
Discovery and Characterization of a New Crustin Antimicrobial Peptide from Amphibalanus amphitrite

 ,
,  ,
,  and
and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Prediction and Identification of Crustin Sequences
2.3. Expression and Purification of Recombinant AaCrus1 (rAaCrus1)
2.4. Identify rAaCrus1 by Liquid Chromatograph Mass Spectrometer (LC-MS)
2.5. Antimicrobial Activity of the rAaCrus1
2.6. Molecular Dynamics (MD) Simulations
2.7. Microorganism-Binding Assay
2.8. Agglutinating Assay
2.9. Electron Microscopy
2.10. PI Staining Assay
2.11. Biofilm Inhibition
2.12. Statistical Analysis
3. Results
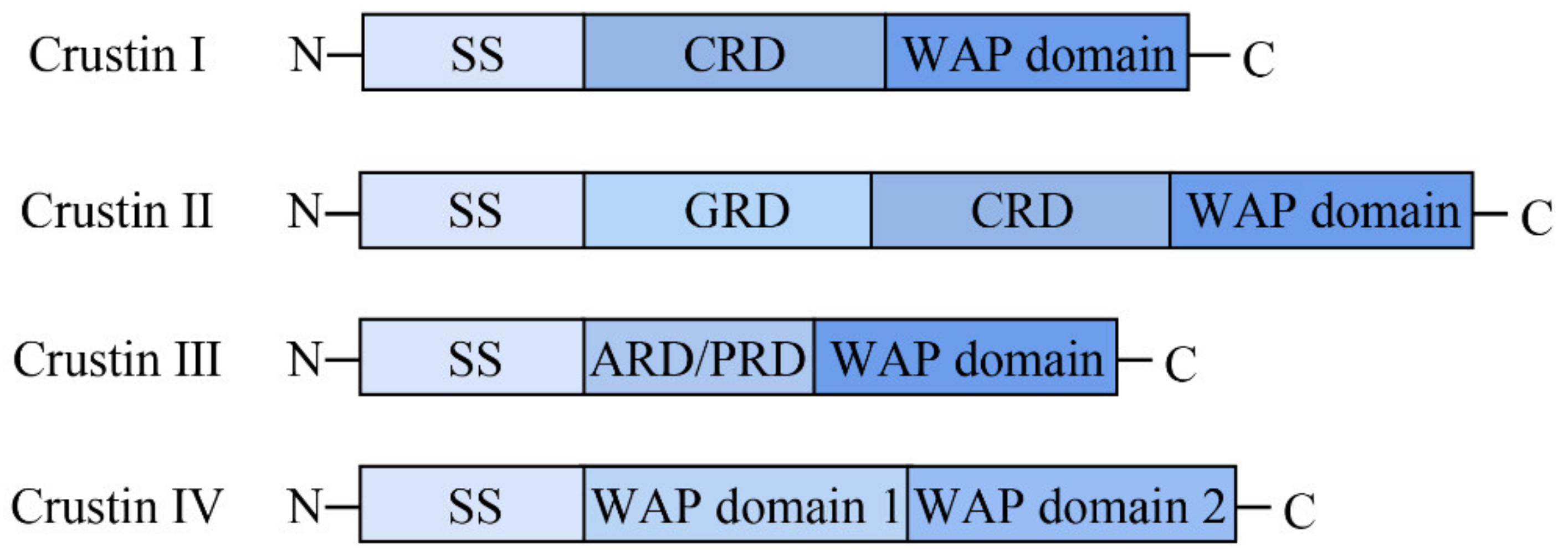
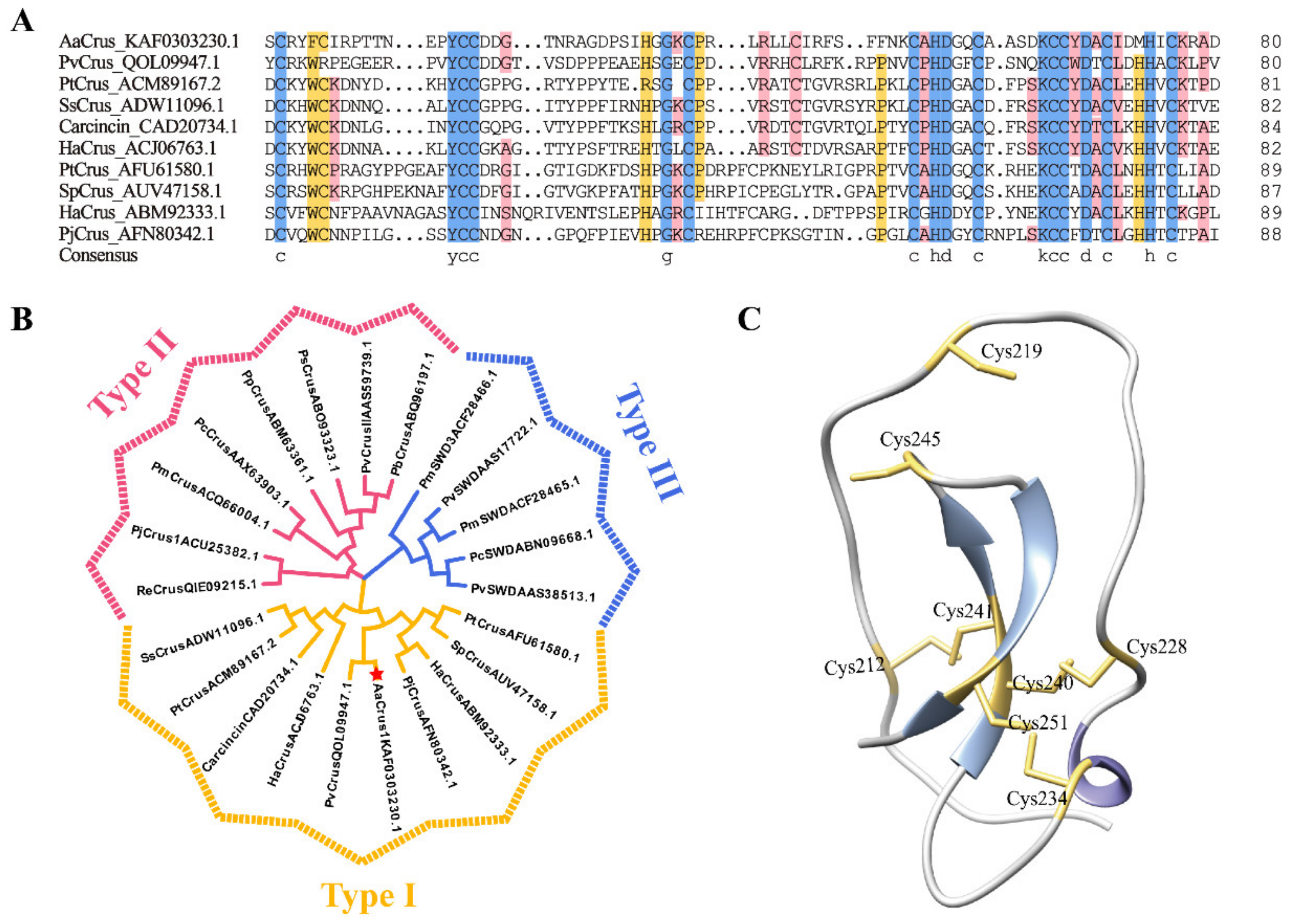
3.1. Sequence and Structure Characterization of AaCrus1
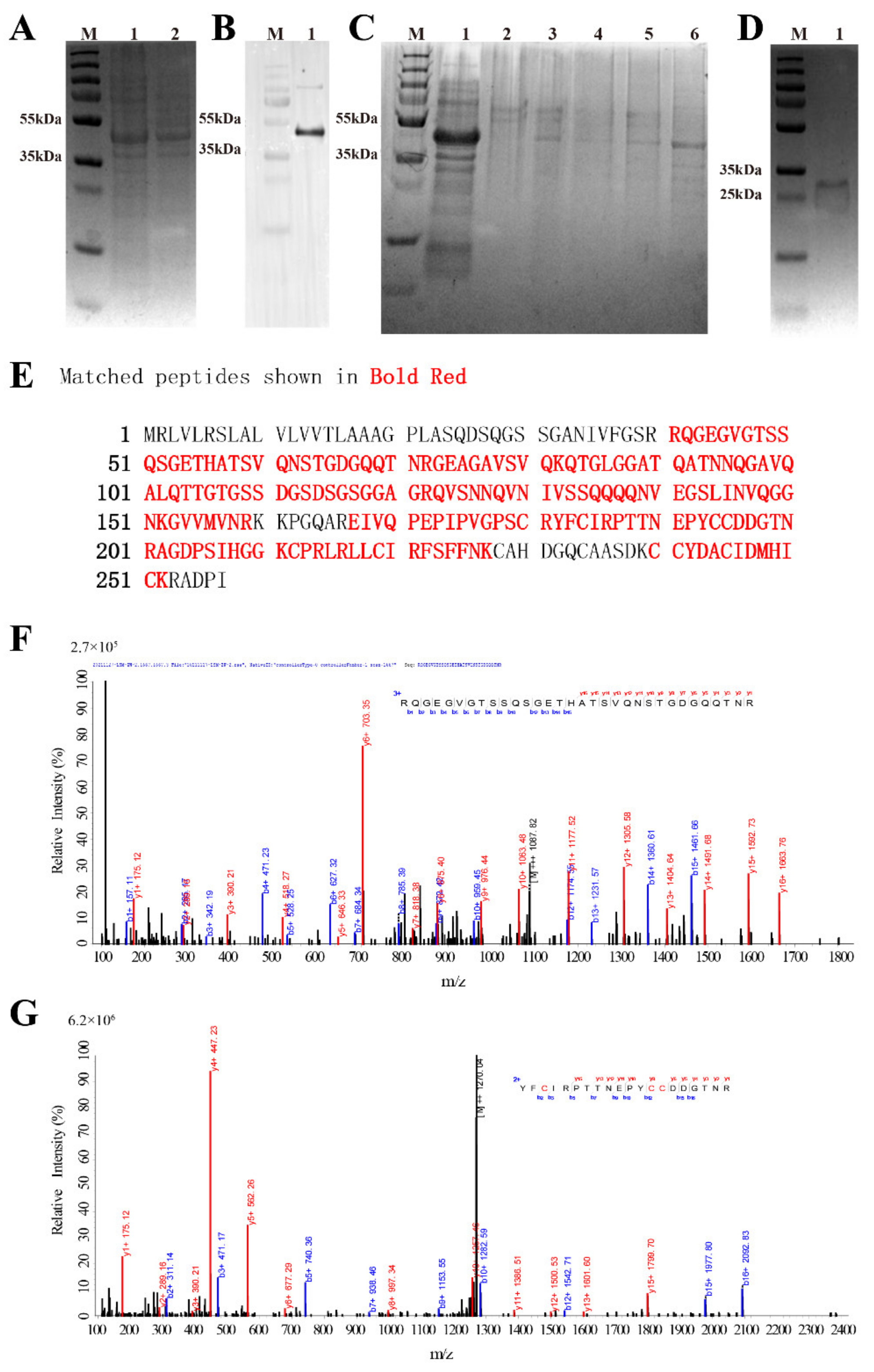
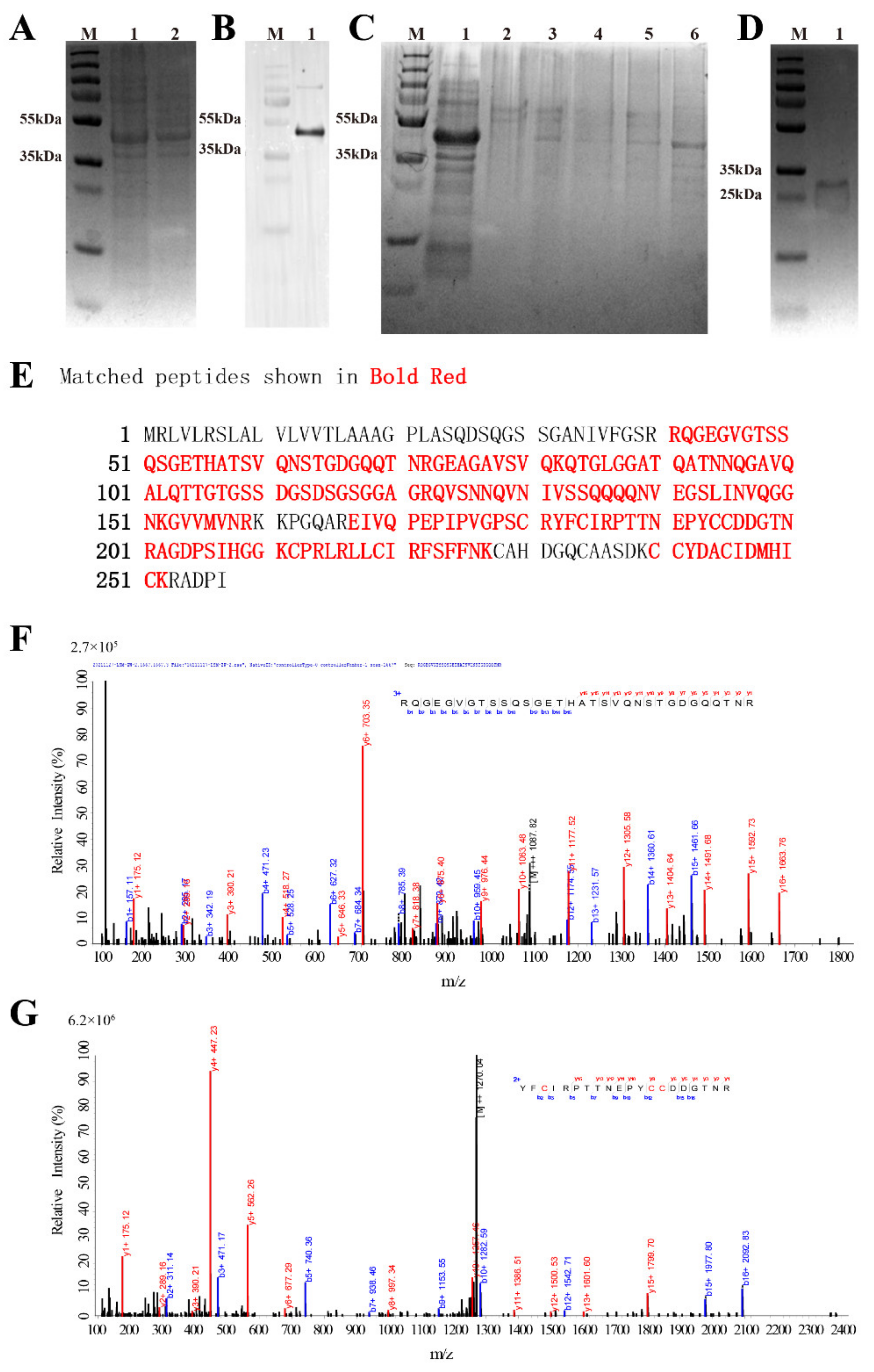
3.2. Recombinant Expression and Purification
3.3. LC-MS Identification Results
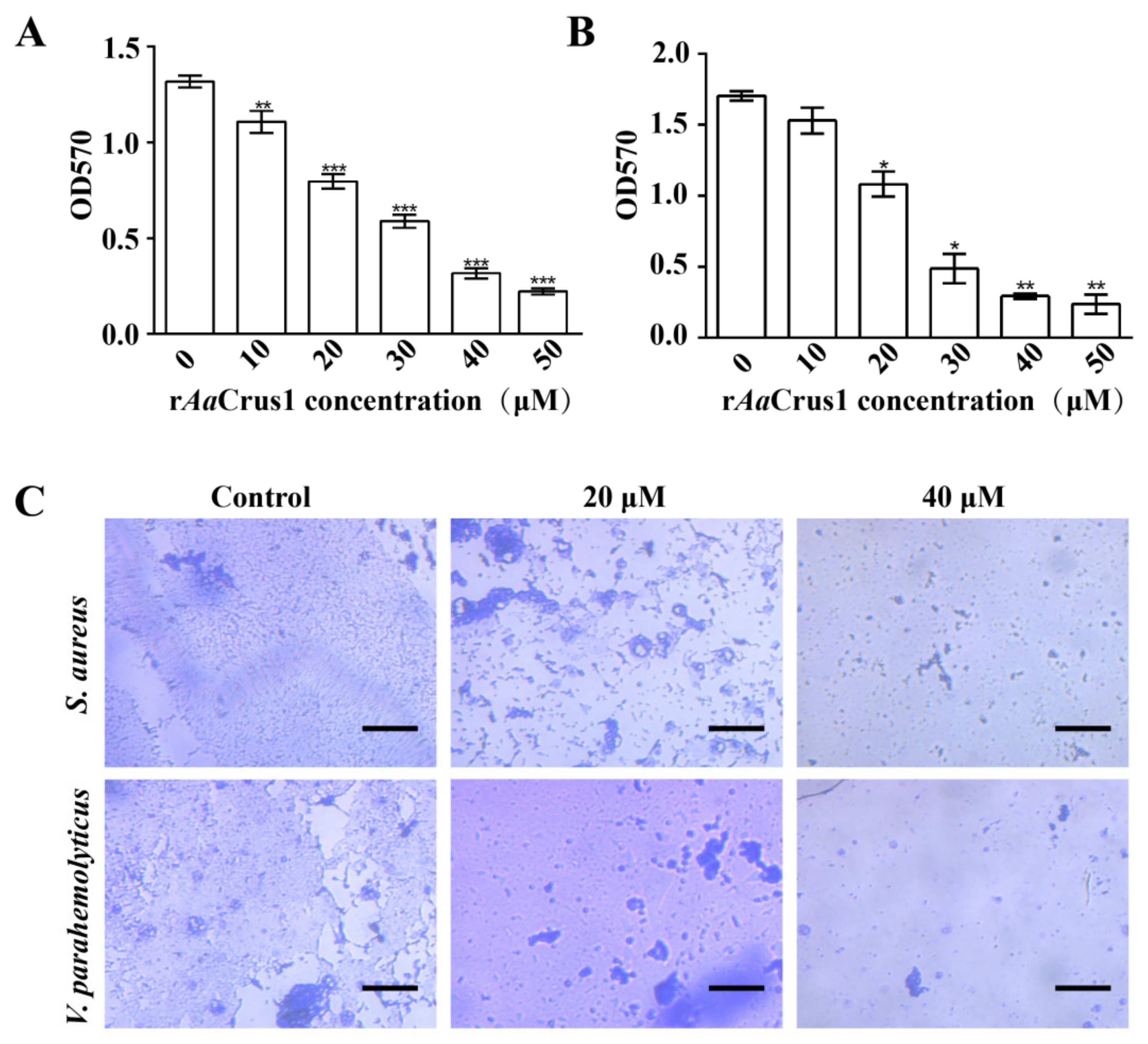
3.4. Antimicrobial Activity of rAaCrus1
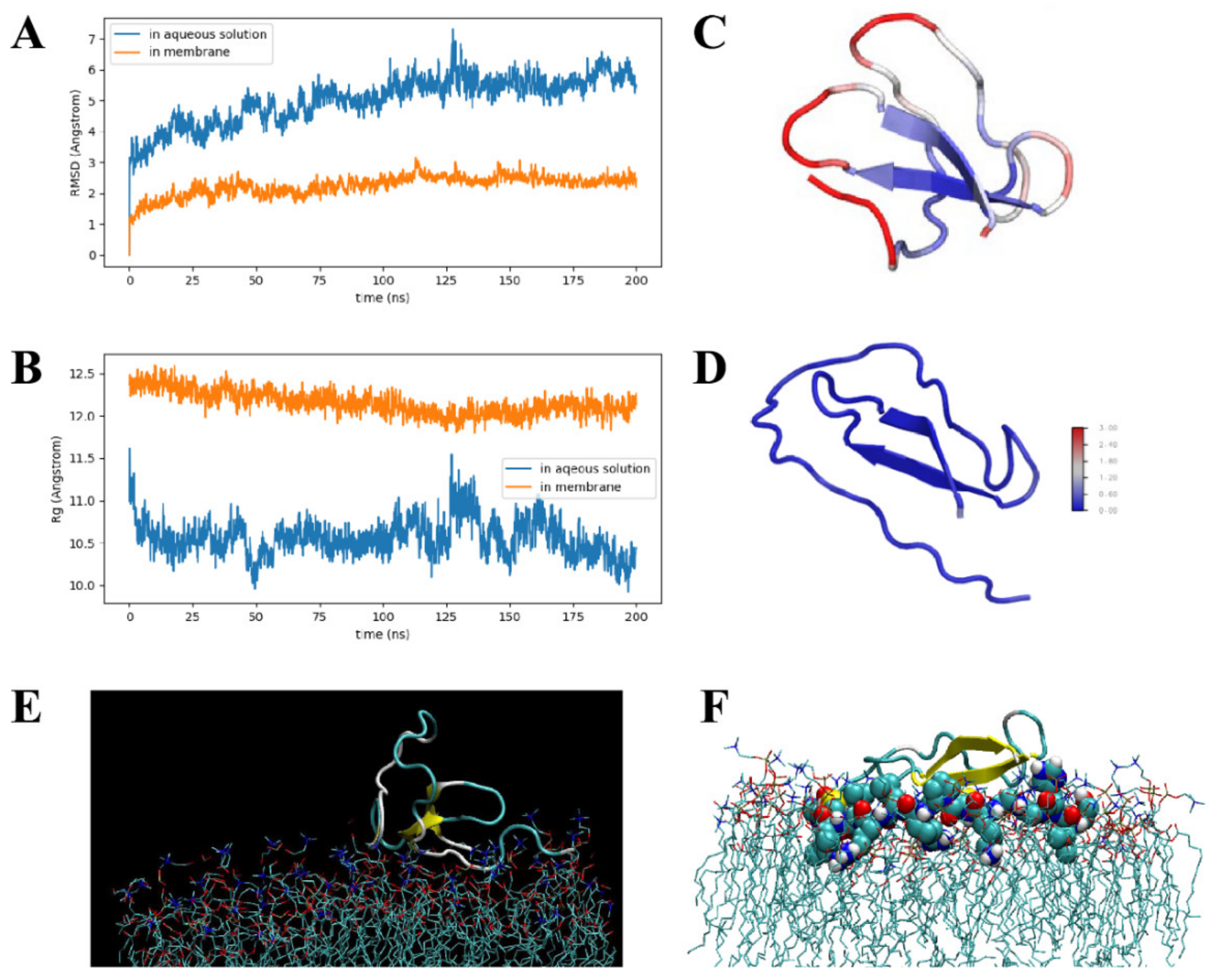
3.5. MD Simulations
3.6. Microorganism-Binding Activity of rAaCrus1
3.7. Agglutination Activity
3.8. Effects of rAaCrus1 on the Bacterial Morphology
3.9. Cell Membrane Permeability
3.10. Biofilm Inhibition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Loo, K.-Y.; Letchumanan, V.; Law, J.W.-F.; Pusparajah, P.; Goh, B.-H.; Ab Mutalib, N.-S.; He, Y.-W.; Lee, L.-H. Incidence of Antibiotic Resistance in Vibrio spp. Rev. Aquac. 2020, 12, 2590–2608. [Google Scholar] [CrossRef]
- Raheem, N.; Straus, S.K. Mechanisms of Action for Antimicrobial Peptides with Antibacterial and Antibiofilm Functions. Front. Microbiol. 2019, 10, 2866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Separovic, F.; O’Brien-Simpson, N.M.; Wade, J.D. Chemically Modified and Conjugated Antimicrobial Peptides against Superbugs. Chem. Soc. Rev. 2021, 50, 4932–4973. [Google Scholar] [CrossRef] [PubMed]
- Aisenbrey, C.; Marquette, A.; Bechinger, B. The mechanisms of action of cationic antimicrobial peptides refined by novel concepts from biophysical investigations. In Antimicrobial Peptides; Matsuzaki, K., Ed.; Springer Nature: Singapore, 2019; pp. 33–64. [Google Scholar] [CrossRef] [Green Version]
- Chung, C.-R.; Jhong, J.-H.; Wang, Z.; Chen, S.; Wan, Y.; Horng, J.-T.; Lee, T.-Y. Characterization and Identification of Natural Antimicrobial Peptides on Different Organisms. Int. J. Mol. Sci. 2020, 21, 986. [Google Scholar] [CrossRef] [Green Version]
- Leite, M.L.; da Cunha, N.B.; Costa, F.F. Antimicrobial Peptides, Nanotechnology, and Natural Metabolites as Novel Approaches for Cancer Treatment. Pharmacol. Ther. 2018, 183, 160–176. [Google Scholar] [CrossRef]
- Smith, V.J.; Chisholm, J.R.S. Antimicrobial proteins in crustaceans. In Phylogenetic Perspectives on the Vertebrate Immune System; Beck, G., Sugumaran, M., Cooper, E.L., Eds.; Springer: Boston, MA, USA, 2001; pp. 95–112. [Google Scholar] [CrossRef]
- Zanjani, T.N.; Miranda-Saksena, M.; Cunningham, L.A.; Dehghani, F. Antimicrobial Peptides of Marine Crustaceans: The Potential and Challenges of Developing Therapeutic Agents. Curr. Med. Chem. 2018, 25, 2245–2259. [Google Scholar] [CrossRef]
- Matos, G.M.; Rosa, R.D. On the Silver Jubilee of Crustacean Antimicrobial Peptides. Rev. Aquac. 2021, 1–19. [Google Scholar] [CrossRef]
- Tincu, J.A.; Taylor, S.W. Antimicrobial Peptides from Marine Invertebrates. Antimicrob. Agents Chemother. 2004, 48, 3645–3654. [Google Scholar] [CrossRef] [Green Version]
- Destoumieux-Garzón, D.; Rosa, R.D.; Schmitt, P.; Barreto, C.; Vidal-Dupiol, J.; Mitta, G.; Gueguen, Y.; Bachère, E. Antimicrobial Peptides in Marine Invertebrate Health and Disease. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150300. [Google Scholar] [CrossRef] [Green Version]
- Smith, V.J.; Fernandes, J.M.O.; Kemp, G.D.; Hauton, C. Crustins: Enigmatic WAP Domain-Containing Antibacterial Proteins from Crustaceans. Dev. Comp. Immunol. 2008, 32, 758–772. [Google Scholar] [CrossRef] [Green Version]
- Vargas-Albores, F.; Martínez-Porchas, M. Crustins Are Distinctive Members of the WAP-Containing Protein Superfamily: An Improved Classification Approach. Dev. Comp. Immunol. 2017, 76, 9–17. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, J.; Sun, Y.; Sun, L. A Crustin from Hydrothermal Vent Shrimp: Antimicrobial Activity and Mechanism. Marine Drugs 2021, 19, 176. [Google Scholar] [CrossRef]
- Li, M.; Ma, C.; Zhu, P.; Yang, Y.; Lei, A.; Chen, X.; Liang, W.; Chen, M.; Xiong, J.; Li, C. A New Crustin Is Involved in the Innate Immune Response of Shrimp Litopenaeus vannamei. Fish Shellfish Immunol. 2019, 94, 398–406. [Google Scholar] [CrossRef]
- Yang, L.; Luo, M.; Guo, Z.; Zuo, H.; Weng, S.; He, J.; Xu, X. A Shrimp Gene Encoding a Single WAP Domain (SWD)-Containing Protein Regulated by JAK-STAT and NF-KB Pathways. Dev. Comp. Immunol. 2020, 104, 103537. [Google Scholar] [CrossRef]
- Tassanakajon, A.; Somboonwiwat, K.; Amparyup, P. Sequence Diversity and Evolution of Antimicrobial Peptides in Invertebrates. Dev. Comp. Immunol. 2015, 48, 324–341. [Google Scholar] [CrossRef]
- Ilić, N.; Novković, M.; Guida, F.; Xhindoli, D.; Benincasa, M.; Tossi, A.; Juretić, D. Selective Antimicrobial Activity and Mode of Action of Adepantins, Glycine-Rich Peptide Antibiotics Based on Anuran Antimicrobial Peptide Sequences. Biochim. Biophys. Acta Biomembr. 2013, 1828, 1004–1012. [Google Scholar] [CrossRef] [Green Version]
- Sruthy, K.S.; Nair, A.; Puthumana, J.; Antony, S.P.; Singh, I.S.B.; Philip, R. Molecular Cloning, Recombinant Expression and Functional Characterization of an Antimicrobial Peptide, Crustin from the Indian White Shrimp, Fenneropenaeus indicus. Fish Shellfish Immunol. 2017, 71, 83–94. [Google Scholar]
- Khandeparker, L.; Anil, A.C.; Desai, D.V. Immuno-Modulation of Settlement Cues in the Barnacle, Amphibalanus amphitrite: Significance of Circulating Haemocytes. Hydrobiologia 2019, 830, 229–241. [Google Scholar] [CrossRef]
- Rajitha, K.; Nancharaiah, Y.V.; Venugopalan, V.P. Role of Bacterial Biofilms and Their EPS on Settlement of Barnacle (Amphibalanus Reticulatus) Larvae. Int. Biodeterior. Biodegrad. 2020, 150, 104958. [Google Scholar] [CrossRef]
- Zhou, L.; Li, G.; Jiao, Y.; Huang, D.; Li, A.; Chen, H.; Liu, Y.; Li, S.; Li, H.; Wang, C. Molecular and Antimicrobial Characterization of a Group G Anti-Lipopolysaccharide Factor (ALF) from Penaeus monodon. Fish Shellfish Immunol. 2019, 94, 149–156. [Google Scholar] [CrossRef]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 Improves Signal Peptide Predictions Using Deep Neural Networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree of Life (ITOL) v5: An Online Tool for Phylogenetic Tree Display and Annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Roy, A.; Kucukural, A.; Zhang, Y. I-TASSER: A Unified Platform for Automated Protein Structure and Function Prediction. Nat. Protoc. 2010, 5, 725–738. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Chen, J.; Li, J.; Shang, C.; Wang, C. Structural Characteristics, Prokaryotic Expression and Activity Analysis of Antimicrobial Peptide ALFPm10 from Penaeus monodon. Int. J. Peptide Res. Ther. 2022, 28, 1–10. [Google Scholar] [CrossRef]
- Li, W.; Lin, F.; Hung, A.; Barlow, A.; Sani, M.-A.; Paolini, R.; Singleton, W.; Holden, J.; Hossain, M.A.; Separovic, F.; et al. Enhancing Proline-Rich Antimicrobial Peptide Action by Homodimerization: Influence of Bifunctional Linker. Chem. Sci. 2022. [Google Scholar] [CrossRef]
- Lin, B.; Hung, A.; Li, R.; Barlow, A.; Singleton, W.; Matthyssen, T.; Sani, M.-A.; Hossain, M.A.; Wade, J.D.; O’Brien-Simpson, N.M.; et al. Systematic Comparison of Activity and Mechanism of Antimicrobial Peptides against Nosocomial Pathogens. Eur. J. Med. Chem. 2022, 231, 114135. [Google Scholar] [CrossRef]
- Schmid, N.; Eichenberger, A.P.; Choutko, A.; Riniker, S.; Winger, M.; Mark, A.E.; van Gunsteren, W.F. Definition and Testing of the GROMOS Force-Field Versions 54A7 and 54B7. Eur. Biophys. J. 2011, 40, 843–856. [Google Scholar] [CrossRef]
- Berger, O.; Edholm, O.; Jähnig, F. Molecular Dynamics Simulations of a Fluid Bilayer of Dipalmitoylphosphatidylcholine at Full Hydration, Constant Pressure, and Constant Temperature. Biophys. J. 1997, 72, 2002–2013. [Google Scholar] [CrossRef] [Green Version]
- Berendsen, H.J.C.; Postma, J.P.M.; van Gunsteren, W.F.; Hermans, J. Interaction models for water in relation to protein hydration. In Intermolecular Forces, Proceedings of the Fourteenth Jerusalem Symposium on Quantum Chemistry and Biochemistry, Jerusalem, Israel, 13–16 April 1981; Pullman, B., Ed.; Springer: Dordrecht, The Netherlands, 1981; pp. 331–342. [Google Scholar]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High Performance Molecular Simulations through Multi-Level Parallelism from Laptops to Supercomputers. SoftwareX 2015, 1, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Essmann, U.; Perera, L.; Berkowitz, M.L.; Darden, T.; Lee, H.; Pedersen, L.G. A Smooth Particle Mesh Ewald Method. J. Chem. Phys. 1995, 103, 8577–8593. [Google Scholar] [CrossRef] [Green Version]
- Darden, T.; York, D.; Pedersen, L. Particle Mesh Ewald: An N Log (N) Method for Ewald Sums in Large Systems. J. Chem. Phys. 1993, 98, 10089–10092. [Google Scholar] [CrossRef] [Green Version]
- Nosé, S. A Molecular Dynamics Method for Simulations in the Canonical Ensemble. Mol. Phys. 1984, 52, 255–268. [Google Scholar] [CrossRef]
- Hoover, W.G. Canonical Dynamics: Equilibrium Phase-Space Distributions. Phys. Rev. A 1985, 31, 1695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parrinello, M.; Rahman, A. Polymorphic Transitions in Single Crystals: A New Molecular Dynamics Method. J. Appl. Phys. 1981, 52, 7182–7190. [Google Scholar] [CrossRef]
- Hess, B.; Bekker, H.; Berendsen, H.J.C.; Fraaije, J.G.E.M. LINCS: A Linear Constraint Solver for Molecular Simulations. J. Comput. Chem. 1997, 18, 1463–1472. [Google Scholar] [CrossRef]
- Li, W.; O’Brien-Simpson, N.M.; Yao, S.; Tailhades, J.; Reynolds, E.C.; Dawson, R.M.; Otvos, L., Jr.; Hossain, M.A.; Separovic, F.; Wade, J.D. C-Terminal Modification and Multimerization Increase the Efficacy of a Proline-Rich Antimicrobial Peptide. Chem. Eur. J. 2017, 23, 390–396. [Google Scholar] [CrossRef]
- Li, W.; O’Brien-Simpson, N.M.; Tailhades, J.; Pantarat, N.; Dawson, R.M.; Otvos, L.; Reynolds, E.C.; Separovic, F.; Hossain, M.A.; Wade, J.D. Multimerization of a Proline-Rich Antimicrobial Peptide, Chex-Arg20, Alters Its Mechanism of Interaction with the Escherichia coli Membrane. Chem. Biol. 2015, 22, 1250–1258. [Google Scholar] [CrossRef]
- Sperstad, S.V.; Haug, T.; Paulsen, V.; Rode, T.M.; Strandskog, G.; Solem, S.T.; Styrvold, O.B.; Stensvåg, K. Characterization of Crustins from the Hemocytes of the Spider Crab, Hyas araneus, and the Red King Crab, Paralithodes camtschaticus. Dev. Comp. Immunol. 2009, 33, 583–591. [Google Scholar] [CrossRef]
- Smith, V.J.; Dyrynda, E.A. Antimicrobial Proteins: From Old Proteins, New Tricks. Mol. Immunol. 2015, 68, 383–398. [Google Scholar] [CrossRef] [Green Version]
- Bandeira, P.T.; Vernal, J.; Matos, G.M.; Farias, N.D.; Terenzi, H.; Pinto, A.R.; Barracco, M.A.; Rosa, R.D. A Type IIa Crustin from the Pink Shrimp Farfantepenaeus paulensis (CrusFpau) Is Constitutively Synthesized and Stored by Specific Granule-Containing Hemocyte Subpopulations. Fish Shellfish Immunol. 2020, 97, 294–299. [Google Scholar] [CrossRef]
- Suthianthong, P.; Pulsook, N.; Supungul, P.; Tassanakajon, A.; Rimphanitchayakit, V. A Double WAP Domain-Containing Protein PmDWD from the Black Tiger Shrimp Penaeus monodon Is Involved in the Controlling of Proteinase Activities in Lymphoid Organ. Fish Shellfish Immunol. 2011, 30, 783–790. [Google Scholar] [CrossRef]
- Li, W.; O’Brien-Simpson, N.M.; Hossain, M.A.; Wade, J.D. The 9-Fluorenylmethoxycarbonyl (Fmoc) Group in Chemical Peptide Synthesis—Its Past, Present, and Future. Aust. J. Chem. 2020, 73, 271–276. [Google Scholar] [CrossRef] [Green Version]
- Panavas, T.; Sanders, C.; Butt, T.R. SUMO Fusion Technology for Enhanced Protein Production in Prokaryotic and Eukaryotic Expression Systems. In SUMO Protocols; Ulrich, H.D., Ed.; Humana Press: Totowa, NJ, USA, 2009; pp. 303–317. [Google Scholar]
- Shimokawa-Falcão, L.H.A.L.; Caporrino, M.C.; Barbaro, K.C.; Della-Casa, M.S.; Magalhães, G.S. Toxin Fused with SUMO Tag: A New Expression Vector Strategy to Obtain Recombinant Venom Toxins with Easy Tag Removal inside the Bacteria. Toxins 2017, 9, 82. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Li, S.; Li, F.; Xiang, J. Structure and Bioactivity of a Modified Peptide Derived from the LPS-Binding Domain of an Anti-Lipopolysaccharide Factor (ALF) of Shrimp. Marine Drugs 2016, 14, 96. [Google Scholar] [CrossRef] [Green Version]
- Malanovic, N.; Lohner, K. Antimicrobial Peptides Targeting Gram-Positive Bacteria. Pharmaceuticals 2016, 9, 59. [Google Scholar] [CrossRef] [Green Version]
- Koprivnjak, T.; Weidenmaier, C.; Peschel, A.; Weiss, J.P. Wall Teichoic Acid Deficiency in Staphylococcus aureus Confers Selective Resistance to Mammalian Group IIA Phospholipase A2 and Human β-Defensin 3. Infect. Immun. 2008, 76, 2169–2176. [Google Scholar] [CrossRef] [Green Version]
- Martin, N.I.; Breukink, E. The Expanding Role of Lipid II as a Target for Lantibiotics. Future Microbiol. 2007, 2, 513–525. [Google Scholar] [CrossRef]
- De Kruijff, B.; van Dam, V.; Breukink, E. Lipid II: A Central Component in Bacterial Cell Wall Synthesis and a Target for Antibiotics. Prostaglandins Leukot. Essent. Fat. Acids 2008, 79, 117–121. [Google Scholar] [CrossRef]
- Fernández, L.; Breidenstein, E.B.M.; Hancock, R.E.W. Creeping Baselines and Adaptive Resistance to Antibiotics. Drug Resist. Updates 2011, 14, 1–21. [Google Scholar] [CrossRef]
- Hancock, R.E.W.; Alford, M.A.; Haney, E.F. Antibiofilm Activity of Host Defence Peptides: Complexity Provides Opportunities. Nat. Rev. Microbiol. 2021, 19, 786–797. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganism | Minimal Inhibitory Concentrations |
|---|---|
| rAaCrus1 | |
| Gram+ | |
| S. aureus | 3.5 μM |
| Bacillus sp. T2 | 3.5 μM |
| Gram− | |
| V. alginolyticus | 7 μM |
| V. parahemolyticus | 28 μM |
| V. harveyi | 28 μM |
| V. anguillarum | 14 μM |
| Microorganisms | Minimal Agglutinating Concentrations (μM) | |
|---|---|---|
| rAaCrus1 + Ca2+ | rAaCrus1 | |
| Gram+ | ||
| S. aureus | 0.875 | 3.5 |
| Bacillus sp. T2 | 1.75 | 3.5 |
| Gram− | ||
| V. alginolyticus | 3.5 | 14 |
| V. parahemolyticus | 1.75 | 7 |
| V. harveyi | 3.5 | 7 |
| V. anguillarum | 7 | 14 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, W.; Xu, X.; Zhang, J.; Ye, T.; Zhou, Q.; Xu, Y.; Li, W.; Hu, Z.; Shang, C. Discovery and Characterization of a New Crustin Antimicrobial Peptide from Amphibalanus amphitrite. Pharmaceutics 2022, 14, 413. https://doi.org/10.3390/pharmaceutics14020413
Zhang W, Xu X, Zhang J, Ye T, Zhou Q, Xu Y, Li W, Hu Z, Shang C. Discovery and Characterization of a New Crustin Antimicrobial Peptide from Amphibalanus amphitrite. Pharmaceutics. 2022; 14(2):413. https://doi.org/10.3390/pharmaceutics14020413
Chicago/Turabian StyleZhang, Wei, Xiaohang Xu, Jun Zhang, Ting Ye, Qiao Zhou, Ying Xu, Wenyi Li, Zhangli Hu, and Chenjing Shang. 2022. "Discovery and Characterization of a New Crustin Antimicrobial Peptide from Amphibalanus amphitrite" Pharmaceutics 14, no. 2: 413. https://doi.org/10.3390/pharmaceutics14020413
APA StyleZhang, W., Xu, X., Zhang, J., Ye, T., Zhou, Q., Xu, Y., Li, W., Hu, Z., & Shang, C. (2022). Discovery and Characterization of a New Crustin Antimicrobial Peptide from Amphibalanus amphitrite. Pharmaceutics, 14(2), 413. https://doi.org/10.3390/pharmaceutics14020413