, U. The MM/PBSA and MM/GBSA methods to estimate ligand-binding affinities. Expert Opin. Drug Discov. 2015, 10, 449–461. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | n/x | IC50 (mM) |
|---|---|---|
| 3 | 6/2 | 1.10 ± 0.17 |
| 4 | 6/2 | ~6 (estimated) |
| 5 | 6/2 | n.d. |
| 6 | 9/3 | 2.29 ± 0.18 |
| 7 | 6/2 | n.d. |
| 8 | 6/2 | ~20 (estimated) |
| 9 | 6/2 | n.d. |
| 10 | 6/2 | 2.31 ± 0.35 |
| 11 | 6/2 | 1.71 ± 0.08 |
| 6-FDF | 9/3 | 17.02 ± 0.75 |
| Fructose | 12/4 | 342 ± 74 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rana, N.; Aziz, M.A.; Oraby, A.K.; Wuest, M.; Dufour, J.; Abouzid, K.A.M.; Wuest, F.; West, F.G. Towards Selective Binding to the GLUT5 Transporter: Synthesis, Molecular Dynamics and In Vitro Evaluation of Novel C-3-Modified 2,5-Anhydro-D-mannitol Analogs. Pharmaceutics 2022, 14, 828. https://doi.org/10.3390/pharmaceutics14040828
Rana N, Aziz MA, Oraby AK, Wuest M, Dufour J, Abouzid KAM, Wuest F, West FG. Towards Selective Binding to the GLUT5 Transporter: Synthesis, Molecular Dynamics and In Vitro Evaluation of Novel C-3-Modified 2,5-Anhydro-D-mannitol Analogs. Pharmaceutics. 2022; 14(4):828. https://doi.org/10.3390/pharmaceutics14040828
Chicago/Turabian StyleRana, Natasha, Marwa A. Aziz, Ahmed K. Oraby, Melinda Wuest, Jennifer Dufour, Khaled A. M. Abouzid, Frank Wuest, and F. G. West. 2022. "Towards Selective Binding to the GLUT5 Transporter: Synthesis, Molecular Dynamics and In Vitro Evaluation of Novel C-3-Modified 2,5-Anhydro-D-mannitol Analogs" Pharmaceutics 14, no. 4: 828. https://doi.org/10.3390/pharmaceutics14040828
APA StyleRana, N., Aziz, M. A., Oraby, A. K., Wuest, M., Dufour, J., Abouzid, K. A. M., Wuest, F., & West, F. G. (2022). Towards Selective Binding to the GLUT5 Transporter: Synthesis, Molecular Dynamics and In Vitro Evaluation of Novel C-3-Modified 2,5-Anhydro-D-mannitol Analogs. Pharmaceutics, 14(4), 828. https://doi.org/10.3390/pharmaceutics14040828