
The Anti-Inflammatory Effect of Nanoarchaeosomes on Human Endothelial Cells


Abstract
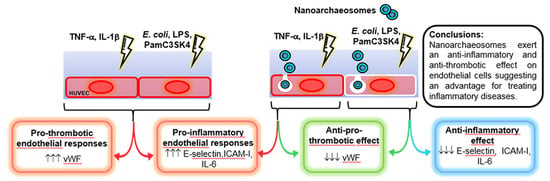
:
1. Introduction
2. Materials and Methods
2.1. Archaebacteria Growth, Extraction & Characterization of Total Polar Archaeolipids
2.2. Preparation of Nanovesicles
2.3. Characterization of Nanovesicles
2.4. Endothelial Cell Culture
2.5. Measurement of Cell Viability and Apoptosis
2.6. EdU Cell Proliferation Assay
2.7. Expression of E-Selectin and ICAM-1
2.8. Measurement of IL-6 and vWF
2.9. ARC Uptake by HUVEC
2.10. Analysis of ERK1/2 and NF-κB Signaling Pathways
2.11. Quantitative PCR (qPCR)
2.12. Statistical Analysis
3. Results
3.1. Structural Characterization
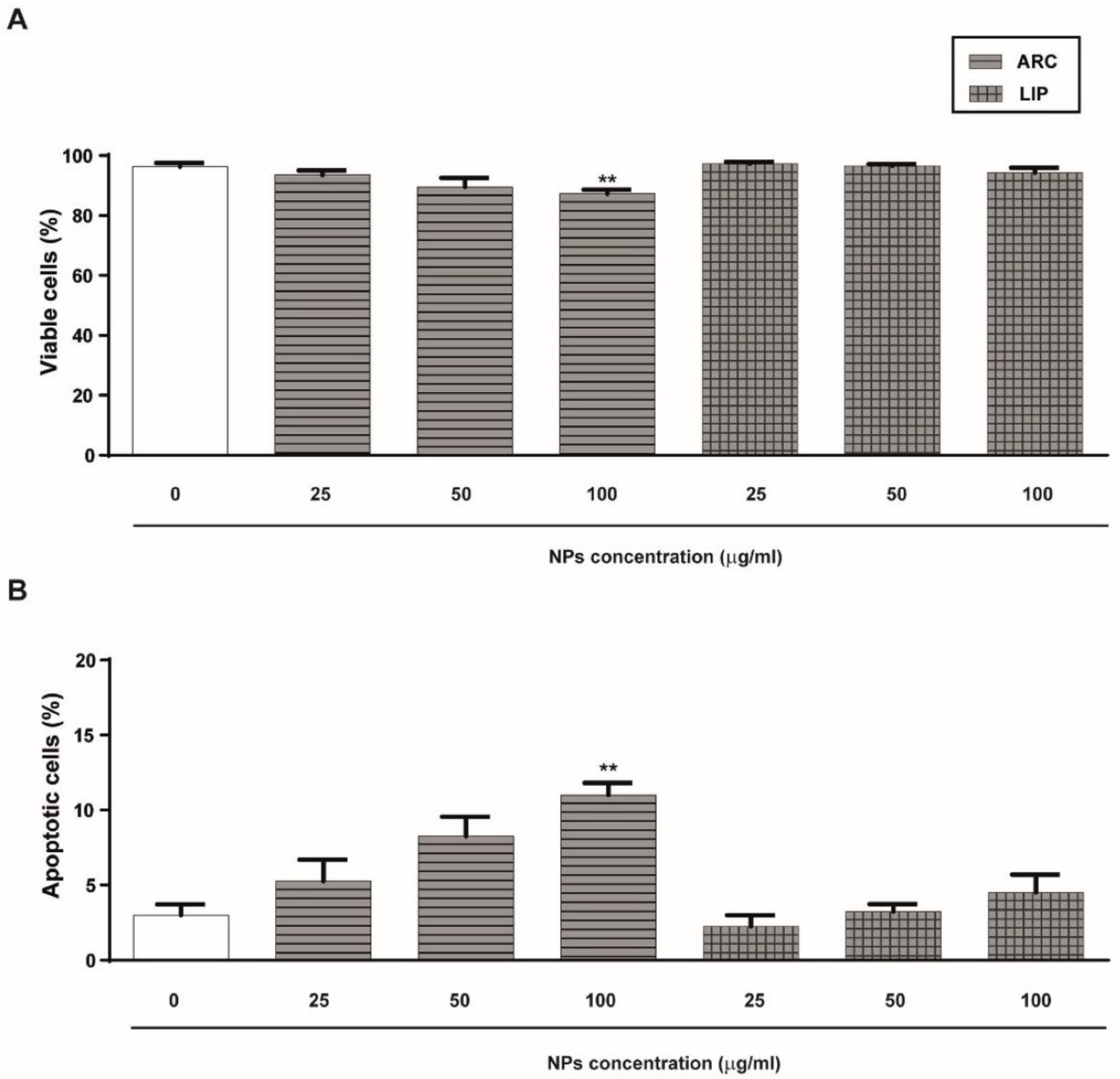
3.2. ARC Do Not Alter HUVEC’s Viability or Proliferation
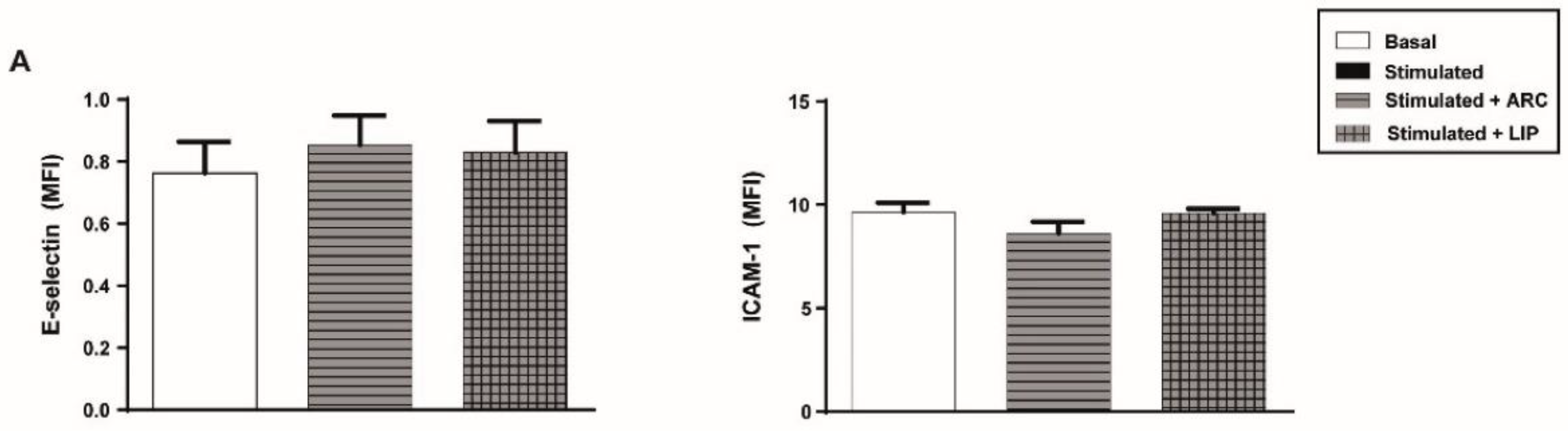
3.3. ARC Selectively Inhibit the Expression of E-Selectin and ICAM-1
3.4. ARC Decrease the Levels of Cytokines Released by Endothelial Cells Stimulated with LPS or Pam3CSK4 but Not with TNF-α or IL-1β
3.5. The Inhibitory Effect of ARC Is Not Due to Their Interaction with LPS, Pam3CSK4, or E. coli or Their Receptors
3.6. ARC Decrease the Levels of vWF Released by Stimulated HUVEC
3.7. ARC Are Internalized by HUVEC
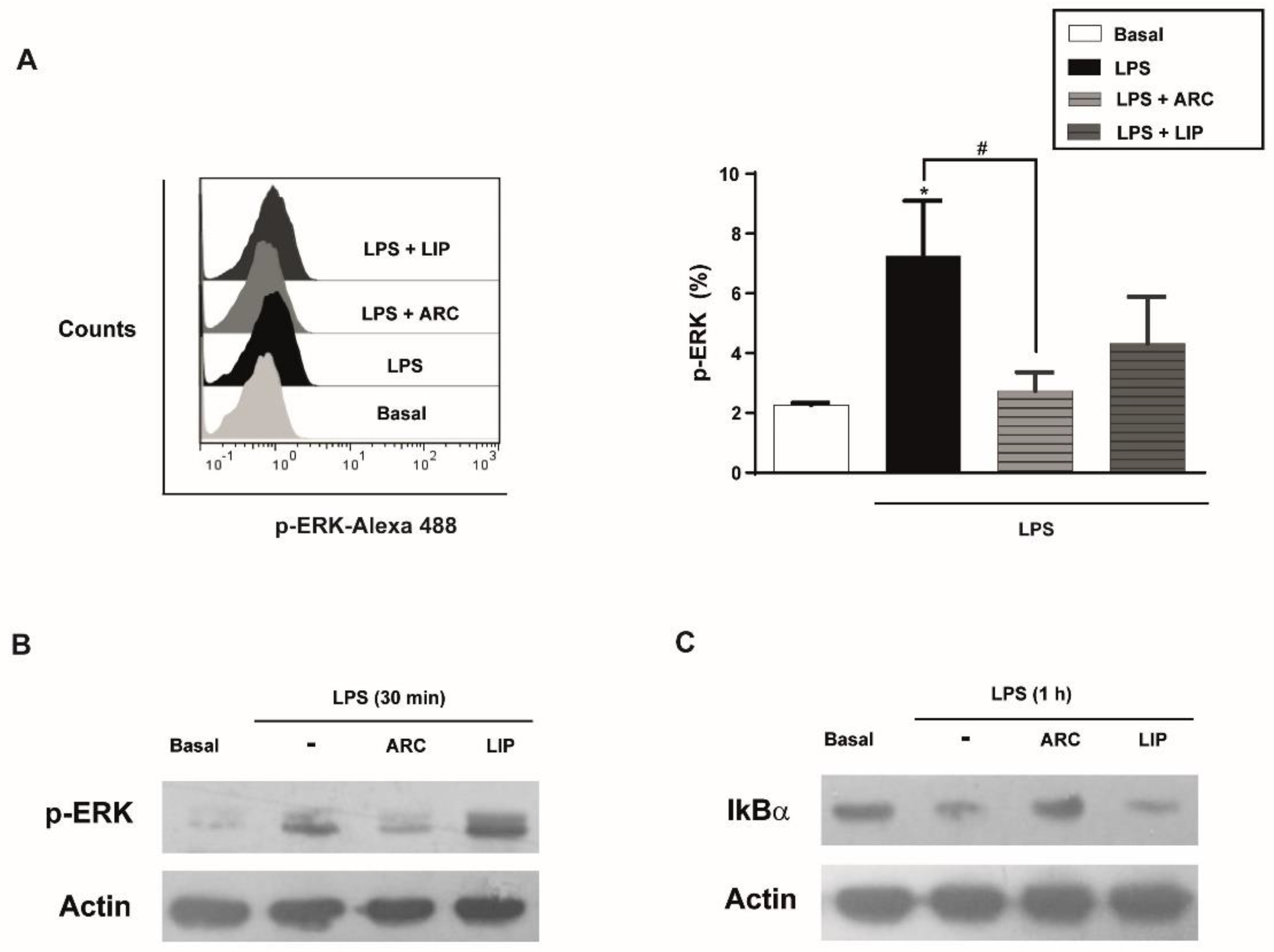
3.8. ARC Attenuate LPS-Induced Activation of NF-κB Pathway
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Younis, M.A.; Tawfeek, H.M.; Abdellatif, A.A.H.; Abdel-Aleem, J.A.; Harashima, H. Clinical Translation of Nanomedicines: Challenges, Opportunities, and Keys. Adv. Drug Deliv. Rev. 2022, 181, 114083. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.; Huang, J.; Moses, M.A. Cancer Nanomedicines in an Evolving Oncology Landscape. Trends Pharmacol. Sci. 2020, 41, 730–742. [Google Scholar] [CrossRef] [PubMed]
- US Food and Drug Administration. FDA Approves First-of-Its Kind Targeted RNA-Based Therapy to Treat a Rare Disease; US Food and Drug Administration: Silver Spring, MD, USA, 2018.
- Suzuki, Y.; Ishihara, H. Difference in the Lipid Nanoparticle Technology Employed in Three Approved SiRNA (Patisiran) and MRNA (COVID-19 Vaccine) Drugs. Drug Metab. Pharmacokinet. 2021, 41, 100424. [Google Scholar] [CrossRef] [PubMed]
- WHO. Cardiovascular Diseases (CVDs); WHO: Geneva, Switzerland, 2019.
- Wang, D.K.; Rahimi, M.; Filgueira, C.S. Nanotechnology Applications for Cardiovascular Disease Treatment: Current and Future Perspectives. Nanomed. Nanotechnol. Biol. Med. 2021, 34, 102387. [Google Scholar] [CrossRef] [PubMed]
- Cicha, I. The Grand Challenges in Cardiovascular Drug Delivery. Front. Drug Deliv. 2021, 1, 1–4. [Google Scholar] [CrossRef]
- Wolinsky, H. A Proposal Linking Clearance of Circulating Lipoproteins to Tissue Metabolic Activity as a Basis for Understanding Atherogenesis. Circ. Res. 1980, 47, 301–311. [Google Scholar] [CrossRef] [Green Version]
- Gimbrone, M.A., Jr.; García-Cardeña, G. Endothelial Cell Dysfunction and the Pathobiology of Atherosclerosis. Circ. Res. 2016, 176, 139–148. [Google Scholar] [CrossRef]
- Liu, X.; Sun, J. Endothelial Cells Dysfunction Induced by Silica Nanoparticles through Oxidative Stress via JNK/P53 and NF-ΚB Pathways. Biomaterials 2010, 31, 8198–8209. [Google Scholar] [CrossRef]
- Tsou, T.C.; Yeh, S.C.; Tsai, F.Y.; Lin, H.J.; Cheng, T.J.; Chao, H.R.; Tai, L.A. Zinc Oxide Particles Induce Inflammatory Responses in Vascular Endothelial Cells via NF-ΚB Signaling. J. Hazard. Mater. 2010, 183, 182–188. [Google Scholar] [CrossRef]
- Shi, J.; Sun, X.; Lin, Y.; Zou, X.; Li, Z.; Liao, Y.; Du, M.; Zhang, H. Endothelial Cell Injury and Dysfunction Induced by Silver Nanoparticles through Oxidative Stress via IKK/NF-ΚB Pathways. Biomaterials 2014, 35, 6657–6666. [Google Scholar] [CrossRef]
- Guo, C.; Xia, Y.; Niu, P.; Jiang, L.; Duan, J.; Yu, Y.; Zhou, X.; Li, Y.; Sun, Z. Silica Nanoparticles Induce Oxidative Stress, Inflammation, and Endothelial Dysfunction in Vitro via Activation of the MAPK/Nrf2 Pathway and Nuclear Factor-ΚB Signaling. Int. J. Nanomed. 2015, 10, 1463–1477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Younis, N.K.; Ghoubaira, J.A.; Bassil, E.P.; Tantawi, H.N.; Eid, A.H. Metal-Based Nanoparticles: Promising Tools for the Management of Cardiovascular Diseases. Nanomed. Nanotechnol. Biol. Med. 2021, 36, 102433. [Google Scholar] [CrossRef] [PubMed]
- Glassman, P.M.; Myerson, J.W.; Ferguson, L.T.; Kiseleva, R.Y.; Shuvaev, V.V.; Brenner, J.S.; Muzykantov, V.R. Targeting Drug Delivery in the Vascular System: Focus on Endothelium. Adv. Drug Deliv. Rev. 2020, 157, 96–117. [Google Scholar] [CrossRef] [PubMed]
- Ding, B.S.; Dziubla, T.; Shuvaev, V.V.; Muro, S.; Muzykantov, V.R. Advanced Drug Delivery Systems That Target the Vascular Endothelium. Mol. Interv. 2006, 6, 98–112. [Google Scholar] [CrossRef] [Green Version]
- Bose, R.J.; Ha, K.; McCarthy, J.R. Bio-Inspired Nanomaterials as Novel Options for the Treatment of Cardiovascular Disease. Drug Discov. Today 2021, 26, 1200–1211. [Google Scholar] [CrossRef]
- Benvegnu, T.; Lemiegre, L.; Cammas-Marion, S. New Generation of Liposomes Called Archaeosomes Based on Natural or Synthetic Archaeal Lipids as Innovative Formulations for Drug Delivery. Recent Pat. Drug Deliv. Formul. 2009, 3, 206–220. [Google Scholar] [CrossRef]
- Gonzalez, R.O.; Higa, L.H.; Cutrullis, R.A.; Bilen, M.; Morelli, I.; Roncaglia, D.I.; Corral, R.S.; Morilla, M.J.; Petray, P.B.; Romero, E.L. Archaeosomes Made of Halorubrum Tebenquichense Total Polar Lipids: A New Source of Adjuvancy. BMC Biotechnol. 2009, 9, 71. [Google Scholar] [CrossRef] [Green Version]
- Higa, L.H.; Corral, R.S.; José Morilla, M.; Romero, E.L.; Petray, P.B. Archaeosomes Display Immunoadjuvant Potential for a Vaccine against Chagas Disease. Hum. Vaccines Immunother. 2013, 9, 409–412. [Google Scholar] [CrossRef] [Green Version]
- Kaur, G.; Garg, T.; Rath, G.; Goyal, A.K. Archaeosomes: An Excellent Carrier for Drug and Cell Delivery. Drug Deliv. 2016, 23, 2497–2512. [Google Scholar] [CrossRef]
- Moghimipour, E.; Kargar, M.; Handali, S. Archaeosomes as Means of Nano-Drug Delivery. Rev. Med. Microbiol. 2014, 25, 40–45. [Google Scholar] [CrossRef]
- Parra, F.L.; Caimi, A.T.; Altube, M.J.; Cargnelutti, D.E.; Vermeulen, M.E.; De Farias, M.A.; Morilla, M.J.; Romero, E.L. Make It Simple: (SR-A1 + TLR7) Macrophage Targeted NANOarchaeosomes. Front. Bioeng. Biotechnol. 2018, 6, 163. [Google Scholar] [CrossRef] [PubMed]
- Kate, M. Chapter 9 Membrane Lipids of Archaea. New Compr. Biochem. 1993, 26, 261–295. [Google Scholar] [CrossRef]
- Schilrreff, P.; Simioni, Y.R.; Jerez, H.E.; Caimi, A.T.; de Farias, M.A.; Villares Portugal, R.; Romero, E.L.; Morilla, M.J. Superoxide Dismutase in Nanoarchaeosomes for Targeted Delivery to Inflammatory Macrophages. Colloids Surf. B Biointerfaces 2019, 179, 479–487. [Google Scholar] [CrossRef] [PubMed]
- Caimi, A.T.; Parra, F.; de Farias, M.A.; Portugal, R.V.; Perez, A.P.; Romero, E.L.; Morilla, M.J. Topical Vaccination with Super-Stable Ready to Use Nanovesicles. Colloids Surf. B Biointerfaces 2017, 152, 114–123. [Google Scholar] [CrossRef]
- Altube, M.J.; Selzer, S.M.; De Farias, M.A.; Portugal, R.V.; Morilla, M.J.; Romero, E.L. Surviving Nebulization-Induced Stress: Dexamethasone in PH-Sensitive Archaeosomes. Nanomedicine 2016, 11, 2103–2117. [Google Scholar] [CrossRef]
- Omri, A.; Agnew, B.J.; Patel, G.B. Short-Term Repeated-Dose Toxicity Profile of Archaeosomes Administered to Mice via Intravenous and Oral Routes. Int. J. Toxicol. 2003, 22, 9–23. [Google Scholar] [CrossRef]
- Julia Altube, M.; Ivonne Caimi, L.; Huck-Iriart, C.; Jose Morilla, M.; Lilia Romero, E.; Rosaria Lauro, M. Pharmaceutics Reparation of an Inflamed Air-Liquid Interface Cultured A549 Cells with Nebulized Nanocurcumin. Pharmaceutics 2021, 13, 1331. [Google Scholar] [CrossRef]
- Higa, L.H.; Jerez, H.E.; De Farias, M.A.; Portugal, R.V.; Romero, E.L.; Morilla, M.J. Ultra-Small Solid Archaeolipid Nanoparticles for Active Targeting to Macrophages of the Inflamed Mucosa. Nanomedicine 2017, 12, 1165–1175. [Google Scholar] [CrossRef]
- Higa, L.H.; Schilrreff, P.; Perez, A.P.; Iriarte, M.A.; Roncaglia, D.I.; Morilla, M.J.; Romero, E.L. Ultradeformable Archaeosomes as New Topical Adjuvants. Nanomed. Nanotechnol. Biol. Med. 2012, 8, 1319–1328. [Google Scholar] [CrossRef]
- Kates, M.; Kushwaha, S.C. Isoprenoids and Polar Lipids of Extreme Halophiles. In Archaea, a Laboratory Manual: Halophiles; DasSarma, S., Fleischmann, E.M., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1995; pp. 35–54. [Google Scholar]
- Böttcher, C.J.F.; Van gent, C.M.; Pries, C. A Rapid and Sensitive Sub-Micro Phosphorus Determination. Anal. Chim. Acta 1961, 24, 203–204. [Google Scholar] [CrossRef]
- Altube, M.J.; Cutro, A.; Bakas, L.; Morilla, M.J.; Disalvo, E.A.; Romero, E.L. Nebulizing Novel Multifunctional Nanovesicles: The Impact of Macrophage-Targeted-PH-Sensitive Archaeosomes on a Pulmonary Surfactant. J. Mater. Chem. B 2017, 5, 8083–8095. [Google Scholar] [CrossRef]
- Jaffe, E.A.; Nachman, R.L.; Becker, C.G.; Minick, C.R. Culture of Human Endothelial Cells Derived from Umbilical Veins. Identification by Morphologic and Immunologic Criteria. J. Clin. Investig. 1973, 52, 2745–2756. [Google Scholar] [CrossRef] [PubMed]
- Negrotto, S.; Malaver, E.; Alvarez, M.E.; Pacienza, N.; D’Atri, L.P.; Pozner, R.G.; Gómez, R.M.; Schattner, M. Aspirin and Salicylate Suppress Polymorphonuclear Apoptosis Delay Mediated by Proinflammatory Stimuli. J. Pharmacol. Exp. Ther. 2006, 319, 972–979. [Google Scholar] [CrossRef] [PubMed]
- Scholz, D.; Devaux, B.; Hirche, A.; Pötzsch, B.; Kropp, B.; Schaper, W.; Schaper, J. Expression of Adhesion Molecules Is Specific and Time-Dependent in Cytokine-Stimulated Endothelial Cells in Culture. Cell Tissue Res. 1996, 284, 415–423. [Google Scholar] [CrossRef] [PubMed]
- Boyle, E.M.; Sato, T.T.; Noel, R.F.; Verrier, E.D.; Pohlman, T.H. Transcriptional Arrest of the Human E-Selectin Gene. J. Surg. Res. 1999, 82, 194–200. [Google Scholar] [CrossRef]
- Carestia, A.; Kaufman, T.; Rivadeneyra, L.; Landoni, V.I.; Pozner, R.G.; Negrotto, S.; D’Atri, L.P.; Gómez, R.M.; Schattner, M. Mediators and Molecular Pathways Involved in the Regulation of Neutrophil Extracellular Trap Formation Mediated by Activated Platelets. J. Leukoc. Biol. 2016, 99, 153–162. [Google Scholar] [CrossRef]
- Nightingale, T.; Cutler, D. The Secretion of von Willebrand Factor from Endothelial Cells; an Increasingly Complicated Story. J. Thromb. Haemost. 2013, 11, 192–201. [Google Scholar] [CrossRef]
- Joyce, D.E.; Grinnell, B.W. Recombinant Human Activated Protein C Attenuates the Inflammatory Response in Endothelium and Monocytes by Modulating Nuclear Factor-ΚB. Crit. Care Med. 2002, 30, S288–S293. [Google Scholar] [CrossRef]
- Berdiaki, A.; Perisynaki, E.; Stratidakis, A.; Kulikov, P.P.; Kuskov, A.N.; Stivaktakis, P.; Henrich-Noack, P.; Luss, A.L.; Shtilman, M.M.; Tzanakakis, G.N.; et al. Assessment of Amphiphilic Poly- N-Vinylpyrrolidone Nanoparticles’ Biocompatibility with Endothelial Cells in Vitro and Delivery of an Anti-Inflammatory Drug. Mol. Pharm. 2020, 17, 4212–4225. [Google Scholar] [CrossRef]
- Beconcini, D.; Felice, F.; Zambito, Y.; Fabiano, A.; Piras, A.M.; Macedo, M.H.; Sarmento, B.; Di Stefano, R. Anti-Inflammatory Effect of Cherry Extract Loaded in Polymeric Nanoparticles: Relevance of Particle Internalization in Endothelial Cells. Pharmaceutics 2019, 11, 500. [Google Scholar] [CrossRef] [Green Version]
- Subash-Babu, P.; Al-Saran, N.; Alshammari, G.M.; Naif Al-Harbi, L.; Hussain Alhussain, M.; Shamlan, G.; Abdulaziz AlSedairy, S.; Abdullah Alshatwi, A.; Tsim, K.; Alshammari, M.G.; et al. Evaluation of Biosafety, Antiobesity, and Endothelial Cells Proliferation Potential of Basil Seed Extract Loaded Organic Solid Lipid Nanoparticle. Front. Pharmacol. 2021, 12, 2419. [Google Scholar] [CrossRef] [PubMed]
- Corbalan, J.J.; Medina, C.; Jacoby, A.; Malinski, T.; Radomski, M.W. Amorphous Silica Nanoparticles Trigger Nitric Oxide/Peroxynitrite Imbalance in Human Endothelial Cells: Inflammatory and Cytotoxic Effects. Int. J. Nanomed. 2011, 6, 2821–2835. [Google Scholar] [CrossRef] [Green Version]
- Nagarajan, M.; Maadurshni, G.B.; Tharani, G.K.; Udhayakumar, I.; Kumar, G.; Mani, K.P.; Sivasubramanian, J.; Manivannan, J. Exposure to Zinc Oxide Nanoparticles (ZnO-NPs) Induces Cardiovascular Toxicity and Exacerbates Pathogenesis–Role of Oxidative Stress and MAPK Signaling. Chem. Biol. Interact. 2022, 351, 109719. [Google Scholar] [CrossRef] [PubMed]
- Maschera, B.; Ray, K.; Burns, K.; Volpe, F. Overexpression of an Enzymically Inactive Interleukin-1-Receptor-Associated Kinase Activates Nuclear Factor-ΚB. Biochem. J. 1999, 339, 227–231. [Google Scholar] [CrossRef]
- Zhang, F.X.; Kirschning, C.J.; Mancinelli, R.; Xu, X.P.; Jin, Y.; Faure, E.; Mantovani, A.; Rothe, M.; Muzio, M.; Arditi, M. Bacterial Lipopolysaccharide Activates Nuclear Factor-ΚB through Interleukin-1 Signaling Mediators in Cultured Human Dermal Endothelial Cells and Mononuclear Phagocytes. J. Biol. Chem. 1999, 274, 7611–7614. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Ghosh, S. Toll-like Receptor-Mediated NF-ΚB Activation: A Phylogenetically Conserved Paradigm in Innate Immunity. J. Clin. Investig. 2001, 107, 13–19. [Google Scholar] [CrossRef] [Green Version]
- Martin, M.U.; Wesche, H. Summary and Comparison of the Signaling Mechanisms of the Toll/Interleukin-1 Receptor Family. Biochim. Biophys. Acta-Mol. Cell Res. 2002, 1592, 265–280. [Google Scholar] [CrossRef] [Green Version]
- Bochkov, V.N.; Kadl, A.; Huber, J.; Gruber, F.; Binder, B.R.; Leitinger, N. Protective Role of Phospholipid Oxidation Products in Endotoxin-Induced Tissue Damage. Nature 2002, 419, 77–81. [Google Scholar] [CrossRef]
- Wu, A.; Hinds, C.J.; Thiemermann, C. High-Density Lipoproteins in Sepsis and Septic Shock: Metabolism, Actions, and Therapeutic Applications. Shock 2004, 21, 210–221. [Google Scholar] [CrossRef]
- Parker, T.S.; Levine, D.M.; Chang, J.C.C.; Laxer, J.; Coffin, C.C.; Rubin, A.L. Reconstituted High-Density Lipoprotein Neutralizes Gram-Negative Bacterial Lipopolysaccharides in Human Whole Blood. Infect. Immun. 1995, 63, 253–258. [Google Scholar] [CrossRef] [Green Version]
- Gupta, H.; Dai, L.; Datta, G.; Garber, D.W.; Grenett, H.; Li, Y.; Mishra, V.; Palgunachari, M.N.; Handattu, S.; Gianturco, S.H.; et al. Inhibition of Lipopolysaccharide-Induced Inflammatory Responses by an Apolipoprotein AI Mimetic Peptide. Circ. Res. 2005, 97, 236–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Triantafilou, M.; Miyake, K.; Golenbock, D.T.; Triantafilou, K. Mediators of Innate Immune Recognition of Bacteria Concentrate in Lipid Rafts and Facilitate Lipopolysaccharide-Induced Cell Activation. J. Cell Sci. 2002, 115, 2603–2611. [Google Scholar] [CrossRef] [PubMed]
- Triantafilou, M.; Gamper, F.G.J.; Haston, R.M.; Mouratis, M.A.; Morath, S.; Hartung, T.; Triantafilou, K. Membrane Sorting of Toll-like Receptor (TLR)-2/6 and TLR2/1 Heterodimers at the Cell Surface Determines Heterotypic Associations with CD36 and Intracellular Targeting. J. Biol. Chem. 2006, 281, 31002–31011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGettrick, A.F.; Brint, E.K.; Palsson-McDermott, E.M.; Rowe, D.C.; Golenbock, D.T.; Gay, N.J.; Fitzgerald, K.A.; O’Neill, L.A.J. Trif-Related Adapter Molecule Is Phosphorylated by PKCε during Toll-like Receptor 4 Signaling. Proc. Natl. Acad. Sci. USA 2006, 103, 9196–9201. [Google Scholar] [CrossRef] [Green Version]
- Abate, W.; Alghaithy, A.A.; Parton, J.; Jones, K.P.; Jackson, S.K. Surfactant Lipids Regulate LPS-Induced Interleukin-8 Production in A549 Lung Epithelial Cells by Inhibiting Translocation of TLR4 into Lipid Raft Domains. J. Lipid Res. 2010, 51, 334–344. [Google Scholar] [CrossRef] [Green Version]
- Walton, K.A.; Hsieh, X.; Gharavi, N.; Wang, S.; Wang, G.; Yeh, M.; Cole, A.L.; Berliner, J.A. Receptors Involved in the Oxidized 1-Palmitoyl-2-Arachidonoyl-Sn-Glycero-3-Phosphorylcholine-Mediated Synthesis of Interleukin-8: A Role for Toll-like Receptor 4 and a Glycosylphosphatidylinositol-Anchored Protein. J. Biol. Chem. 2003, 278, 29661–29666. [Google Scholar] [CrossRef] [Green Version]
- Yamauchi, K.; Doi, K.; Yoshida, Y.; Kinoshita, M. Archaebacterial Lipids: Highly Proton-Impermeable Membranes from 1,2-Diphytanyl-Sn-Glycero-3-Phosphocoline. BBA-Biomembranes 1993, 1146, 178–182. [Google Scholar] [CrossRef]
- Róg, T.; Vattulainen, I.; Bunker, A.; Karttunen, M. Glycolipid Membranes through Atomistic Simulations: Effect of Glucose and Galactose Head Groups on Lipid Bilayer Properties. J. Phys. Chem. B 2007, 111, 10146–10154. [Google Scholar] [CrossRef]
- Kitano, T.; Onoue, T.; Yamauchi, K. Archaeal Lipids Forming a Low Energy-Surface on Air-Water Interface. Chem. Phys. Lipids 2003, 126, 225–232. [Google Scholar] [CrossRef]
- Chollet, J.L.; Jozwiakowski, M.J.; Phares, K.R.; Reiter, M.J.; Roddy, P.J.; Schultz, H.J.; Ta, Q.V.; Tomai, M.A. Development of a Topically Active Imiquimod Formulation. Pharm. Dev. Technol. 1999, 4, 35–43. [Google Scholar] [CrossRef]
- Carrer, D.C.; Higa, L.H.; Tesoriero, M.V.D.; Morilla, M.J.; Roncaglia, D.I.; Romero, E.L. Structural Features of Ultradeformable Archaeosomes for Topical Delivery of Ovalbumin. Colloids Surf. B Biointerfaces 2014, 121, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Mosior, M.; Newton, A.C. Mechanism of Interaction of Protein Kinase C with Phorbol Esters. J. Biol. Chem. 1995, 270, 25526–25533. [Google Scholar] [CrossRef] [Green Version]
- Galbiati, F.; Razani, B.; Lisanti, M.P. Emerging Themes in Lipid Rafts and Caveolae. Cell 2001, 106, 403–411. [Google Scholar] [CrossRef] [Green Version]
- Oakley, F.O.; Smith, R.L.; Engelhardt, J.F. Lipid Rafts and Caveolin-1 Coordinate Interleukin-1β (IL-1β)-Dependent Activation of NFκB by Controlling Endocytosis of Nox2 and IL-1β Receptor 1 from the Plasma Membrane. J. Biol. Chem. 2009, 284, 33255–33264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doan, J.E.S.; Windmiller, D.A.; Riches, D.W.H. Differential Regulation of TNF-R1 Signaling: Lipid Raft Dependency of P42 Mapk/Erk2 Activation, but Not NF-ΚB Activation. J. Immunol. 2004, 172, 7654–7660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, T.; Read, M.A.; Neish, A.S.; Whitley, M.Z.; Thanos, D.; Maniatis, T. Transcriptional Regulation of Endothelial Cell Adhesion Molecules: NF-κB and Cytokine-inducible Enhancers. FASEB J. 1995, 9, 899–909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bürgin-Maunder, C.S.; Brooks, P.R.; Russell, F.D. Omega-3 Fatty Acids Modulate Weibel-Palade Body Degranulation and Actin Cytoskeleton Rearrangement in PMA-Stimulated Human Umbilical Vein Endothelial Cells. Mar. Drugs 2013, 11, 4435–4450. [Google Scholar] [CrossRef] [Green Version]
- El-Mansi, S.; Nightingale, T.D. Emerging Mechanisms to Modulate VWF Release from Endothelial Cells. Int. J. Biochem. Cell Biol. 2021, 131, 105900. [Google Scholar] [CrossRef]
- Springer, T.A.; Cybulsky, M.I. Traffic Signals on Endothelium for Leukocytes in Health, Inflammation, and Atherosclerosis. In Atherosclerosis and Coronary Artery Disease; Lippincott-Raven Publishers: Philadelphia, PA, USA, 1995; pp. 511–537. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Formulation | Size (nm ± SD) | PDI | ζ Potential (mV ± SD) | TL (mg/mL ± SD) | RhPE (μg/mL ± SD) | RhPE/TL (μg/mg ± SD) |
|---|---|---|---|---|---|---|
| ARC | 168.9 ± 11 | 0.19 ± 0.01 | −41.5 ± 4.5 | 7.5 ± 1.3 | - | - |
| LIP | 276.7 ± 11 | 0.30 ± 0.08 | −5.4 ± 1.4 | 7.0 ± 1.7 | - | - |
| ARC-RhPE | 159.7 ± 7.6 | 0.19 ± 0.02 | −37.0 ± 4.0 | 6.9 ± 0.6 | 3.2 ± 0.6 | 0.48 ± 0.09 |
| LIP-RhPE | 251.1 ± 9.1 | 0.29 ± 0.07 | −3.7 ± 0.6 | 6.0 ± 0.3 | 2.8 ± 0.9 | 0.45 ± 0.12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Charó, N.; Jerez, H.; Tatti, S.; Romero, E.L.; Schattner, M. The Anti-Inflammatory Effect of Nanoarchaeosomes on Human Endothelial Cells. Pharmaceutics 2022, 14, 736. https://doi.org/10.3390/pharmaceutics14040736
Charó N, Jerez H, Tatti S, Romero EL, Schattner M. The Anti-Inflammatory Effect of Nanoarchaeosomes on Human Endothelial Cells. Pharmaceutics. 2022; 14(4):736. https://doi.org/10.3390/pharmaceutics14040736
Chicago/Turabian StyleCharó, Nancy, Horacio Jerez, Silvio Tatti, Eder Lilia Romero, and Mirta Schattner. 2022. "The Anti-Inflammatory Effect of Nanoarchaeosomes on Human Endothelial Cells" Pharmaceutics 14, no. 4: 736. https://doi.org/10.3390/pharmaceutics14040736
APA StyleCharó, N., Jerez, H., Tatti, S., Romero, E. L., & Schattner, M. (2022). The Anti-Inflammatory Effect of Nanoarchaeosomes on Human Endothelial Cells. Pharmaceutics, 14(4), 736. https://doi.org/10.3390/pharmaceutics14040736