
In Vitro Investigations on Optimizing and Nebulization of IVT-mRNA Formulations for Potential Pulmonary-Based Alpha-1-Antitrypsin Deficiency Treatment †



Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture
2.3. Preparation of IVT-mRNA Complexes
2.4. Size Measurements
2.5. Transfection of Cultured Cells
2.6. Luciferase Assay
2.7. Fluorescence Microscopy
2.8. MTT-Based Cytotoxicity Assay
2.9. Nebulization
2.10. Detection of Alpha-1-Antitrypsin Using Enzyme-Linked Immunosorbent Assay (ELISA)
2.11. Alpha-1-Antitrypsin Functional Assay
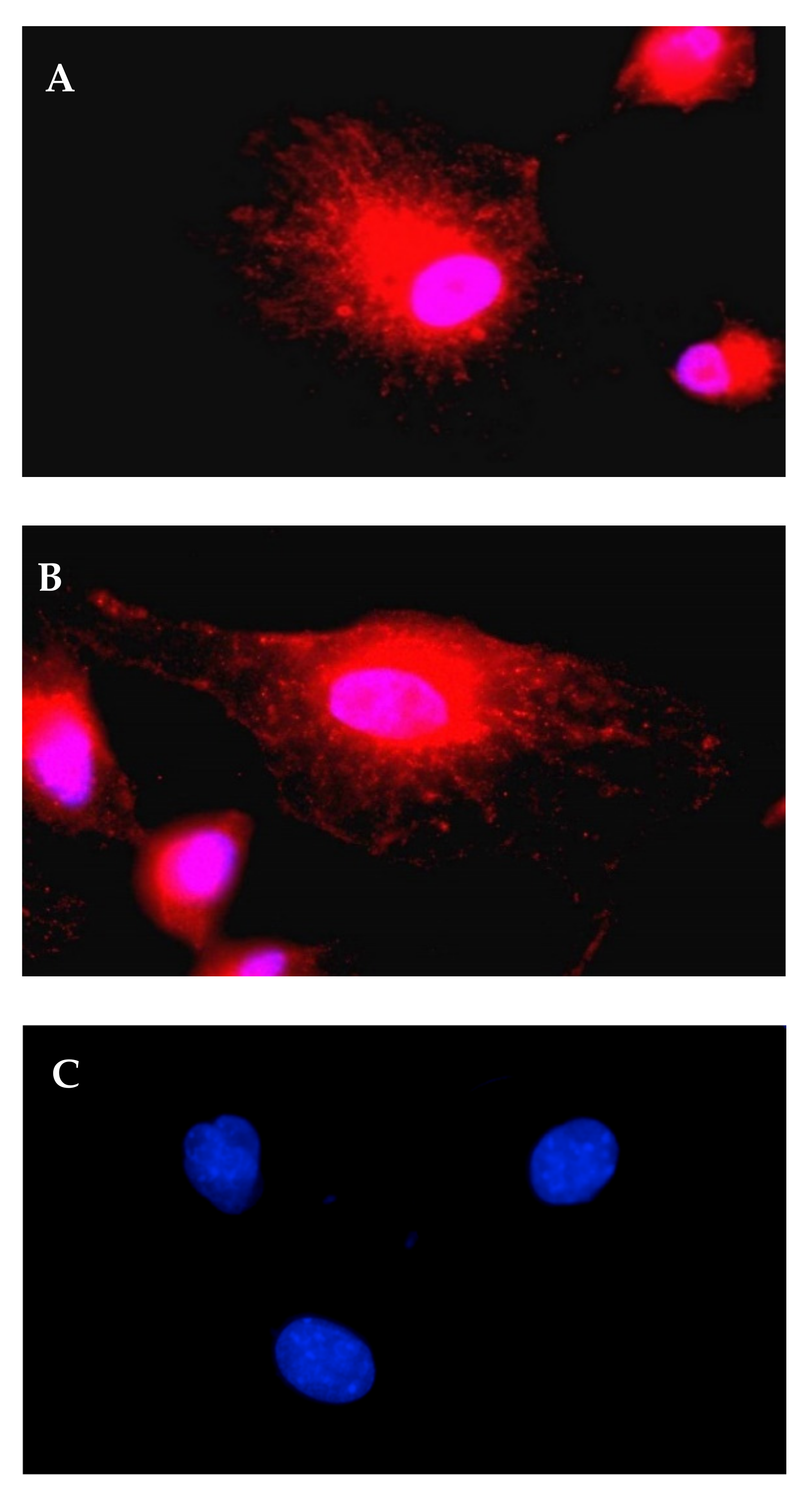
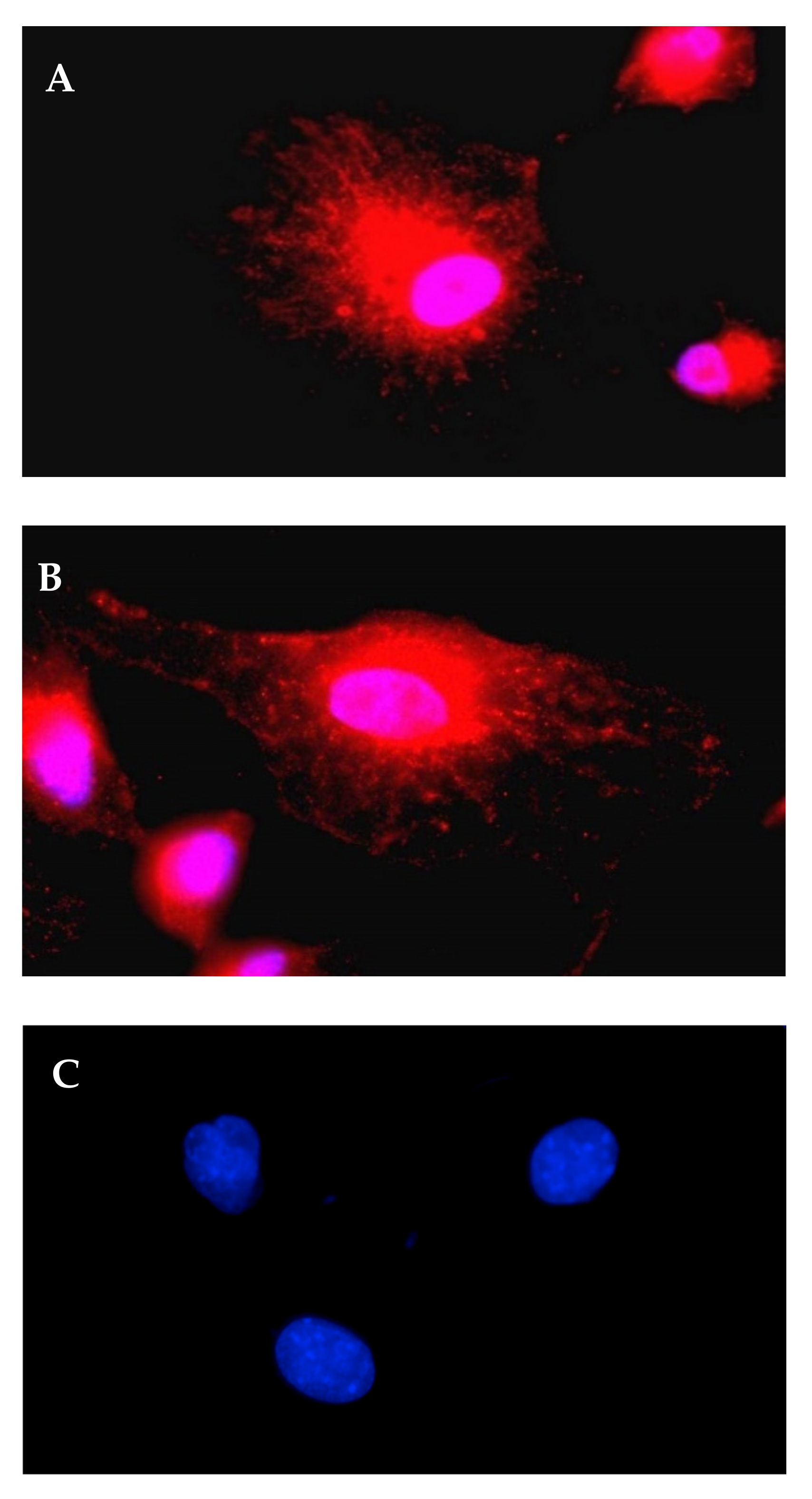
2.12. Immunofluorescence
2.13. Statistical Analysis
3. Results
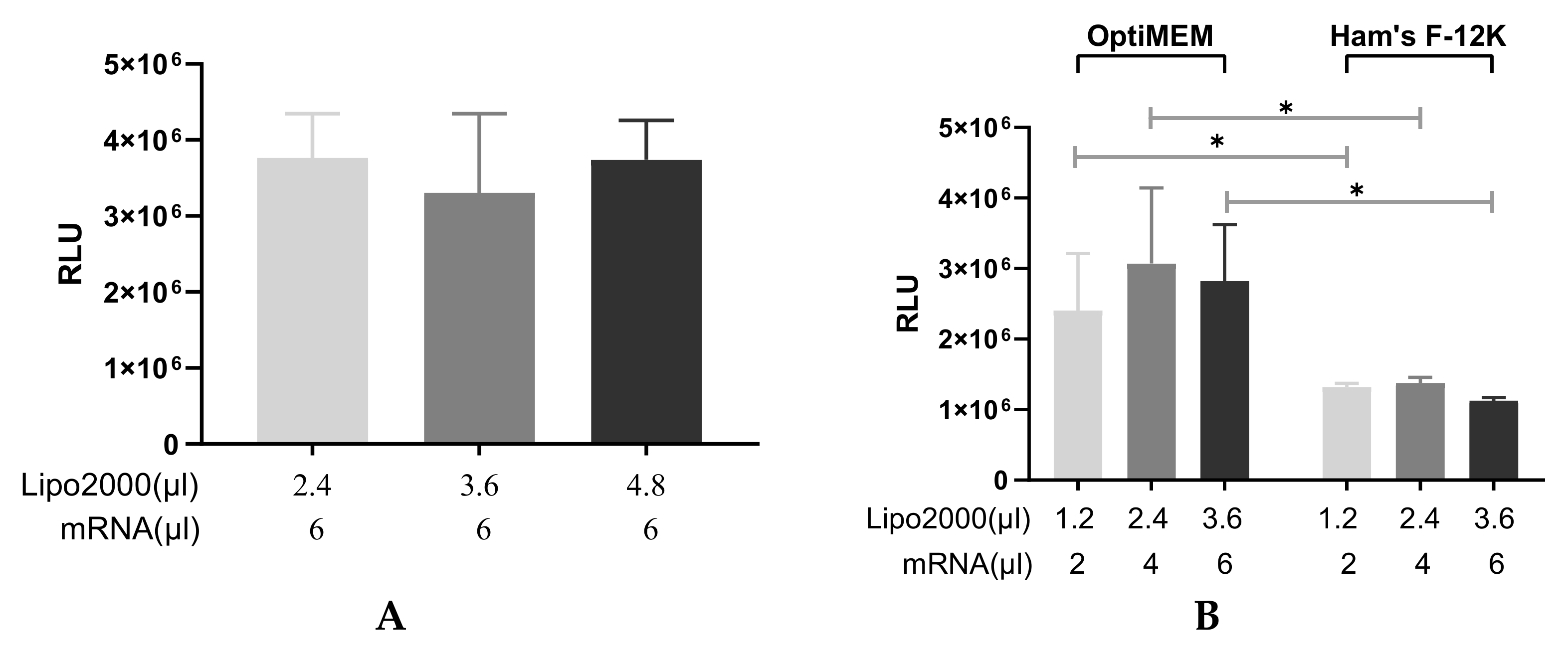
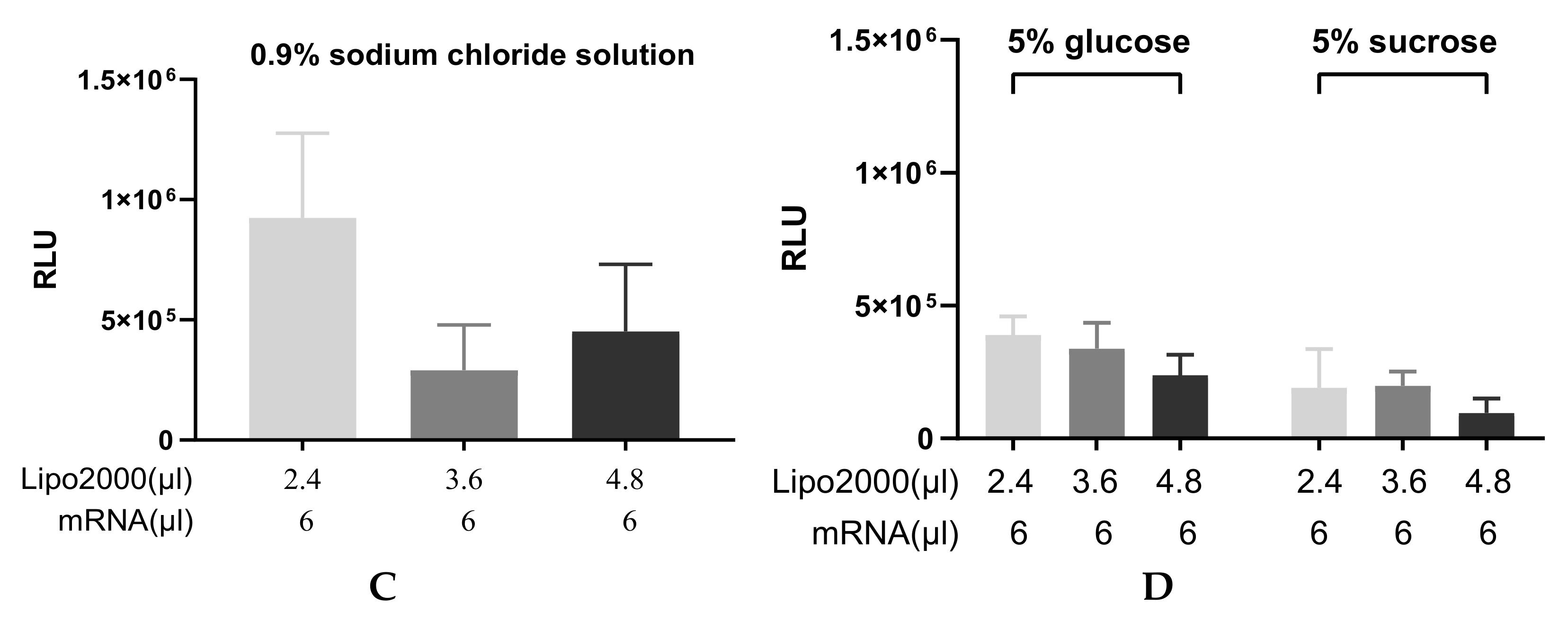
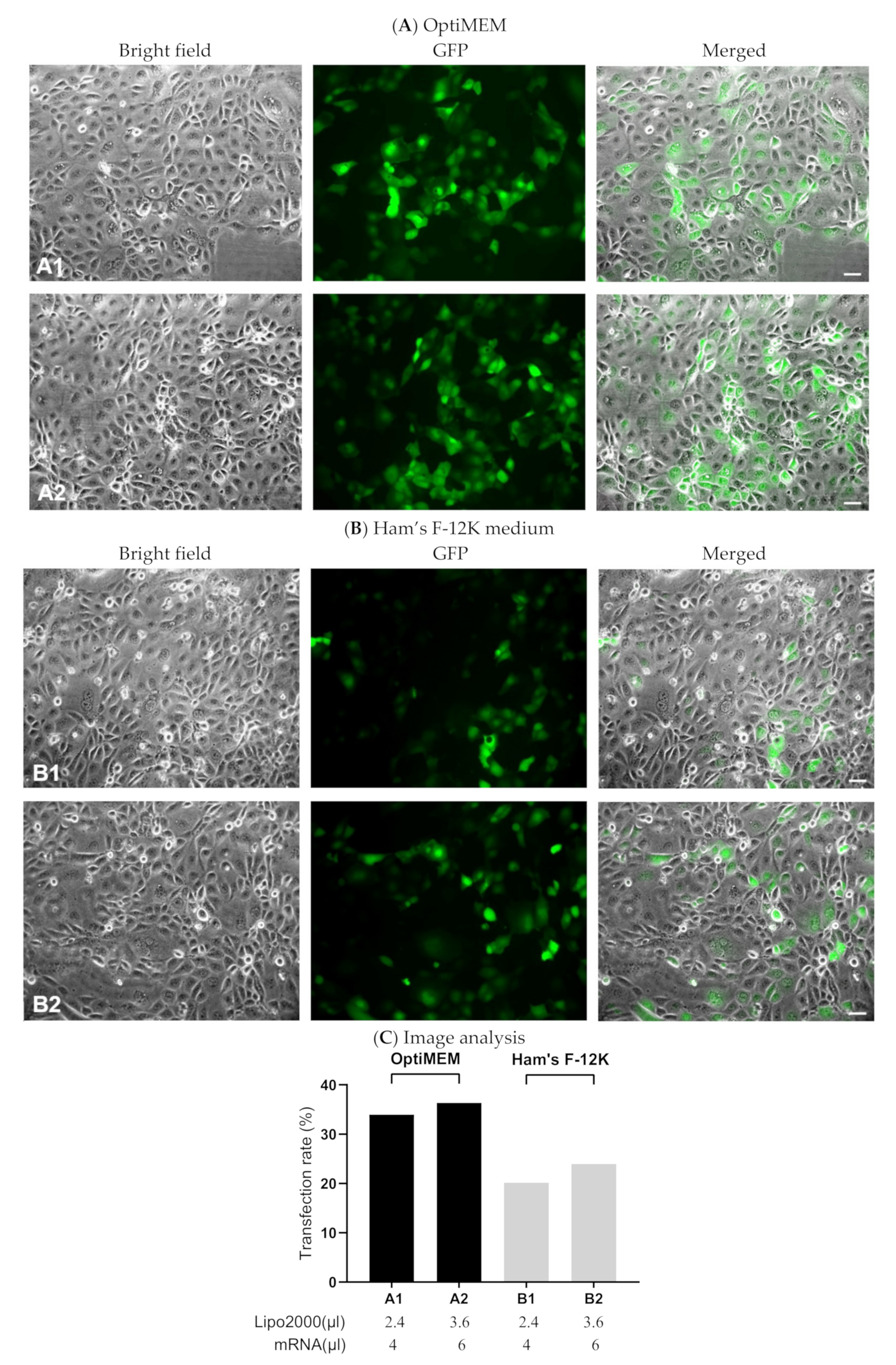
3.1. Optimization of the Transfection Process
3.1.1. Finding the Optimal Transfection Conditions
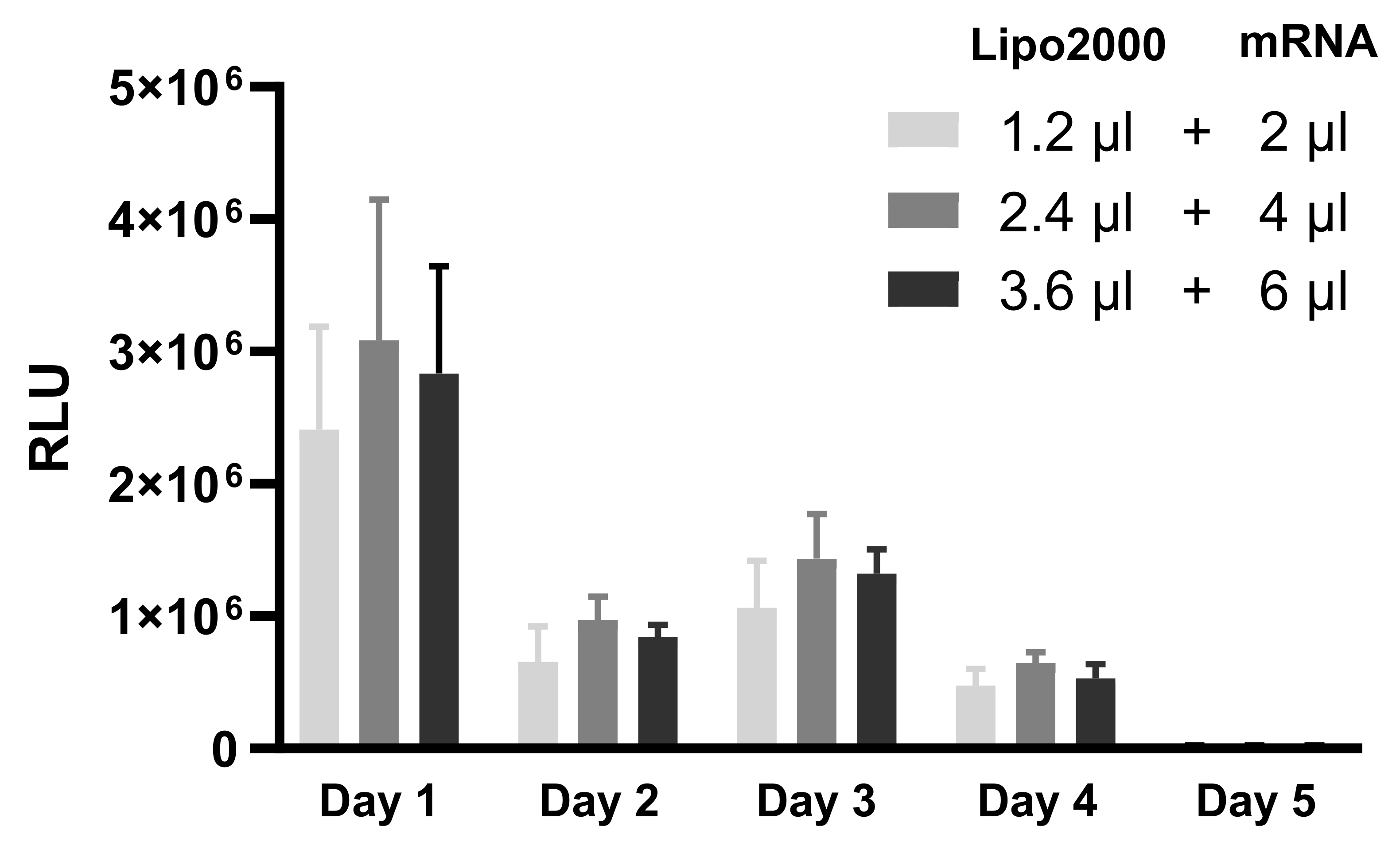
3.1.2. Duration of Protein Production
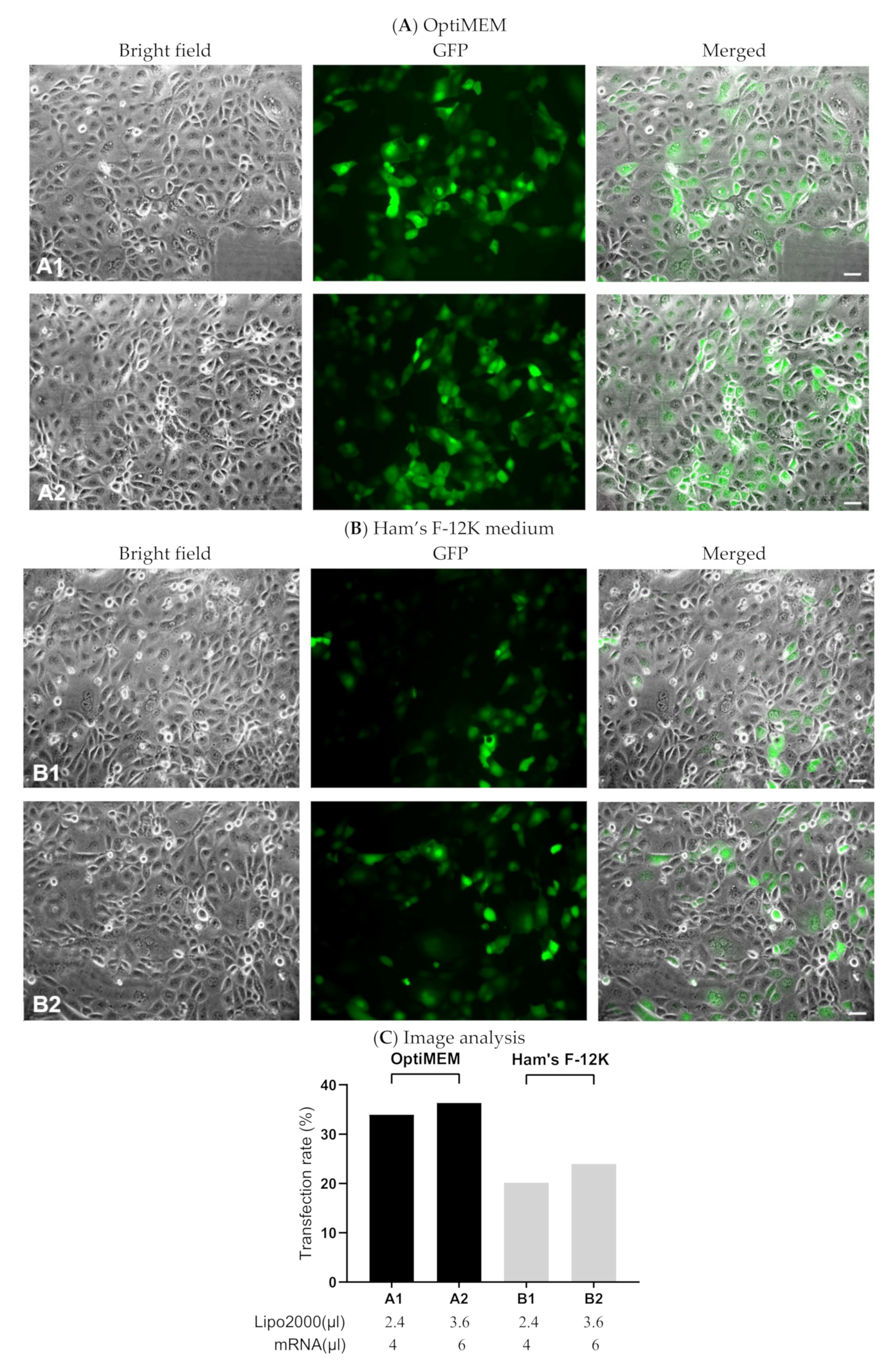
3.1.3. Evaluating Transfection Efficiency via GFP-mRNA
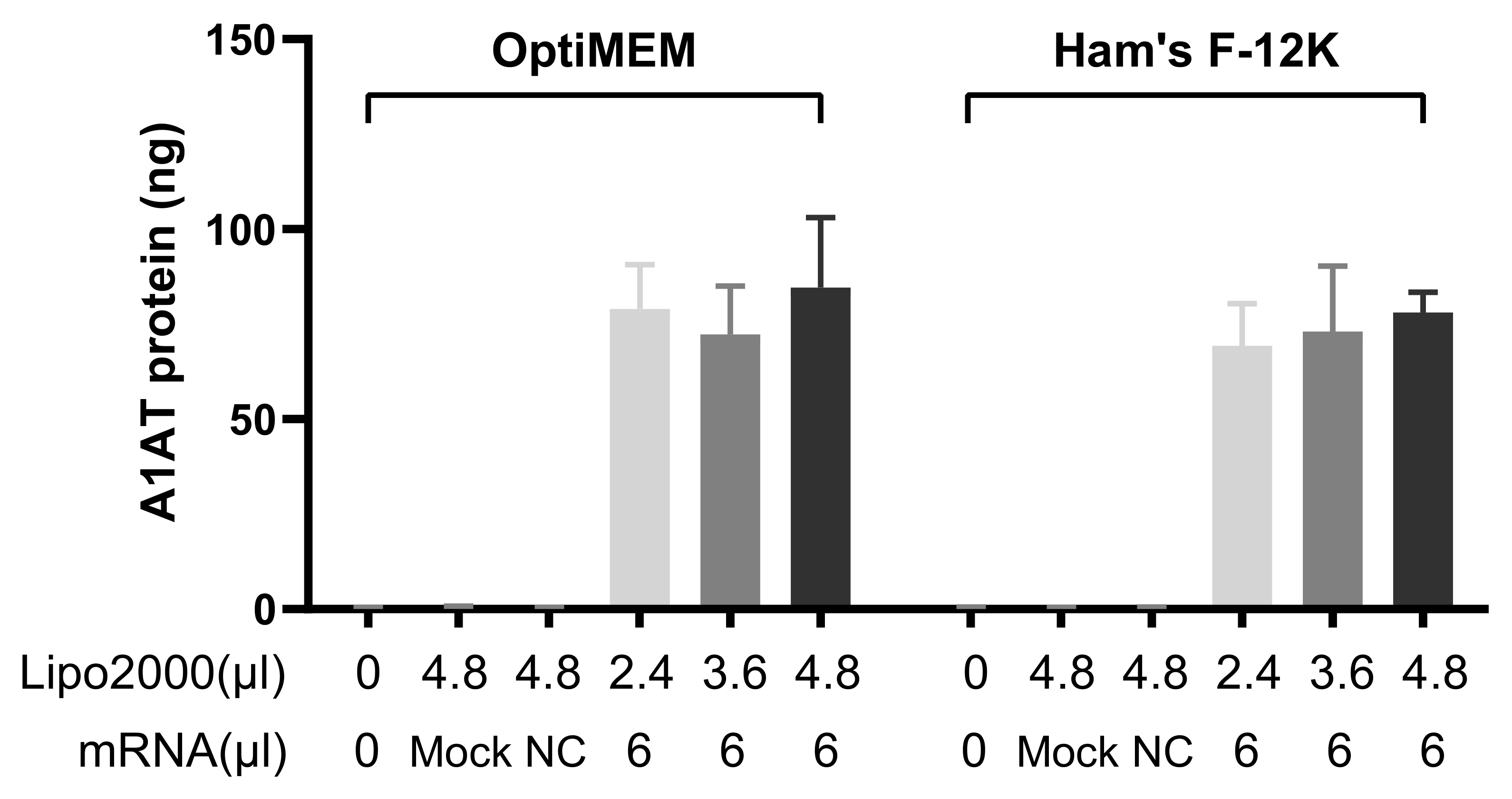
3.2. Transfection with IVT-mRNA-Encoding Alpha-1-Antitrypsin (A1AT)
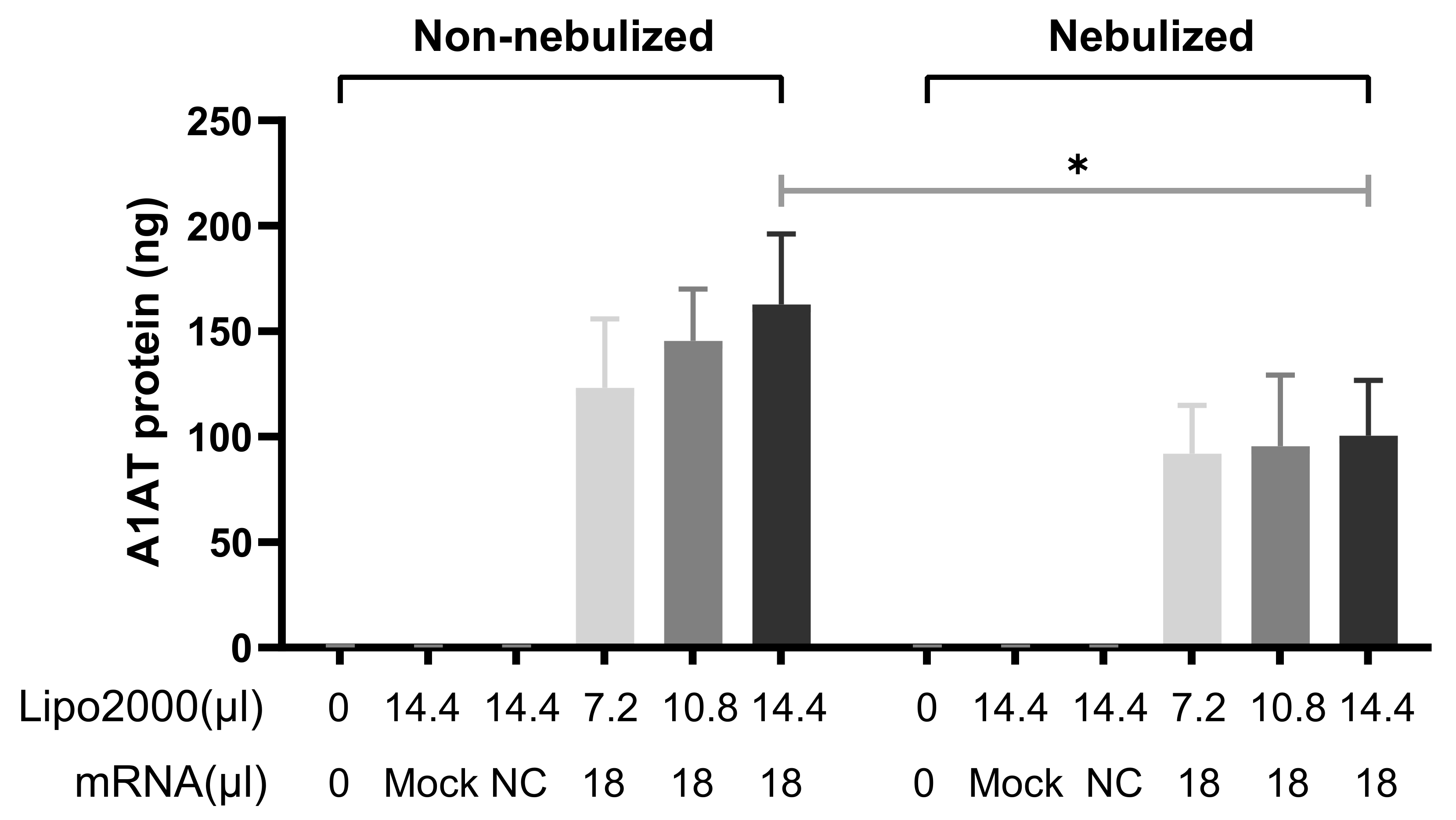
3.2.1. A1AT Expression Mediated by A1AT-mRNA Lipoplexes
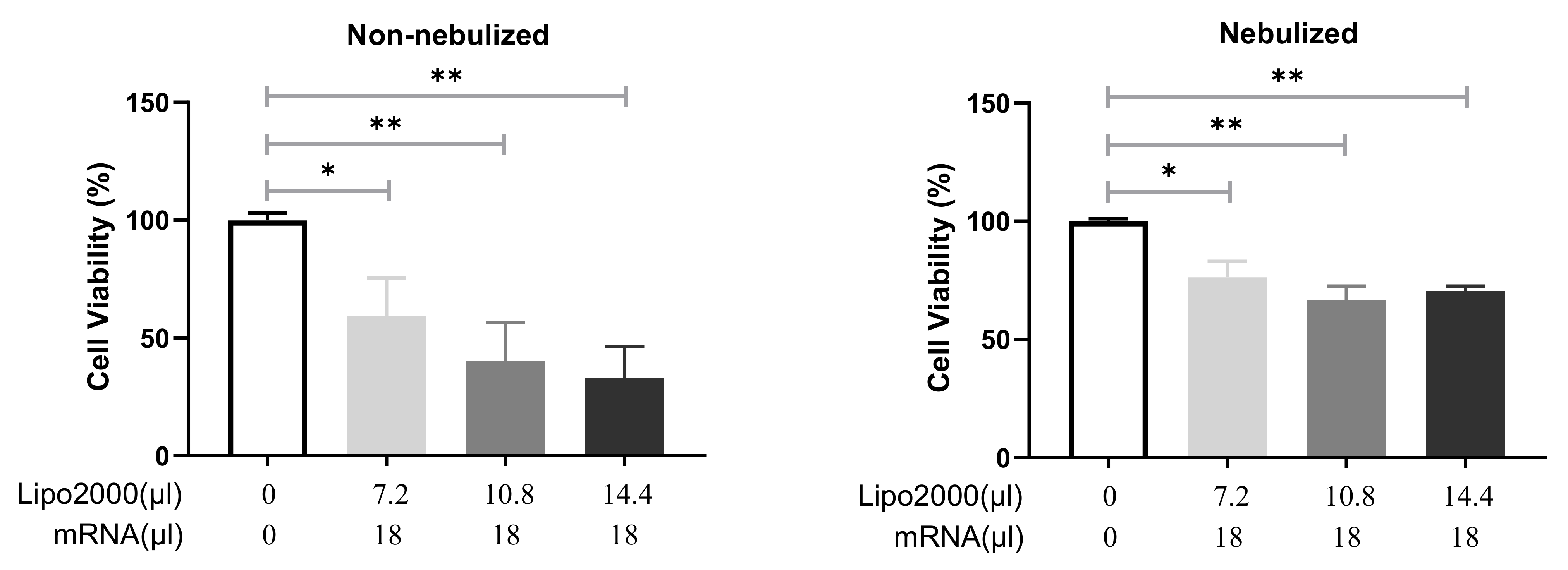
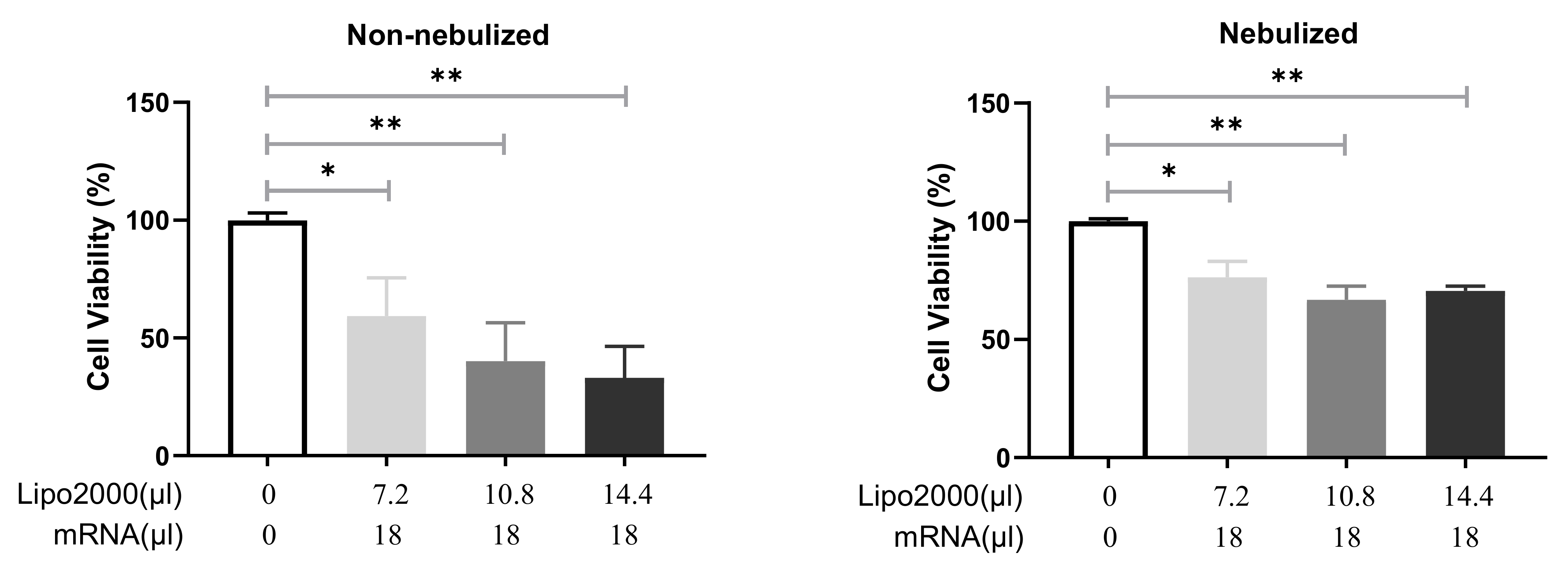
3.2.2. Cell Viability after Transfection with Different IVT-mRNA Lipoplexes
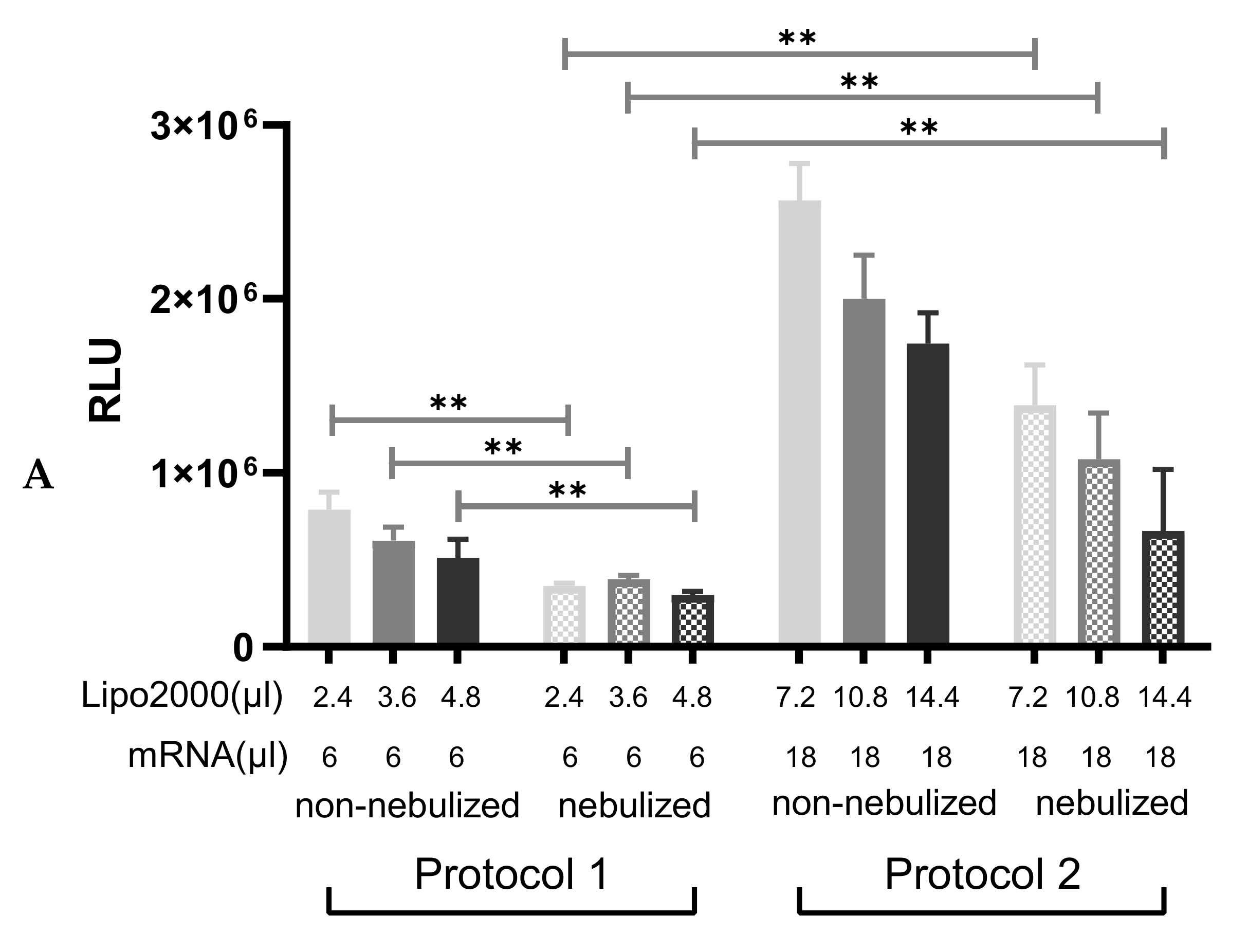
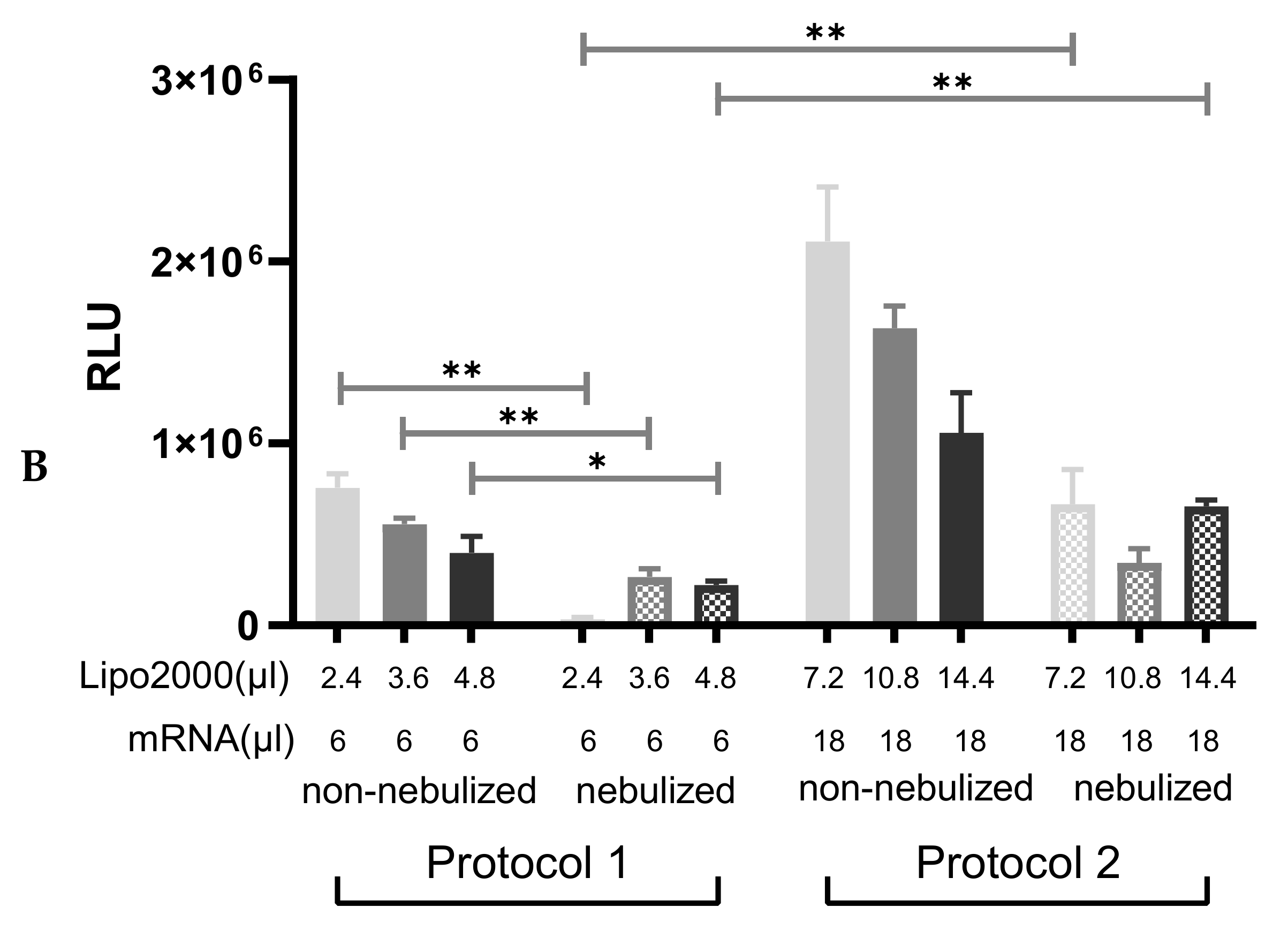
3.3. Nebulization of IVT-mRNA Complexes
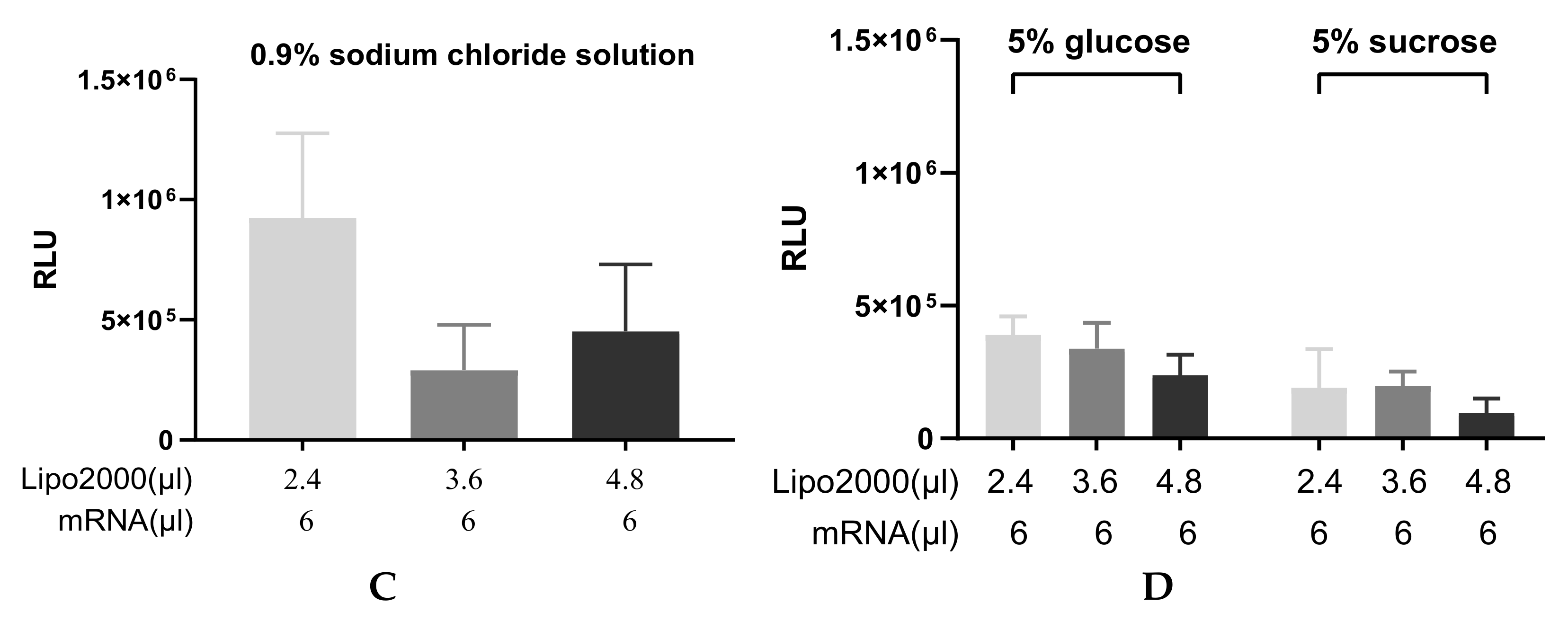
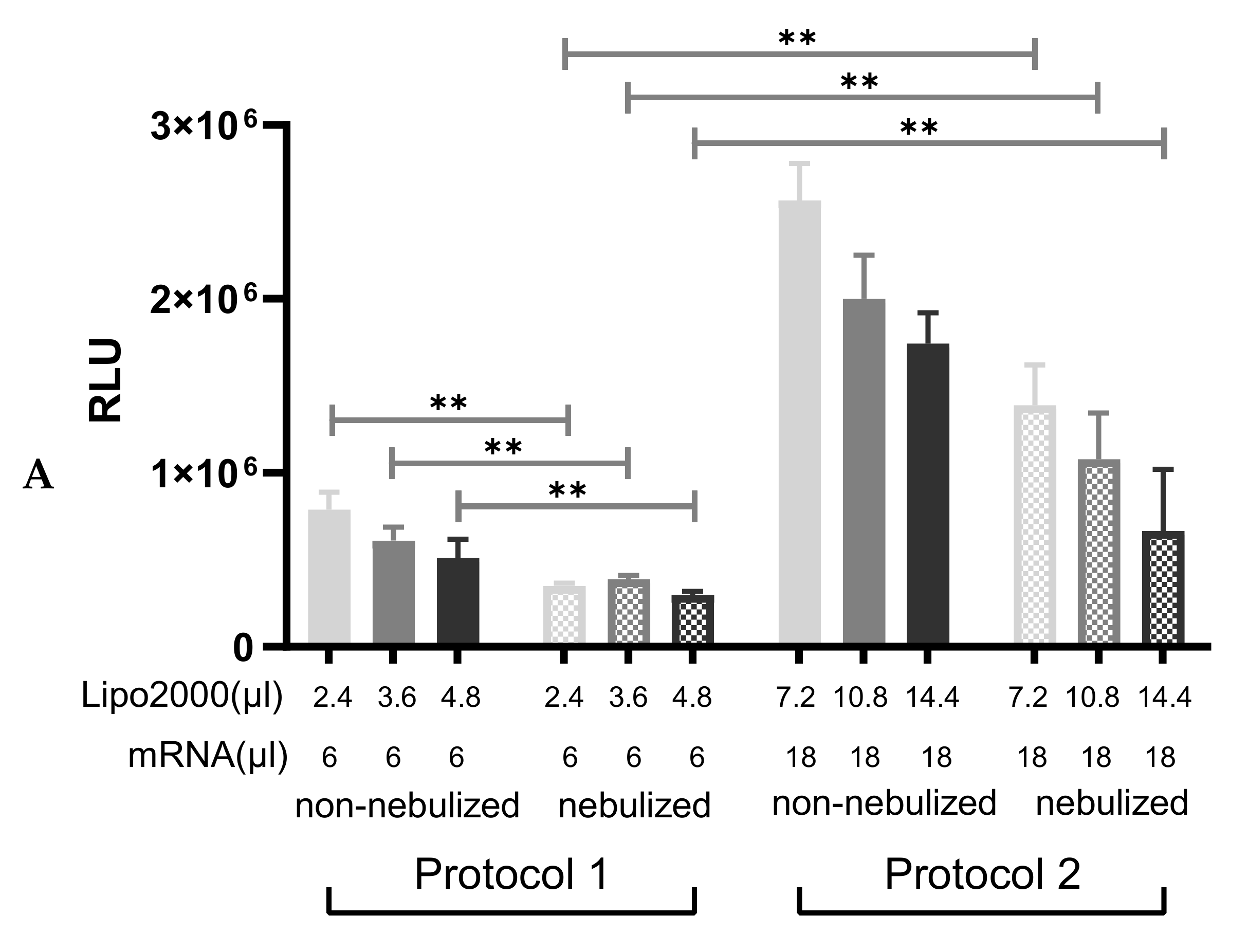
3.3.1. Particle Size and Transfection Efficiency of Lipoplexes after the Nebulization
3.3.2. Cell Viability following Transfection with Protocol 2
3.3.3. Nebulization of A1AT-mRNA/Lipofectamine2000 Complexes
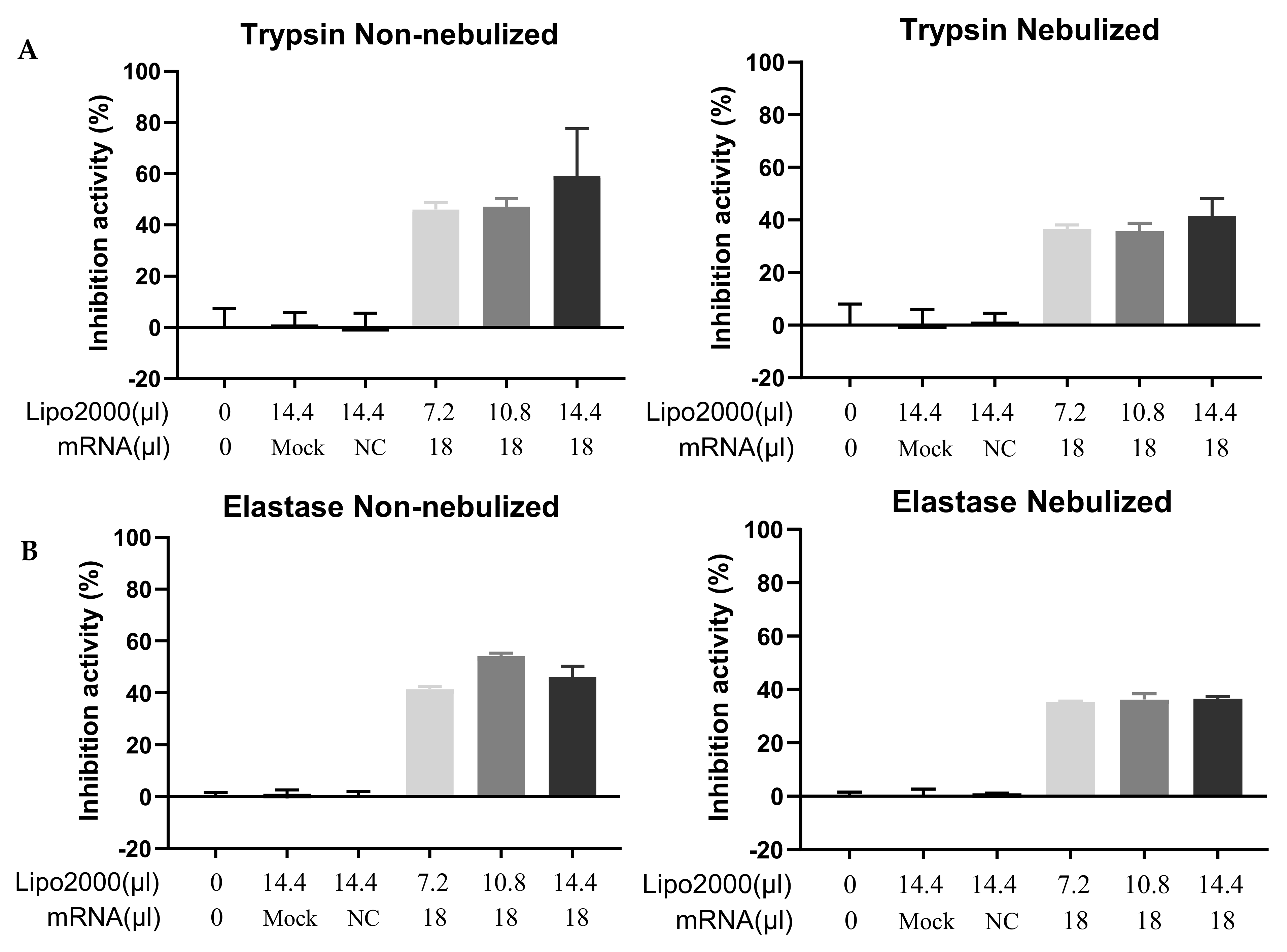
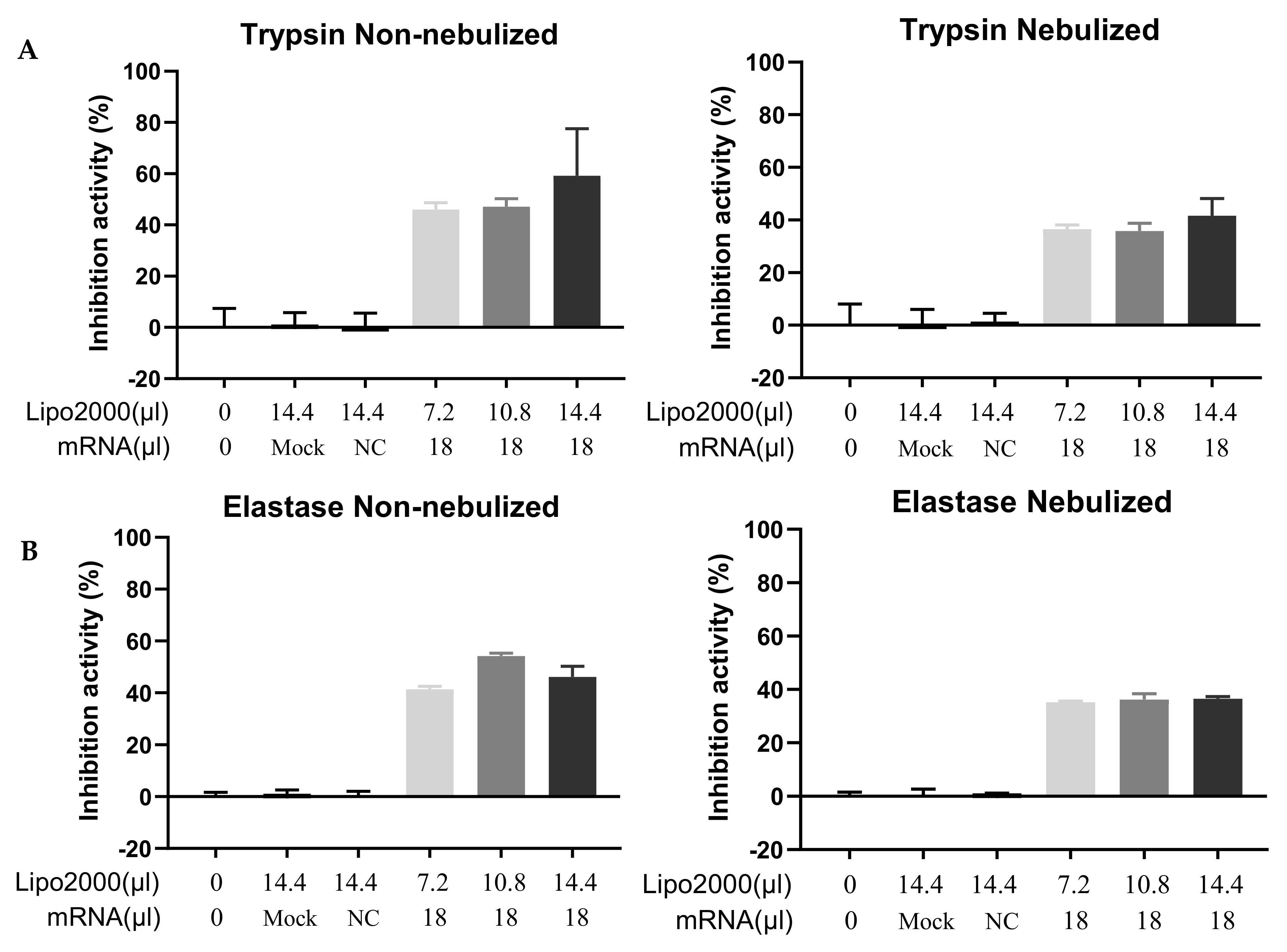
3.4. Functional Test of the Secreted A1AT
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- High, K.A.; Roncarolo, M.G. Gene therapy. N. Engl. J. Med. 2019, 381, 455–464. [Google Scholar] [CrossRef] [PubMed]
- Laczkó, D.; Hogan, M.J.; Toulmin, S.A.; Hicks, P.; Lederer, K.; Gaudette, B.T.; Castaño, D.; Amanat, F.; Muramatsu, H.; Oguin, T.H.; et al. A Single Immunization with Nucleoside-Modified mRNA Vaccines Elicits Strong Cellular and Humoral Immune Responses against SARS-CoV-2 in Mice. Immunity 2020, 53, 724–732. [Google Scholar] [CrossRef]
- Guan, S.; Rosenecker, J. Nanotechnologies in delivery of mRNA therapeutics using nonviral vector-based delivery systems. Gene Ther. 2017, 24, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Kormann, M.S.D.; Hasenpusch, G.; Aneja, M.K.; Nica, G.; Flemmer, A.W.; Herber-Jonat, S.; Huppmann, M.; Mays, L.E.; Illenyi, M.; Schams, A.; et al. Expression of therapeutic proteins after delivery of chemically modified mRNA in mice. Nat. Biotechnol. 2011, 29, 154–157. [Google Scholar] [CrossRef] [PubMed]
- Tavernier, G.; Andries, O.; Demeester, J.; Sanders, N.N.; De Smedt, S.C.; Rejman, J. mRNA as gene therapeutic: How to control protein expression. J. Control. Release 2011, 150, 238–247. [Google Scholar] [CrossRef] [PubMed]
- Karikó, K.; Buckstein, M.; Ni, H.; Weissman, D. Suppression of RNA recognition by Toll-like receptors: The impact of nucleoside modification and the evolutionary origin of RNA. Immunity 2005, 23, 165–175. [Google Scholar] [CrossRef] [Green Version]
- Karikó, K.; Muramatsu, H.; Ludwig, J.; Weissman, D. Generating the optimal mRNA for therapy: HPLC purification eliminates immune activation and improves translation of nucleoside-modified, protein-encoding mRNA. Nucleic Acids Res. 2011, 39, e142. [Google Scholar] [CrossRef] [Green Version]
- Sahin, U.; Karikó, K.; Türeci, Ö. mRNA-based therapeutics—Developing a new class of drugs. Nat. Rev. Drug Discov. 2014, 13, 759–780. [Google Scholar] [CrossRef]
- Routy, J.P.; Boulassel, M.-R.; Yassine-Diab, B.; Nicolette, C.; Healey, D.; Jain, R.; Landry, C.; Yegorov, O.; Tcherepanova, I.; Monesmith, T. Immunologic activity and safety of autologous HIV RNA-electroporated dendritic cells in HIV-1 infected patients receiving antiretroviral therapy. Clin. Immunol. 2010, 134, 140–147. [Google Scholar] [CrossRef] [Green Version]
- Kübler, H.; Scheel, B.; Gnad-Vogt, U.; Miller, K.; Schultze-Seemann, W.; Vom Dorp, F.; Parmiani, G.; Hampel, C.; Wedel, S.; Trojan, L.; et al. Self-adjuvanted mRNA vaccination in advanced prostate cancer patients: A first-in-man phase I/IIa study. J. Immunother. Cancer 2015, 3, 26. [Google Scholar] [CrossRef] [Green Version]
- Kranz, L.M.; Diken, M.; Haas, H.; Kreiter, S.; Loquai, C.; Reuter, K.C.; Meng, M.; Fritz, D.; Vascotto, F.; Hefesha, H.; et al. Systemic RNA delivery to dendritic cells exploits antiviral defence for cancer immunotherapy. Nature 2016, 534, 396–401. [Google Scholar] [CrossRef] [PubMed]
- Pardi, N.; Hogan, M.J.; Pelc, R.S.; Muramatsu, H.; Andersen, H.; DeMaso, C.R.; Dowd, K.A.; Sutherland, L.L.; Scearce, R.M.; Parks, R.; et al. Zika virus protection by a single low-dose nucleoside-modified mRNA vaccination. Nature 2017, 543, 248–251. [Google Scholar] [CrossRef]
- Khoury, H.J.; Collins, R.H.; Blum, W.; Stiff, P.S.; Elias, L.; Lebkowski, J.S.; Reddy, A.; Nishimoto, K.P.; Sen, D.; Wirth, E.D. Immune responses and long-term disease recurrence status after telomerase-based dendritic cell immunotherapy in patients with acute myeloid leukemia. Cancer 2017, 123, 3061–3072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahl, K.; Senn, J.J.; Yuzhakov, O.; Bulychev, A.; Brito, L.A.; Hassett, K.J.; Laska, M.E.; Smith, M.; Almarsson, Ö.; Thompson, J. Preclinical and Clinical Demonstration of Immunogenicity by mRNA Vaccines against H10N8 and H7N9 Influenza Viruses. Mol. Ther. J. Am. Soc. Gene Ther. 2017, 25, 1316–1327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, E.E.; Frenck, R.W.J.; Falsey, A.R.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Neuzil, K.; Mulligan, M.J.; Bailey, R.; et al. Safety and Immunogenicity of Two RNA-Based Covid-19 Vaccine Candidates. N. Engl. J. Med. 2020, 383, 2439–2450. [Google Scholar] [CrossRef]
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef]
- Beck-Broichsitter, M.; Merkel, O.M.; Kissel, T. Controlled pulmonary drug and gene delivery using polymeric nano-carriers. J. Control. Release 2012, 161, 214–224. [Google Scholar] [CrossRef]
- Katz, M.; Fargnoli, A.; Gubara, S.; Fish, K.; Weber, T.; Bridges, C.; Hajjar, R.; Ishikawa, K. Targeted Gene Delivery through the Respiratory System: Rationale for Intratracheal Gene Transfer. J. Cardiovasc. Dev. Dis. 2019, 6, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahu, I.; Haque, A.K.M.A.; Weidensee, B.; Weinmann, P.; Kormann, M.S.D. Recent Developments in mRNA-Based Protein Supplementation Therapy to Target Lung Diseases. Mol. Ther. 2019, 27, 803–823. [Google Scholar] [CrossRef] [Green Version]
- Beck, S.E.; Laube, B.L.; Barberena, C.I.; Fischer, A.C.; Adams, R.J.; Chesnut, K.; Flotte, T.R.; Guggino, W.B. Deposition and expression of aerosolized rAAV vectors in the lungs of Rhesus macaques. Mol. Ther. 2002, 6, 546–554. [Google Scholar] [CrossRef] [PubMed]
- Alton, E.W.F.W.; Armstrong, D.K.; Ashby, D.; Bayfield, K.J.; Bilton, D.; Bloomfield, E.V.; Boyd, A.C.; Brand, J.; Buchan, R.; Calcedo, R.; et al. Repeated nebulisation of non-viral CFTR gene therapy in patients with cystic fibrosis: A randomised, double-blind, placebo-controlled, phase 2b trial. Lancet Respir. Med. 2015, 3, 684–691. [Google Scholar] [CrossRef] [Green Version]
- Birchall, J.C.; Kellaway, I.W.; Gumbleton, M. Physical stability and in-vitro gene expression efficiency of nebulised lipid-peptide-DNA complexes. Int. J. Pharm. 2000, 197, 221–231. [Google Scholar] [CrossRef]
- Johler, S.M.; Rejman, J.; Guan, S.; Rosenecker, J. Nebulisation of IVT mRNA Complexes for Intrapulmonary Administration. PLoS ONE 2015, 10, e0137504. [Google Scholar] [CrossRef]
- Patel, A.K.; Kaczmarek, J.C.; Bose, S.; Kauffman, K.J.; Mir, F.; Heartlein, M.W.; DeRosa, F.; Langer, R.; Anderson, D.G. Inhaled Nanoformulated mRNA Polyplexes for Protein Production in Lung Epithelium. Adv. Mater. 2019, 31, e1805116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandhaus, R.A.; Turino, G.; Brantly, M.L.; Campos, M.; Cross, C.E.; Goodman, K.; Hogarth, D.K.; Knight, S.L.; Stocks, J.M.; Stoller, J.K.; et al. The Diagnosis and Management of Alpha-1 Antitrypsin Deficiency in the Adult. Chronic Obstr. Pulm. Dis. (Miami Fla.) 2016, 3, 668–682. [Google Scholar] [CrossRef] [Green Version]
- Brantly, M.; Nukiwa, T.; Crystal, R.G. Molecular basis of alpha-1-antitrypsin deficiency. Am. J. Med. 1988, 84, 13–31. [Google Scholar] [CrossRef]
- De Serres, F.; Blanco, I. Role of alpha-1 antitrypsin in human health and disease. J. Intern. Med. 2014, 276, 311–335. [Google Scholar] [CrossRef]
- Santangelo, S.; Scarlata, S.; Poeta, M.L.; Bialas, A.J.; Paone, G.; Incalzi, R.A. Alpha-1 Antitrypsin Deficiency: Current Perspective from Genetics to Diagnosis and Therapeutic Approaches. Curr. Med. Chem. 2017, 24, 65–90. [Google Scholar] [CrossRef]
- Stoller, J.K. Alpha-1 antitrypsin deficiency: An underrecognized, treatable cause of COPD. Cleve. Clin. J. Med. 2016, 83, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Chapman, K.R.; Chorostowska-Wynimko, J.; Koczulla, A.R.; Ferrarotti, I.; McElvaney, N.G. Alpha 1 antitrypsin to treat lung disease in alpha 1 antitrypsin deficiency: Recent developments and clinical implications. Int. J. Chron. Obstruct. Pulmon. Dis. 2018, 13, 419–432. [Google Scholar] [CrossRef] [Green Version]
- Janciauskiene, S.M.; Bals, R.; Koczulla, R.; Vogelmeier, C.; Köhnlein, T.; Welte, T. The discovery of α1-antitrypsin and its role in health and disease. Respir. Med. 2011, 105, 1129–1139. [Google Scholar] [CrossRef] [Green Version]
- McElvaney, O.J.; McEvoy, N.L.; McElvaney, O.F.; Carroll, T.P.; Murphy, M.P.; Dunlea, D.M.; Ní Choileáin, O.; Clarke, J.; O’Connor, E.; Hogan, G.; et al. Characterization of the Inflammatory Response to Severe COVID-19 Illness. Am. J. Respir. Crit. Care Med. 2020, 202, 812–821. [Google Scholar] [CrossRef] [PubMed]
- De Loyola, M.B.; Dos Reis, T.T.A.; de Oliveira, G.X.L.M.; da Fonseca Palmeira, J.; Argañaraz, G.A.; Argañaraz, E.R. Alpha-1-antitrypsin: A possible host protective factor against Covid-19. Rev. Med. Virol. 2020, 31, e2157. [Google Scholar] [CrossRef]
- Strange, C. Anti-Proteases and Alpha-1 Antitrypsin Augmentation Therapy. Respir. Care 2018, 63, 690–698. [Google Scholar] [CrossRef] [PubMed]
- Wewers, M.D.; Crystal, R.G. Alpha-1 antitrypsin augmentation therapy. COPD 2013, 10 (Suppl. 1), 64–67. [Google Scholar] [CrossRef] [PubMed]
- Chiuchiolo, M.J.; Crystal, R.G. Gene Therapy for Alpha-1 Antitrypsin Deficiency Lung Disease. Ann. Am. Thorac. Soc. 2016, 13 (Suppl. 4), S352–S369. [Google Scholar] [CrossRef] [Green Version]
- Connolly, B.; Isaacs, C.; Cheng, L.; Asrani, K.H.; Subramanian, R.R. SERPINA1 mRNA as a Treatment for Alpha-1 Antitrypsin Deficiency. J. Nucleic Acids 2018, 2018, 8247935. [Google Scholar] [CrossRef]
- Rejman, J.; Tavernier, G.; Bavarsad, N.; Demeester, J.; De Smedt, S.C. mRNA transfection of cervical carcinoma and mesenchymal stem cells mediated by cationic carriers. J. Control. Release 2010, 147, 385–391. [Google Scholar] [CrossRef]
- Choi, Y.J.; Kim, S.; Choi, Y.; Nielsen, T.B.; Yan, J.; Lu, A.; Ruan, J.; Lee, H.-R.; Wu, H.; Spellberg, B.; et al. SERPINB1-mediated checkpoint of inflammatory caspase activation. Nat. Immunol. 2019, 20, 276–287. [Google Scholar] [CrossRef]
- Callaghan, P.J.; Ferrick, B.; Rybakovsky, E.; Thomas, S.; Mullin, J.M. Epithelial barrier function properties of the 16HBE14o- human bronchial epithelial cell culture model. Biosci. Rep. 2020, 40, BSR20201532. [Google Scholar] [CrossRef]
- Guan, S.; Munder, A.; Hedtfeld, S.; Braubach, P.; Glage, S.; Zhang, L.; Lienenklaus, S.; Schultze, A.; Hasenpusch, G.; Garrels, W.; et al. Self-assembled peptide–poloxamine nanoparticles enable in vitro and in vivo genome restoration for cystic fibrosis. Nat. Nanotechnol. 2019, 14, 287–297. [Google Scholar] [CrossRef]
- Dalby, B.; Cates, S.; Harris, A.; Ohki, E.C.; Tilkins, M.L.; Price, P.J.; Ciccarone, V.C. Advanced transfection with Lipofectamine 2000 reagent: Primary neurons, siRNA, and high-throughput applications. Methods 2004, 33, 95–103. [Google Scholar] [CrossRef]
- De Haes, W.; Rejman, J.; Pollard, C.; Merlin, C.; Vekemans, M.; Florence, E.; De Smedt, S.C.; Grooten, J.; Vanham, G.; De Koker, S.; et al. Lipoplexes carrying mRNA encoding Gag protein modulate dendritic cells to stimulate HIV-specific immune responses. Nanomedicine 2013, 8, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Rejman, J.; Oberle, V.; Zuhorn, I.S.; Hoekstra, D. Size-dependent internalization of particles via the pathways of clathrin- and caveolae-mediated endocytosis. Biochem. J. 2004, 377, 159–169. [Google Scholar] [CrossRef]
- Breunig, M.; Lungwitz, U.; Liebl, R.; Goepferich, A. Breaking up the correlation between efficacy and toxicity for nonviral gene delivery. Proc. Natl. Acad. Sci. USA 2007, 104, 14454–14459. [Google Scholar] [CrossRef] [Green Version]
- Ernst, N.; Ulrichskötter, S.; Schmalix, W.A.; Rädler, J.; Galneder, R.; Mayer, E.; Gersting, S.; Plank, C.; Reinhardt, D.; Rosenecker, J. Interaction of liposomal and polycationic transfection complexes with pulmonary surfactant. J. Gene Med. 1999, 1, 331–340. [Google Scholar] [CrossRef]
- Densmore, C.L.; Orson, F.M.; Xu, B.; Kinsey, B.M.; Waldrep, J.C.; Hua, P.; Bhogal, B.; Knight, V. Aerosol delivery of robust polyethyleneimine-DNA complexes for gene therapy and genetic immunization. Mol. Ther. 2000, 1, 180–188. [Google Scholar] [CrossRef]
- Rudolph, C.; Ortiz, A.; Schillinger, U.; Jauernig, J.; Plank, C.; Rosenecker, J. Methodological optimization of polyethylenimine (PEI)-based gene delivery to the lungs of mice via aerosol application. J. Gene Med. 2005, 7, 59–66. [Google Scholar] [CrossRef]
- Ari, A. Jet, Ultrasonic, and Mesh Nebulizers: An Evaluation of Nebulizers for Better Clinical Outcomes. Eurasian J. Pulmonol. 2014, 16, 1–7. [Google Scholar] [CrossRef]
- Abdulkarim, A.; Craig, T.J. Alpha 1 Antitrypsin Mutation. In Treasure Island (Florida, USA); StatPearls Publishing: Treasure Island, FL, USA, 2018. [Google Scholar]
- Koga, H.; Miyahara, N.; Fuchimoto, Y.; Ikeda, G.; Waseda, K.; Ono, K.; Tanimoto, Y.; Kataoka, M.; Gelfand, E.W.; Tanimoto, M.; et al. Inhibition of neutrophil elastase attenuates airway hyperresponsiveness and inflammation in a mouse model of secondary allergen challenge: Neutrophil elastase inhibition attenuates allergic airway responses. Respir. Res. 2013, 14, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mecham, R.P. Elastin in lung development and disease pathogenesis. Matrix Biol. 2018, 73, 6–20. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IVT-mRNA/Lipofectamine2000 Lipoplexes | Amounts of Lipofectamine2000 (µL) | Amounts of IVT-mRNA (µL) | Size (nm) Non-Nebulized | Size (nm) Nebulized |
|---|---|---|---|---|
| 1st Setting | 2.4 | 6.0 | 397 ± 118 | 649 ± 173 |
| 1st Setting | 3.6 | 6.0 | 467 ± 67 | 543 ± 123 |
| 1st Setting | 4.8 | 6.0 | 324 ± 46 | 628 ± 76 |
| 2nd Setting | 1.2 | 2.0 | 458 ± 89 | 737 ± 113 |
| 2nd Setting | 2.4 | 4.0 | 480 ± 74 | 676 ± 138 |
| 2nd Setting | 3.6 | 6.0 | 378 ± 84 | 586 ± 161 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guan, S.; Darmstädter, M.; Xu, C.; Rosenecker, J. In Vitro Investigations on Optimizing and Nebulization of IVT-mRNA Formulations for Potential Pulmonary-Based Alpha-1-Antitrypsin Deficiency Treatment. Pharmaceutics 2021, 13, 1281. https://doi.org/10.3390/pharmaceutics13081281
Guan S, Darmstädter M, Xu C, Rosenecker J. In Vitro Investigations on Optimizing and Nebulization of IVT-mRNA Formulations for Potential Pulmonary-Based Alpha-1-Antitrypsin Deficiency Treatment. Pharmaceutics. 2021; 13(8):1281. https://doi.org/10.3390/pharmaceutics13081281
Chicago/Turabian StyleGuan, Shan, Max Darmstädter, Chuanfei Xu, and Joseph Rosenecker. 2021. "In Vitro Investigations on Optimizing and Nebulization of IVT-mRNA Formulations for Potential Pulmonary-Based Alpha-1-Antitrypsin Deficiency Treatment" Pharmaceutics 13, no. 8: 1281. https://doi.org/10.3390/pharmaceutics13081281
APA StyleGuan, S., Darmstädter, M., Xu, C., & Rosenecker, J. (2021). In Vitro Investigations on Optimizing and Nebulization of IVT-mRNA Formulations for Potential Pulmonary-Based Alpha-1-Antitrypsin Deficiency Treatment. Pharmaceutics, 13(8), 1281. https://doi.org/10.3390/pharmaceutics13081281