
Phages against Pathogenic Bacterial Biofilms and Biofilm-Based Infections: A Review



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. The Bacterial Biofilm
2.1. The Composition and Architecture of the Bacterial Biofilm
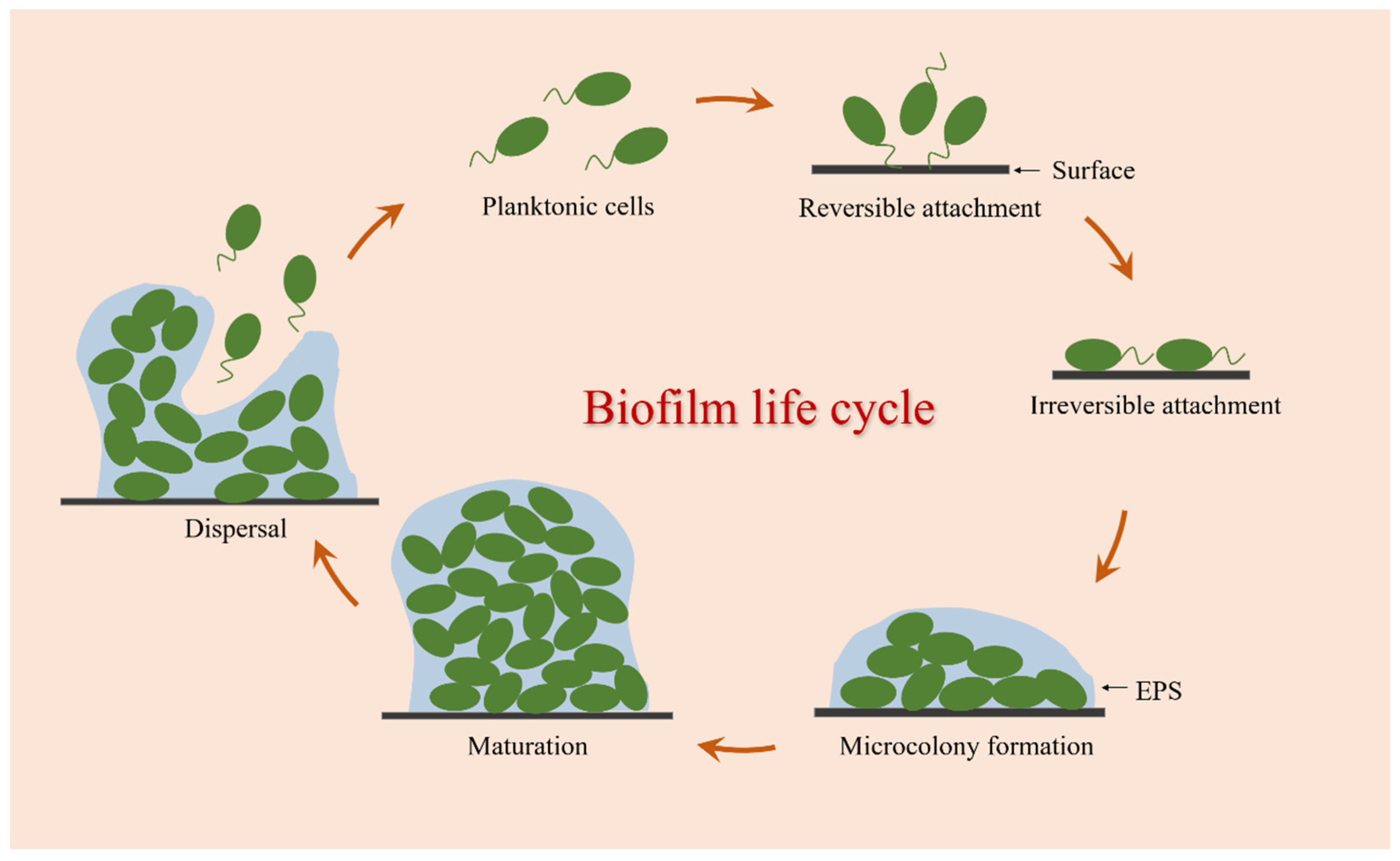
2.2. The Formation of Bacterial Biofilm
3. Interactions between Phages and Bacterial Biofilms
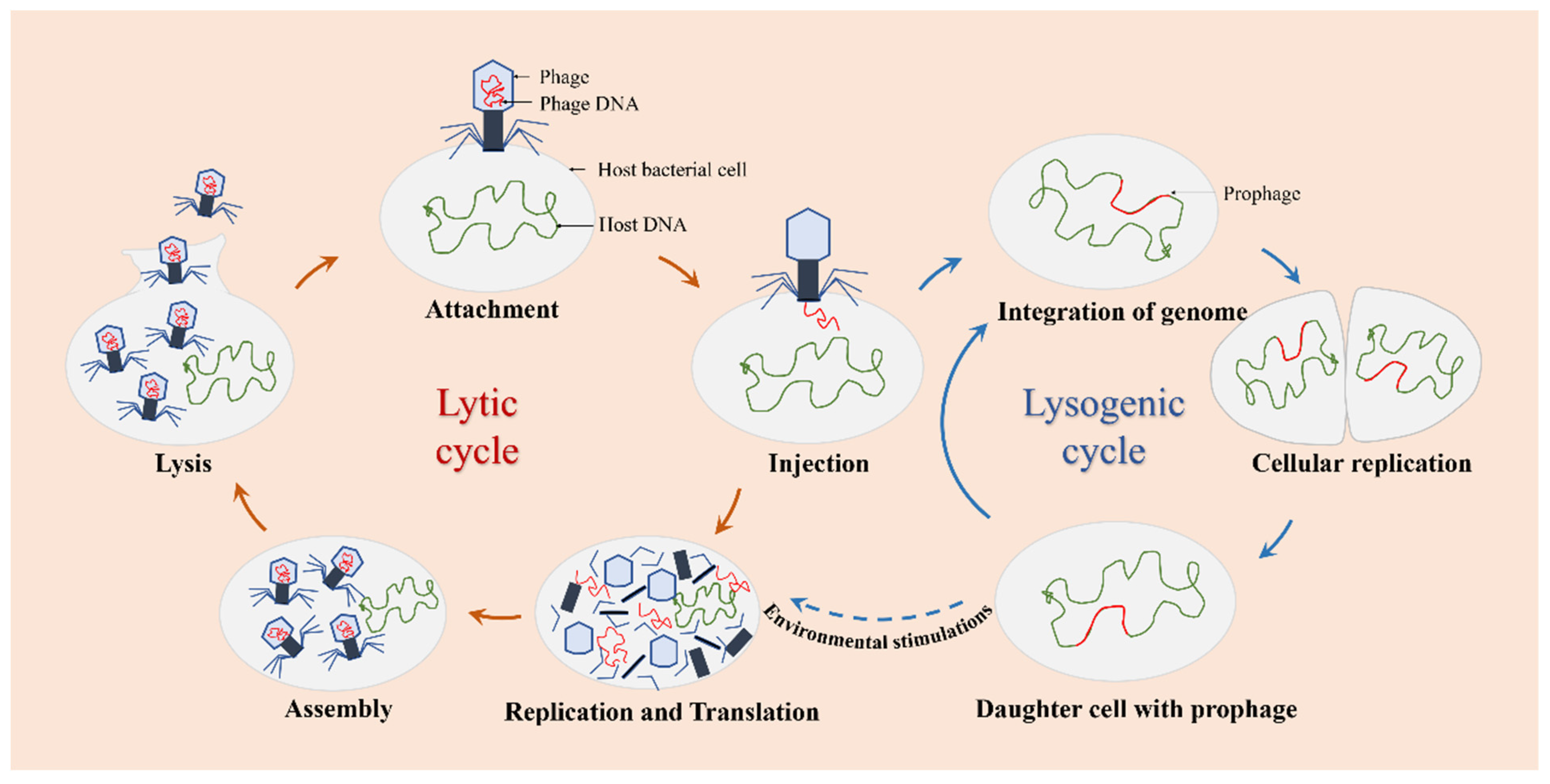
3.1. Phages Infect Bacterial Biofilms
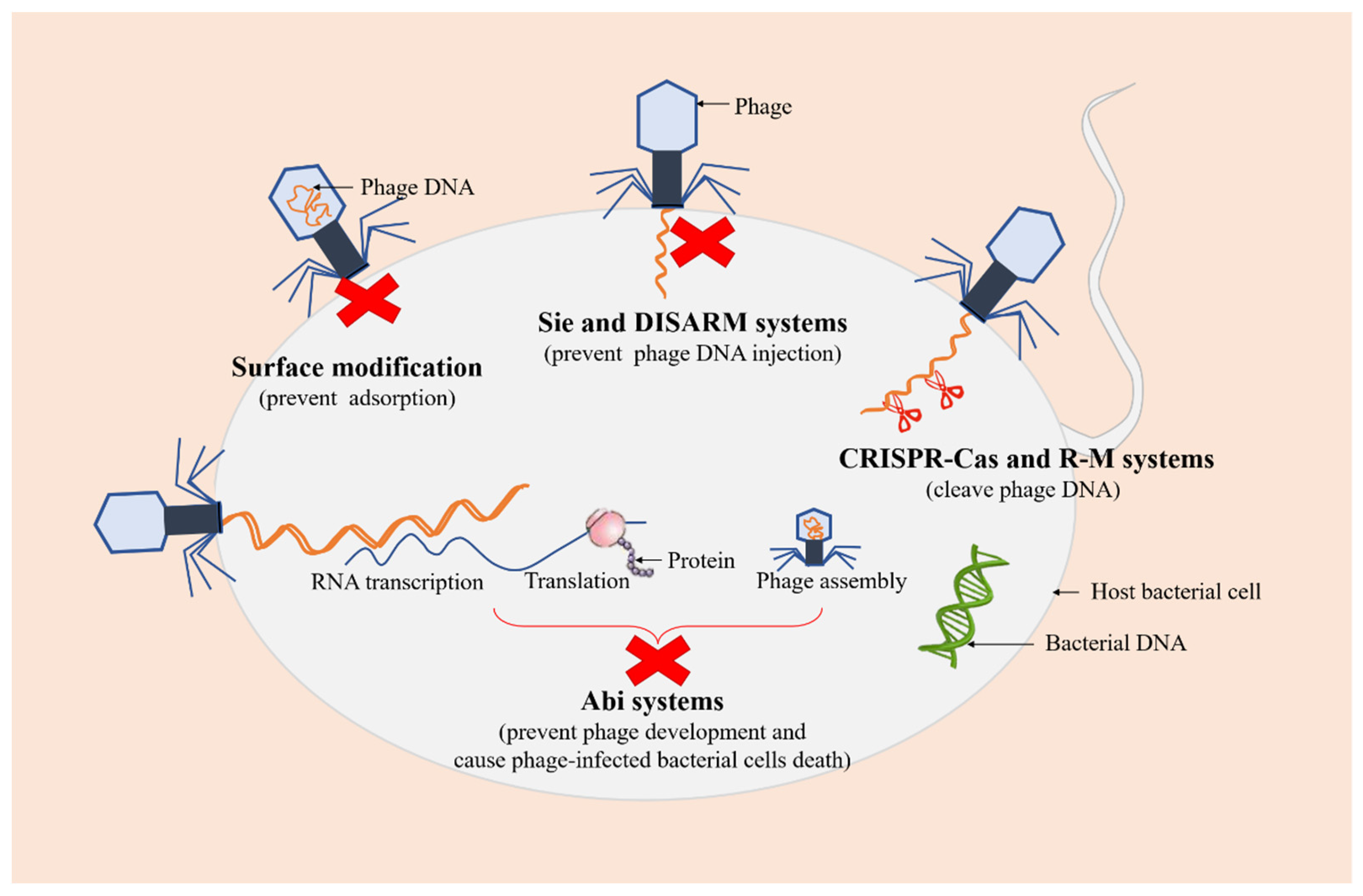
3.2. Bacterial Biofilms Resist Phage Infections
4. Phage-Based Strategies for Preventing and Controlling Pathogenic Bacteria Biofilms
4.1. Phage Cocktail Therapy
4.2. The Combination of Phage with Antibiotics
4.3. Genetically Engineered Phages
4.4. Phage-Derived Enzymes
4.5. The Combination of Phage with Other Strategies
5. Conclusions and Perspectives
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Khan, F.; Pham, D.T.N.; Oloketuyi, S.F.; Kim, Y.-M. Antibiotics application strategies to control biofilm formation in pathogenic bacteria. Curr. Pharm. Biotechnol. 2020, 21, 270–286. [Google Scholar] [CrossRef] [PubMed]
- Amankwah, S.; Abdella, K.; Kassa, T. Bacterial biofilm destruction: A focused review on the recent use of phage-based strategies with other antibiofilm agents. Nanotechnol. Sci. Appl. 2021, 14, 161–177. [Google Scholar] [CrossRef] [PubMed]
- Jamal, M.; Ahmad, W.; Andleeb, S.; Jalil, F.; Imran, M.; Nawaz, M.A.; Hussain, T.; Ali, M.; Rafiq, M.; Kamil, M.A. Bacterial biofilm and associated infections. J. Chin. Med. Assoc. 2018, 81, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Vishwakarma, A.; Dang, F.; Ferrell, A.; Barton, H.A.; Joy, A. Peptidomimetic polyurethanes inhibit bacterial biofilm formation and disrupt surface established biofilms. J. Am. Chem. Soc. 2021, 143, 9440–9449. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; van der Mei, H.C.; Busscher, H.J.; Ren, Y. Enhanced bacterial killing by vancomycin in staphylococcal biofilms disrupted by novel, DMMA-modified carbon dots depends on EPS production. Colloids Surf. B-Biointerfaces 2020, 193, 111114. [Google Scholar] [CrossRef] [PubMed]
- Larsen, J.; Raisen, C.L.; Ba, X.; Sadgrove, N.J.; Padilla-Gonzalez, G.F.; Simmonds, M.S.J.; Loncaric, I.; Kerschner, H.; Apfalter, P.; Hartl, R.; et al. Emergence of methicillin resistance predates the clinical use of antibiotics. Nature 2022, 602, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Szafranski, S.P.; Slots, J.; Stiesch, M. The human oral phageome. Periodontology 2000 2021, 86, 79–96. [Google Scholar] [CrossRef]
- Lin, Y.W.; Chang, R.Y.; Rao, G.G.; Jermain, B.; Han, M.L.; Zhao, J.X.; Chen, K.; Wang, J.P.; Barr, J.J.; Schooley, R.T.; et al. Pharmacokinetics/pharmacodynamics of antipseudomonal bacteriophage therapy in rats: A proof-of-concept study. Clin. Microbiol. Infect. 2020, 26, 1229–1235. [Google Scholar] [CrossRef]
- Reuter, M.; Kruger, D.H. Approaches to optimize therapeutic bacteriophage and bacteriophage-derived products to combat bacterial infections. Virus Genes 2020, 56, 136–149. [Google Scholar] [CrossRef]
- Guo, D.M.; Chen, J.C.; Zhao, X.Y.; Luo, Y.N.; Jin, M.L.; Fan, F.X.; Park, C.; Yang, X.M.; Sun, C.Q.; Yan, J.; et al. Genetic and cemical engineering of phages for controlling multidrug-resistant bacteria. Antibiotics 2021, 10, 202. [Google Scholar] [CrossRef]
- Uruen, C.; Chopo-Escuin, G.; Tommassen, J.; Mainar-Jaime, R.C.; Arenas, J. Biofilms as promoters of bacterial antibiotic resistance and tolerance. Antibiotics 2021, 10, 3. [Google Scholar] [CrossRef] [PubMed]
- Chou, K.-T.; Lee, D.-Y.D.; Chiou, J.-G.; Galera-Laporta, L.; Ly, S.; Garcia-Ojalvo, J.; Suel, G.M. A segmentation clock patterns cellular differentiation in a bacterial biofilm. Cell 2022, 185, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Flemming, H.C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Karygianni, L.; Ren, Z.; Koo, H.; Thurnheer, T. Biofilm matrixome: Extracellular components in structured microbial communities. Trends Microbiol. 2020, 28, 668–681. [Google Scholar] [CrossRef] [PubMed]
- Siddik, A.; Satheesh, S. Characterization and assessment of barnacle larval settlement-inducing activity of extracellular polymeric substances isolated from marine biofilm bacteria. Sci. Rep. 2019, 9, 17849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Low, K.E.; Howell, P.L. Gram-negative synthase-dependent exopolysaccharide biosynthetic machines. Curr. Opin. Struct. Biol. 2018, 53, 32–44. [Google Scholar] [CrossRef] [PubMed]
- Steiger, E.L.; Muelli, J.R.; Braissant, O.; Waltimo, T.; Astasov-Frauenhoffer, M. Effect of divalent ions on cariogenic biofilm formation. BMC Microbiol. 2020, 20, 287. [Google Scholar] [CrossRef]
- Devaraj, A.; Novotny, L.A.; Robledo-Avila, F.H.; Buzzo, J.R.; Mashburn-Warren, L.; Jurcisek, J.A.; Tjokro, N.O.; Partida-Sanchez, S.; Bakaletz, L.O.; Goodman, S.D. The extracellular innate-immune effector HMGB1 limits pathogenic bacterial biofilm proliferation. J. Clin. Investig. 2021, 131, e140527. [Google Scholar] [CrossRef]
- Paharik, A.E.; Horswill, A.R. The staphylococcal biofilm: Adhesins, regulation, and host response. Microbiol. Spectr. 2016, 4, 529–566. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, E.; Greene, N.P.; Crow, A.; Koronakis, V. Insights into bacterial lipoprotein trafficking from a structure of LolA bound to the LolC periplasmic domain. Proc. Natl. Acad. Sci. USA 2018, 115, E7389–E7397. [Google Scholar] [CrossRef] [Green Version]
- Campoccia, D.; Montanaro, L.; Arciola, C.R. Extracellular DNA (eDNA). A Major Ubiquitous Element of the Bacterial Biofilm Architecture. Int. J. Mol. Sci. 2021, 22, 9100. [Google Scholar] [CrossRef] [PubMed]
- Kavanaugh, J.S.; Flack, C.E.; Lister, J.; Ricker, E.B.; Ibberson, C.B.; Jenul, C.; Moormeier, D.E.; Delmain, E.A.; Bayles, K.W.; Horswill, A.R. Identification of Extracellular DNA-Binding Proteins in the Biofilm Matrix. Mbio 2019, 10, e01137-19. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Liu, Q.; Li, X.; Ma, S.; Hu, H.; Wu, B.; Zhang, X.-x.; Ren, H. In-situ monitoring AHL-mediated quorum-sensing regulation of the initial phase of wastewater biofilm formation. Environ. Int. 2020, 135, 105326. [Google Scholar] [CrossRef] [PubMed]
- Renner, L.D.; Weibel, D.B. Physicochemical regulation of biofilm formation. MRS Bull. 2011, 36, 347–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berne, C.; Brun, Y.V. The two chemotaxis clusters in Caulobacter crescentus play different roles in chemotaxis and biofilm regulation. J. Bacteriol. 2019, 201, e00071-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toyofuku, M.; Inaba, T.; Kiyokawa, T.; Obana, N.; Yawata, Y.; Nomura, N. Environmental factors that shape biofilm formation. Biosci. Biotechnol. Biochem. 2016, 80, 7–12. [Google Scholar] [CrossRef] [PubMed]
- McCall, A.D.; Kumar, R.; Edgerton, M. Candida albicans Sfl1/Sfl2 regulatory network drives the formation of pathogenic microcolonies. PLoS Pathog. 2018, 14, e1007316. [Google Scholar] [CrossRef]
- Nassar, R.; Hachim, M.; Nassar, M.; Kaklamanos, E.G.; Jamal, M.; Williams, D.; Senok, A. Microbial metabolic genes crucial for S. aureus biofilms: An insight from re-analysis of publicly available microarray datasets. Front. Microbiol. 2021, 11, 3598. [Google Scholar] [CrossRef]
- Nishikawa, M.; Kobayashi, K. Calcium prevents biofilm dispersion in Bacillus subtilis. J. Bacteriol. 2021, 203, e00114-21. [Google Scholar] [CrossRef]
- Wang, T.; Huang, W.; Duan, Q.; Wang, J.; Cheng, H.; Shao, J.; Li, F.; Wu, D. Sodium houttuyfonate in vitro inhibits biofilm dispersion and expression of bdlA in Pseudomonas aeruginosa. Mol. Biol. Rep. 2019, 46, 471–477. [Google Scholar] [CrossRef]
- Batinovic, S.; Wassef, F.; Knowler, S.A.; Rice, D.T.F.; Stanton, C.R.; Rose, J.; Tucci, J.; Nittami, T.; Vinh, A.; Drummond, G.R.; et al. Bacteriophages in natural and artificial environments. Pathogens 2019, 8, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nair, A.; Ghugare, G.S.; Khairnar, K. An appraisal of bacteriophage isolation techniques from environment. Microb. Ecol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Vlassov, V.V.; Tikunova, N.V.; Morozova, V.V. Bacteriophages as therapeutic preparations: What restricts their application in medicine. Biochem.-Mosc. 2020, 85, 1350–1361. [Google Scholar] [CrossRef]
- Salmond, G.P.C.; Fineran, P.C. A century of the phage: Past, present and future. Nat. Rev. Microbiol. 2015, 13, 777–786. [Google Scholar] [CrossRef] [PubMed]
- Giri, N. Bacteriophage structure, classification, assembly and phage therapy. Biosci. Biotechnol. Res. Asia 2021, 18, 239–250. [Google Scholar] [CrossRef]
- Ramirez-Chamorro, L.; Boulanger, P.; Rossier, O. Strategies for bacteriophage T5 mutagenesis: Expanding the toolbox for phage genome engineering. Front. Microbiol. 2021, 12, 816. [Google Scholar] [CrossRef]
- Maghsoodi, A.; Chatterjee, A.; Andricioaei, I.; Perkins, N.C. How the phage T4 injection machinery works including energetics, forces, and dynamic pathway. Proc. Natl. Acad. Sci. USA 2019, 116, 25097–25105. [Google Scholar] [CrossRef]
- Swanson, N.A.; Lokareddy, R.K.; Li, F.; Hou, C.-F.; Pavlenok, M.; Niederweis, M.; Cingolani, G. Expression and purification of phage T7 ejection proteins for cryo-EM analysis. STAR Protoc. 2021, 2, 100960. [Google Scholar] [CrossRef]
- Ashy, R.A.; Agusti, S. Low host abundance and high temperature determine switching from lytic to lysogenic cycles in planktonic microbial communities in a Tropical Sea (Red Sea). Viruses 2020, 12, 761. [Google Scholar] [CrossRef]
- Wang, Q.; Guan, Z.Y.; Pei, K.; Wang, J.; Liu, Z.; Yin, P.; Peng, D.H.; Zou, T.T. Structural basis of the arbitrium peptide-AimR communication system in the phage lysis-lysogeny decision. Nat. Microbiol. 2018, 3, 1266–1273. [Google Scholar] [CrossRef]
- Elahi, Y.; Nowroozi, J.; Fard, R.M.N. Isolation and characterization of bacteriophages from wastewater sources on Enterococcus spp. isolated from clinical samples. Iran. J. Microbiol. 2021, 13, 671–677. [Google Scholar] [CrossRef] [PubMed]
- Pires, D.P.; Oliveira, H.; Melo, L.D.R.; Sillankorva, S.; Azeredo, J. Bacteriophage-encoded depolymerases: Their diversity and biotechnological applications. Appl. Microbiol. Biotechnol. 2016, 100, 2141–2151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lubowska, N.; Piechowicz, L. Staphylococcus aureus biofilm and the role of bacteriophages in its eradication. Postepy Hig. I Med. Dosw. 2018, 72, 101–107. [Google Scholar] [CrossRef]
- Topka-Bielecka, G.; Dydecka, A.; Necel, A.; Bloch, S.; Nejman-Falenczyk, B.; Wegrzyn, G.; Wegrzyn, A. Bacteriophage-derived depolymerases against bacterial biofilm. Antibiotics 2021, 10, 175. [Google Scholar] [CrossRef]
- Vazquez, R.; Garcia, P. Synergy between two chimeric lysins to kill Streptococcus pneumoniae. Front. Microbiol. 2019, 10, 1251. [Google Scholar] [CrossRef] [Green Version]
- Castillo, D.; Rorbo, N.; Jorgensen, J.; Lange, J.; Tan, D.; Kalatzis, P.G.; Lo Svenningsen, S.; Middelboe, M. Phage defense mechanisms and their genomic and phenotypic implications in the fish pathogen Vibrio anguillarum. FEMS Microbiol. Ecol. 2019, 95, fiz004. [Google Scholar] [CrossRef]
- Reyes-Robles, T.; Dillard, R.S.; Cairns, L.S.; Silva-Valenzuela, C.A.; Housman, M.; Ali, A.; Wright, E.R.; Camilli, A. Vibrio cholerae outer membrane vesicles inhibit bacteriophage infection. J. Bacteriol. 2018, 200, e00792-17. [Google Scholar] [CrossRef] [Green Version]
- Harvey, H.; Bondy-Denomy, J.; Marquis, H.; Sztanko, K.M.; Davidson, A.R.; Burrows, L.L. Pseudomonas aeruginosa defends against phages through type IV pilus glycosylation. Nat. Microbiol. 2018, 3, 47–52. [Google Scholar] [CrossRef]
- Biggs, K.R.H.; Bailes, C.L.; Scott, L.; Wichman, H.A.; Schwartz, E.J. Ecological approach to understanding superinfection inhibition in bacteriophage. Viruses 2021, 13, 1389. [Google Scholar] [CrossRef]
- Shi, K.; Oakland, J.T.; Kurniawan, F.; Moeller, N.H.; Banerjee, S.; Aihara, H. Structural basis of superinfection exclusion by bacteriophage T4 Spackle. Commun. Biol. 2020, 3, 691. [Google Scholar] [CrossRef]
- Ofir, G.; Melamed, S.; Sberro, H.; Mukamel, Z.; Silverman, S.; Yaakov, G.; Doron, S.; Sorek, R. DISARM is a widespread bacterial defence system with broad anti-phage activities. Nat. Microbiol. 2018, 3, 90–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Dai, L.; Dong, J.; Chen, C.; Zhu, J.; Rao, V.B.; Tao, P. Covalent modifications of the bacteriophage genome confer a degree of resistance to bacterial CRISPR systems. J. Virol. 2020, 94, e01630-20. [Google Scholar] [CrossRef] [PubMed]
- Roy, D.; Huguet, K.T.; Grenier, F.; Burrus, V. IncC conjugative plasmids and SXT/R391 elements repair double-strand breaks caused by CRISPR-Cas during conjugation. Nucleic Acids Res. 2020, 48, 8815–8827. [Google Scholar] [CrossRef] [PubMed]
- Wilkowska, K.; Mruk, I.; Furmanek-Blaszk, B.; Sektas, M. Low-level expression of the Type II restriction-modification system confers potent bacteriophage resistance in Escherichia coli. DNA Res. 2020, 27, dsaa003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smirnov, S.V.; Morozova, N.E.; Khodorkovskii, M.A.; Severinov, K.V. Fluorescence microscopy study of the effect of Esp1396I restriction-modification system proteins concentrations on protection against lambda phage. In Proceedings of the International Conference on PhysicA.SPb, Saint Petersburg, Russia, 23–25 October 2018. [Google Scholar]
- Mekler, V.; Kuznedelov, K.; Minakhin, L.; Murugan, K.; Sashital, D.G.; Severinov, K. CRISPR-Cas molecular beacons as tool for studies of assembly of CRISPR-Cas effector complexes and their interactions with DNA. Cris.-Cas Enzym. 2019, 616, 337–363. [Google Scholar]
- Borges, A.L.; Castro, B.; Govindarajan, S.; Solvik, T.; Escalante, V.; Bondy-Denomy, J. Bacterial alginate regulators and phage homologs repress CRISPR-Cas immunity. Nat. Microbiol. 2020, 5, 679–687. [Google Scholar] [CrossRef]
- Fineran, P.C. Resistance is not futile: Bacterial ‘innate’ and CRISPR-Cas ‘adaptive’ immune systems. Microbiology 2019, 165, 834–841. [Google Scholar] [CrossRef]
- Lopatina, A.; Tal, N.; Sorek, R. Abortive infection: Bacterial suicide as an antiviral immune strategy. Annu. Rev. Virol. 2020, 7, 371–384. [Google Scholar] [CrossRef]
- Feyereisen, M.; Mahony, J.; O’Sullivan, T.; Boer, V.; van Sinderen, D. Identification of a prophage-encoded abortive infection system in Levilactobacillus brevis. Korean J. Microbiol. Biotechnol. 2020, 48, 322–327. [Google Scholar] [CrossRef]
- Trotter, A.J.; Aydin, A.; Strinden, M.J.; O’Grady, J. Recent and emerging technologies for the rapid diagnosis of infection and antimicrobial resistance. Curr. Opin. Microbiol. 2019, 51, 39–45. [Google Scholar] [CrossRef]
- Udaondo, Z.; Matilla, M.A. Mining for novel antibiotics in the age of antimicrobial resistance. Microb. Biotechnol. 2020, 13, 1702–1704. [Google Scholar] [CrossRef] [PubMed]
- Montso, P.K.; Mlambo, V.; Ateba, C.N. Efficacy of novel phages for control of multi-drug resistant Escherichia coli O177 on artificially contaminated beef and their potential to disrupt biofilm formation. Food Microbiol. 2021, 94, 103647. [Google Scholar] [CrossRef] [PubMed]
- Maszewska, A.; Zygmunt, M.; Grzejdziak, I.; Rozalski, A. Use of polyvalent bacteriophages to combat biofilm of Proteus mirabilis causing catheter-associated urinary tract infections. J. Appl. Microbiol. 2018, 125, 1253–1265. [Google Scholar] [CrossRef] [PubMed]
- Malik, S.; Nehra, K.; Rana, J.S. Bacteriophage cocktail and phage antibiotic synergism as promising alternatives to conventional antibiotics for the control of multi-drug-resistant uropathogenic Escherichia coli. Virus Res. 2021, 302, 198496. [Google Scholar] [CrossRef] [PubMed]
- Zurabov, F.; Zhilenkov, E. Characterization of four virulent Klebsiella pneumoniae bacteriophages, and evaluation of their potential use in complex phage preparation. Virol. J. 2021, 18, 9. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Wang, S.; Guo, Z.; Liu, H.; Sun, D.; Yan, G.; Hu, D.; Du, C.; Feng, X.; Han, W.; et al. A guard-killer phage cocktail effectively lyses the host and inhibits the development of phage-resistant strains of Escherichia coli. Appl. Microbiol. Biotechnol. 2018, 102, 971–983. [Google Scholar] [CrossRef]
- Oechslin, F.; Piccardi, P.; Mancini, S.; Gabard, J.; Moreillon, P.; Entenza, J.M.; Resch, G.; Que, Y.A. Synergistic interaction between phage therapy and antibiotics clears Pseudomonas Aeruginosa infection in endocarditis and reduces virulence. J. Infect. Dis. 2017, 215, 703–712. [Google Scholar] [CrossRef] [Green Version]
- Townsend, E.M.; Moat, J.; Jameson, E. CAUTI’s next top model—Model dependent Klebsiella biofilm inhibition by bacteriophages and antimicrobials. Biofilm 2020, 2, 100038. [Google Scholar] [CrossRef]
- Chaudhry, W.N.; Concepcion-Acevedo, J.; Park, T.; Andleeb, S.; Bull, J.J.; Levin, B.R. Synergy and order effects of antibiotics and phages in killing Pseudomonas aeruginosa biofilms. PLoS ONE 2017, 12, e0168615. [Google Scholar] [CrossRef]
- Kumaran, D.; Taha, M.; Yi, Q.L.; Ramirez-Arcos, S.; Diallo, J.S.; Carli, A.; Abdelbary, H. Does treatment order matter? Investigating the ability of bacteriophage to augment antibiotic activity against Staphylococcus aureus biofilms. Front. Microbiol. 2018, 9, 127. [Google Scholar] [CrossRef] [Green Version]
- Gibb, B.; Hyman, P.; Schneider, C. The many applications of engineered bacteriophages-an overview. Pharmaceuticals 2021, 14, 634. [Google Scholar] [CrossRef]
- Huss, P.; Raman, S. Engineered bacteriophages as programmable biocontrol agents. Curr. Opin. Biotechnol. 2020, 61, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Shi, D.L.; Li, Y.X.; Xiao, Y.Y.; Chen, M.M.; Chen, L.; Du, H.; Zhang, W. Recombination of T4-like phages and its activity against pathogenic Escherichia coli in planktonic and biofilm forms. Virol. Sin. 2020, 35, 651–661. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.K.; Collins, J.J. Dispersing biofilms with engineered enzymatic bacteriophage. Proc. Natl. Acad. Sci. USA 2007, 104, 11197–11202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dedrick, R.M.; Guerrero-Bustamante, C.A.; Garlena, R.A.; Russell, D.A.; Ford, K.; Harris, K.; Gilmour, K.C.; Soothill, J.; Jacobs-Sera, D.; Schooley, R.T.; et al. Engineered bacteriophages for treatment of a patient with a disseminated drug-resistant Mycobacterium abscessus. Nat. Med. 2019, 25, 730–733. [Google Scholar] [CrossRef] [PubMed]
- Born, Y.; Fieseler, L.; Thony, V.; Leimer, N.; Duffy, B.; Loessner, M.J. Engineering of bacteriophages Y2::dpoL1-C and Y2::luxAB for efficient control and rapid detection of the fire blight pathogen, Erwinia amylovora. Appl. Environ. Microbiol. 2017, 83, e00341-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azeredo, J.; Garcia, P.; Drulis-Kawa, Z. Targeting biofilms using phages and their enzymes. Curr. Opin. Biotechnol. 2021, 68, 251–261. [Google Scholar] [CrossRef]
- Fernandez, L.; Gutierrez, D.; Garcia, P.; Rodriguez, A. The perfect bacteriophage for therapeutic applications-a quick guide. Antibiotics 2019, 8, 126. [Google Scholar] [CrossRef] [Green Version]
- Park, D.W.; Park, J.H. Characterization of a novel phage depolymerase specific to Escherichia coli O157:H7 and biofilm control on abiotic surfaces. J. Microbiol. 2021, 59, 1002–1009. [Google Scholar] [CrossRef]
- Olszak, T.; Shneider, M.M.; Latka, A.; Maciejewska, B.; Browning, C.; Sycheva, L.V.; Cornelissen, A.; Danis-Wlodarczyk, K.; Senchenkova, S.N.; Shashkov, A.S.; et al. The O-specific polysaccharide lyase from the phage LKA1 tailspike reduces Pseudomonas virulence. Sci. Rep. 2017, 7, 16302. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.Q.; Wang, R.; Xu, M.S.; Liu, Y.N.; Zhu, X.C.; Qiu, J.F.; Liu, Q.M.; He, P.; Li, Q.T. A novel polysaccharide depolymerase encoded by the phage SH-KP152226 confers specific activity against multidrug-resistant Klebsiella pneumoniae via biofilm degradation. Front. Microbiol. 2019, 10, 2768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Li, P.; Chen, L.; Guo, G.L.; Xiao, Y.Y.; Chen, L.; Du, H.; Zhang, W. Identification of a phage-derived depolymerase specific for KL64 capsule of Klebsiella pneumoniae and its anti-biofilm effect. Virus Genes 2021, 57, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Lusiak-Szelachowska, M.; Weber-Dabrowska, B.; Zaczek, M.; Gorski, A. Anti-biofilm activity of bacteriophages and lysins in chronic rhinosinusitis. Acta Virol. 2021, 65, 127–140. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Liu, M.; Zhang, P.F.; Leung, S.S.Y.; Xia, J. Membrane-permeable antibacterial enzyme against multidrug-resistant Acinetobacter baumannii. Acs Infect. Dis. 2021, 7, 2192–2204. [Google Scholar] [CrossRef] [PubMed]
- Landlinger, C.; Tisakova, L.; Oberbauer, V.; Schwebs, T.; Muhammad, A.; Latka, A.; Van Simaey, L.; Vaneechoutte, M.; Guschin, A.; Resch, G.; et al. Engineered phage endolysin eliminates Gardnerella biofilm without damaging beneficial bacteria in bacterial vaginosis ex vivo. Pathogens 2021, 10, 54. [Google Scholar] [CrossRef]
- Letrado, P.; Corsini, B.; Diez-Martinez, R.; Bustamante, N.; Yuste, J.E.; Garcia, P. Bactericidal synergism between antibiotics and phage endolysin Cpl-711 to kill multidrug-resistant pneumococcus. Future Microbiol. 2018, 13, 1215–1223. [Google Scholar] [CrossRef] [Green Version]
- Olsen, N.M.C.; Thiran, E.; Hasler, T.; Vanzieleghem, T.; Belibasakis, G.N.; Mahillon, J.; Loessner, M.J.; Schmelcher, M. Synergistic removal of static and dynamic Staphylococcus aureus biofilms by combined treatment with a bacteriophage endolysin and a polysaccharide depolymerase. Viruses 2018, 10, 438. [Google Scholar] [CrossRef] [Green Version]
- Remmington, A.; Turner, C.E. The DNases of pathogenic Lancefield streptococci. Microbiol.-Sgm 2018, 164, 242–250. [Google Scholar] [CrossRef]
- Pei, R.; Lamas-Samanamud, G.R. Inhibition of biofilm formation by T7 bacteriophages producing quorum-quenching enzymes. Appl. Environ. Microbiol. 2014, 80, 5340–5348. [Google Scholar] [CrossRef] [Green Version]
- Stachler, E.; Kull, A.; Julian, T.R. Bacteriophage treatment before chemical disinfection can enhance removal of plastic-surface-associated Pseudomonas aeruginosa. Appl. Environ. Microbiol. 2021, 87, e00980-21. [Google Scholar] [CrossRef]
- He, X.W.; Yang, Y.J.; Guo, Y.C.; Lu, S.G.; Du, Y.; Li, J.J.; Zhang, X.P.; Leung, N.L.C.; Zhao, Z.; Niu, G.L.; et al. Phage-guided targeting, discriminative imaging, and synergistic killing of bacteria by AIE bioconjugates. J. Am. Chem. Soc. 2020, 142, 3959–3969. [Google Scholar] [CrossRef] [PubMed]
- Seethalakshmi, P.S.; Rajeev, R.; Kiran, G.S.; Selvin, J. Shrimp disease management for sustainable aquaculture: Innovations from nanotechnology and biotechnology. Aquac. Int. 2021, 29, 1591–1620. [Google Scholar] [CrossRef]
- Bone, S.; Alum, A.; Markovski, J.; Hristovski, K.; Bar-Zeev, E.; Kaufman, Y.; Abbaszadegan, M.; Perreault, F. Physisorption and chemisorption of T4 bacteriophages on amino functionalized silica particles. J. Colloid Interface Sci. 2018, 532, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.; Wang, Z.; Marcos-Hernandez, M.; Zuo, P.; Zhang, D.; Powell, C.; Pan, A.Y.; Villagrn, D.; Wong, M.S.; Alvarez, P.J.J. Bottom-up biofilm eradication using bacteriophage-loaded magnetic nanocomposites: A computational and experimental study. Environ. Sci.-Nano 2019, 6, 3539–3550. [Google Scholar] [CrossRef]
- Li, L.L.; Yu, P.F.; Wang, X.F.; Yu, S.S.; Mathieu, J.; Yu, H.Q.; Alvarez, P.J.J. Enhanced biofilm penetration for microbial control by polyvalent phages conjugated with magnetic colloidal nanoparticle clusters (CNCs). Environ. Sci.-Nano 2017, 4, 1817–1826. [Google Scholar] [CrossRef]
- Han, X.; Lv, P.; Wang, L.-G.; Long, F.; Ma, X.-L.; Liu, C.; Feng, Y.-J.; Yang, M.-F.; Xiao, X. Impact of nano-TiO2 on horizontal transfer of resistance genes mediated by filamentous phage transduction. Environ. Sci.-Nano 2020, 7, 1214–1224. [Google Scholar] [CrossRef]
- Williams, K.P.; Branda, S.; Krishnakumar, R. Therapeutic Phages and methods Thereof. U.S. Patent 11,066,691, 20 July 2021. [Google Scholar]
- Da Costa Garcia, M.A.; Sousa De Sao Jose, C.J.; Rodrigues Leandro, C.I.; Dias Antunes Marcal Da Silva, F.M.R.P.; Ferreira Llorente Grancho Lourenco, S. Antibacterial Phage, Phage Peptides and Methods of Use Thereof. U.S. Patent 09,682,110, 20 June 2017. [Google Scholar]
- Fischetti, V.; Loomis, L. Use of Bacterial Phage Associated Lysing Enzymes for Treating Various Illnesses. U.S. Patent 6,238,661, 29 May 2001. [Google Scholar]
- Fischetti, V.A.; Loeffler, J.M.; Nelson, D. Phage-Associated Lytic Enzymes For Treatment of Streptococcus Pneumoniae and Related Conditions. U.S. Patent 07,569,223, 4 August 2009. [Google Scholar]
- Belcher, A.M.; Tsedev, U.; Lam, F. Homogeneous Engineered Phage Populations. U.S. Patent 10,987,388, 27 April 2021. [Google Scholar]
- Bruessow, H.; Chennoufi, S.; Sidoti, J.; Bruttin, A. Isolated Phages and Their Use in Food or Pet Food Products. U.S. Patent 8,021,657, 2011. [Google Scholar]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.; Lu, H.; Zhang, S.; Shi, Y.; Chen, Q. Phages against Pathogenic Bacterial Biofilms and Biofilm-Based Infections: A Review. Pharmaceutics 2022, 14, 427. https://doi.org/10.3390/pharmaceutics14020427
Liu S, Lu H, Zhang S, Shi Y, Chen Q. Phages against Pathogenic Bacterial Biofilms and Biofilm-Based Infections: A Review. Pharmaceutics. 2022; 14(2):427. https://doi.org/10.3390/pharmaceutics14020427
Chicago/Turabian StyleLiu, Siyu, Hongyun Lu, Shengliang Zhang, Ying Shi, and Qihe Chen. 2022. "Phages against Pathogenic Bacterial Biofilms and Biofilm-Based Infections: A Review" Pharmaceutics 14, no. 2: 427. https://doi.org/10.3390/pharmaceutics14020427
APA StyleLiu, S., Lu, H., Zhang, S., Shi, Y., & Chen, Q. (2022). Phages against Pathogenic Bacterial Biofilms and Biofilm-Based Infections: A Review. Pharmaceutics, 14(2), 427. https://doi.org/10.3390/pharmaceutics14020427