Synchronous Langat Virus Infection of Haemaphysalis longicornis Using Anal Pore Microinjection

Abstract
:1. Introduction
2. Materials and Methods
2.1. Ticks and Animals
2.2. Cells and Virus
2.3. Tick Infection
2.4. Detection of Langat Virus RNA
2.5. Langat Virus Titration Among LGTV-Infected Adult Ticks
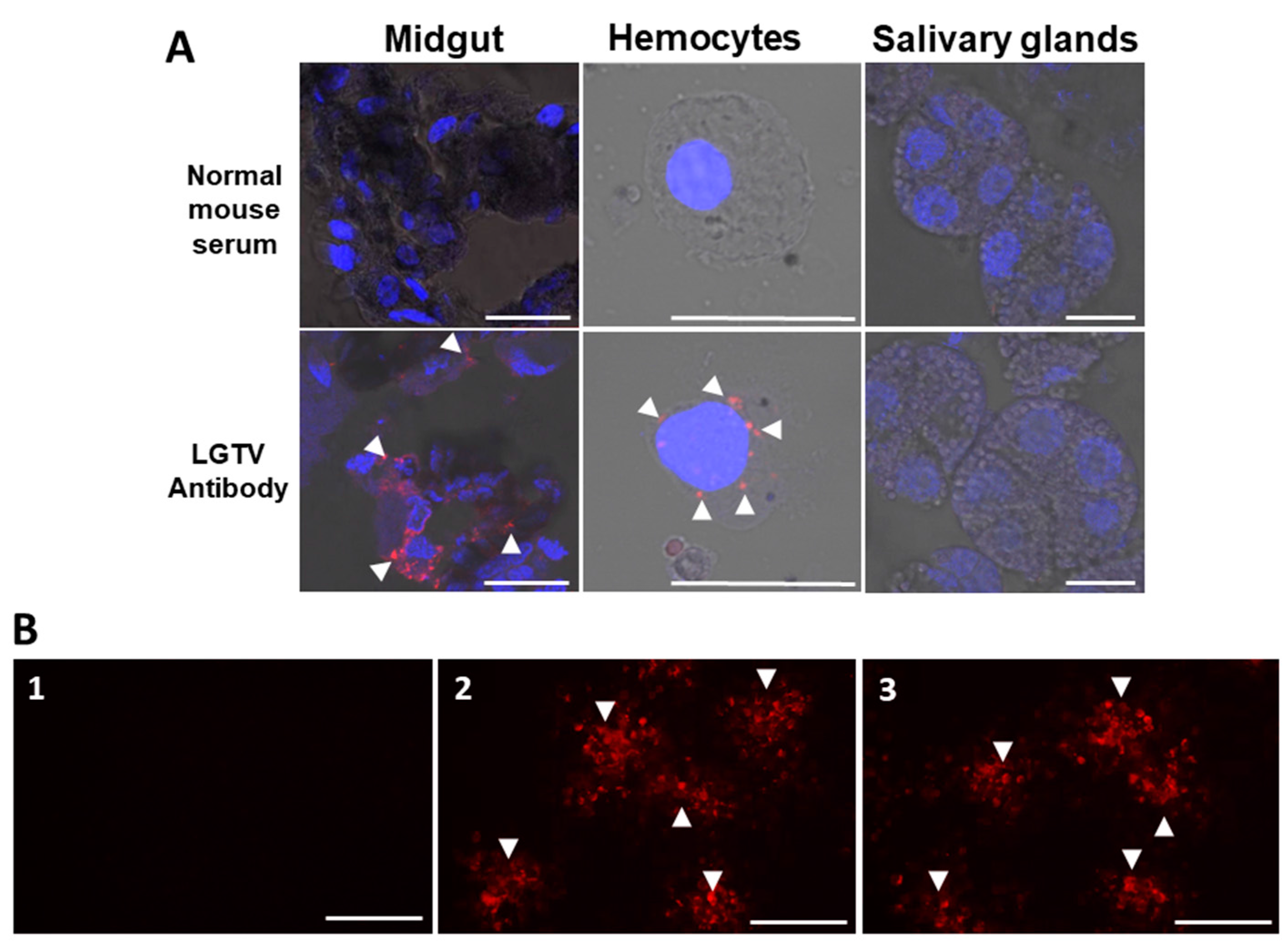
2.6. Detection of Langat Virus Antigens in Tick Organs Using Indirect Immunofluorescent Antibody Test
2.7. Langat Virus Transmission from Ticks to Mice
2.8. Transovarial Transmission of Langat Virus in Ticks
2.9. Statistical Analysis
3. Results
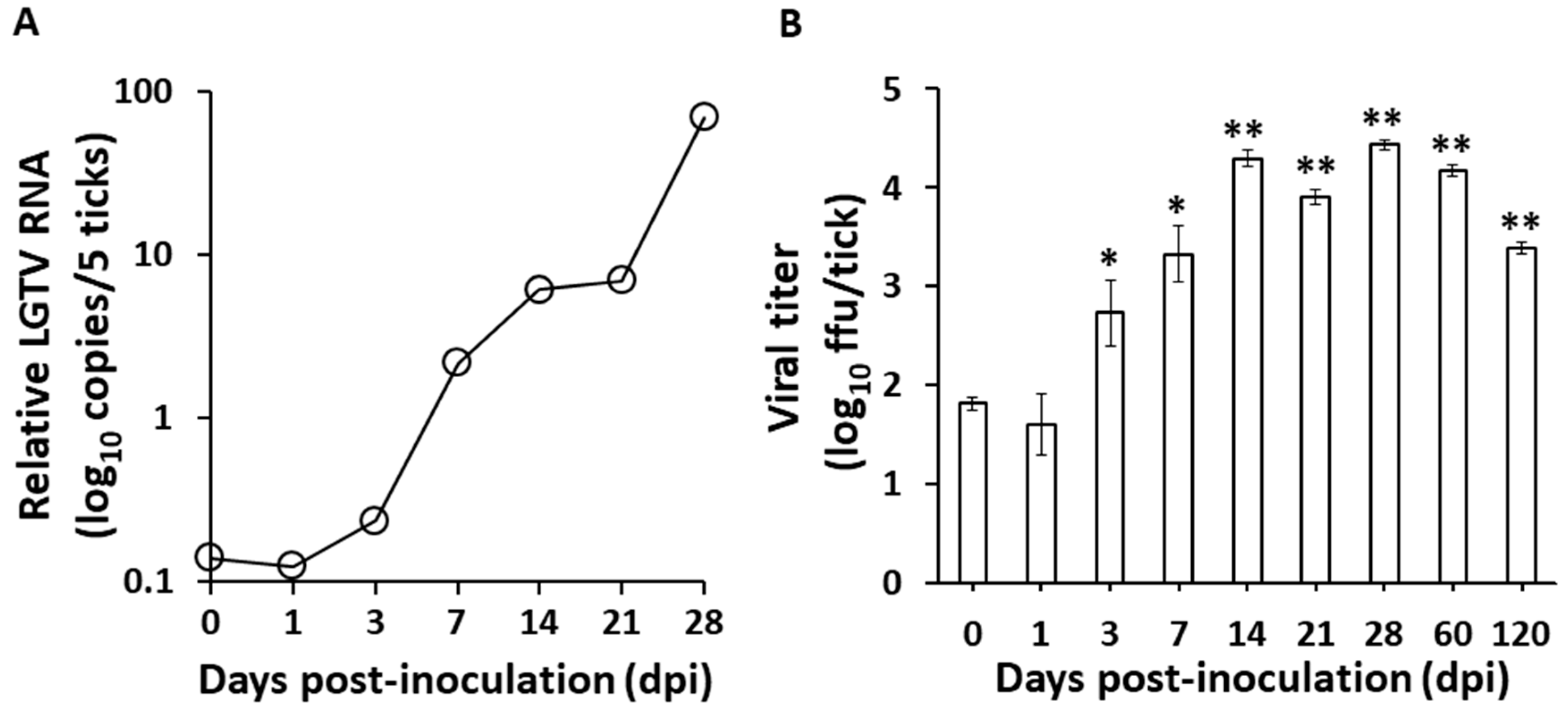
3.1. Langat Virus Infection in Ticks
3.2. Langat Virus Transmission to Mice
3.3. Transovarial Transmission of Langat Virus in Ticks
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- de la Fuente, J.; Estrada-Pena, A.; Venzal, J.M.; Kocan, K.M.; Sonenshine, D.E. Overview: Ticks as vectors of pathogens that cause disease in humans and animals. Front. Biosci. 2008, 13, 6938–6946. [Google Scholar] [CrossRef] [PubMed]
- Mitzel, D.N.; Wolfinbarger, J.B.; Long, R.D.; Masnick, M.; Best, S.M.; Bloom, M.E. Tick-borne flavivirus infection in Ixodes scapularis larvae: Development of a novel method for synchronous viral infection of ticks. Virology 2007, 365, 410–418. [Google Scholar] [CrossRef] [PubMed]
- Lehrer, A.T.; Holbrook, M.R. Tick-borne Encephalitis Vaccines. J. Bioterror. Biodef. 2011, 2011 (Suppl. S1), 003. [Google Scholar] [CrossRef] [PubMed]
- Turell, M.J.; Durden, L.A. Experimental transmission of Langat (Tick-Borne Encephalitis complex) virus by the soft tick Ornithodoros sonrai (Acari: Argasidae). J. Med. Entomol. 1994, 31, 148–151. [Google Scholar] [CrossRef] [PubMed]
- Hoogstraal, H.; Roberts, F.H.; Kohls, G.M.; Tipton, V.J. Review of Haemaphysalis (kaiseriana) longicornis Neumann (resurrected) of Australia, New Zealand, New Caledonia, Fiji, Japan, Korea, and Northeastern China and USSR, and its parthenogenetic and bisexual populations (Ixodoidea, Ixodidae). J. Parasitol. 1968, 54, 1197–1213. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, T. Bovine piroplasmosis in Japan. Jpn. Agric. Res. Q. 1968, 3, 23–31. [Google Scholar]
- Luo, L.M.; Zhao, L.; Wen, H.L.; Zhang, Z.T.; Liu, J.W.; Fang, L.Z.; Xue, Z.F.; Ma, D.Q.; Zhang, X.S.; Ding, S.J.; Lei, X.Y.; Yu, X.J. Haemaphysalis longicornis ticks as reservoir and vector of severe fever with thrombocytopenia syndrome virus in China. Emerg. Infect. Dis. 2015, 21, 1770–1776. [Google Scholar] [CrossRef] [PubMed]
- Hoogstraal, H. Changing patterns of tick-borne diseases in modern society. Annu. Rev. Entomol. 1981, 26, 75–99. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Jeong, Y.E.; Yun, S.M.; Lee, I.Y.; Han, M.G.; Ju, Y.R. Molecular evidence for tick-borne encephalitis virus in ticks in South Korea. Med. Vet. Entomol. 2009, 23, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Chernesky, M.A. Powassan virus transmission by ixodid ticks infected after feeding on viremic rabbits injected intravenously. Can. J. Microbiol. 1969, 15, 521–526. [Google Scholar] [CrossRef] [PubMed]
- Steele, G.M.; Nuttall, P.A. Difference in vector competence of two species of sympatric ticks, Amblyomma variegatum and Rhipicephalus appendiculatus, for Dugbe virus (Nairovirus, Bunyaviridae). Virus Res. 1989, 14, 73–84. [Google Scholar] [CrossRef]
- Kariu, T.; Coleman, A.S.; Anderson, J.F.; Pal, U. Methods for rapid transfer and localization of lyme disease pathogens within the tick gut. J. Vis. Exp. 2011, 48, 2544. [Google Scholar] [CrossRef] [PubMed]
- Zandi, K.; Teoh, B.T.; Sam, S.S.; Wong, P.F.; Mustafa, M.R.; Abubakar, S. In vitro antiviral activity of fisetin, rutin and naringenin against dengue virus type-2. J. Med. Plants Res. 2011, 23, 5534–5539. [Google Scholar]
- Takechi, R.; Galay, R.L.; Matsuo, T.; Maeda, H.; Kusakisako, K.; Talactac, M.R.; Mochizuki, M.; Fujisaki, K.; Tanaka, T. Role of the tumor necrosis factor receptor-associated factor-type zinc finger domain containing protein 1 (TRAFD1) from the hard tick Haemaphysalis longicornis in immunity against bacterial infection. Ticks Tick-Borne Dis. 2016, 7, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Ayers, M.; Adachi, D.; Johnson, G.; Andonova, M.; Drebot, M.; Tellier, R. A single tube RT-PCR assay for the detection of mosquito-borne flaviviruses. J. Virol. Methods. 2006, 135, 235–239. [Google Scholar] [CrossRef] [PubMed]
- Maeda, H.; Miyata, T.; Kusakisako, K.; Galay, R.L.; Talactac, M.R.; Umemiya-Shirafuj, R.; Mochizuki, M.; Fujisaki, K.; Tanaka, T. A novel C-type lectin with triple carbohydrate recognition domains has critical roles for the hard tick Haemaphysalis longicornis against Gram-negative bacteria. Dev. Comp. Immunol. 2015, 57, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Aung, K.M.; Boldbaatar, D.; Liao, M.; Umemiya-Shirafuji, R.; Nakao, S.; Matsuoka, T.; Tanaka, T.; Fujisaki, K. Identification and characterization of class B scavenger receptor CD36 from the hard tick, Haemaphysalis longicornis. Parasitol. Res. 2011, 108, 273–285. [Google Scholar] [CrossRef] [PubMed]
- Umemiya, R.; Matsuo, T.; Hatta, T.; Sakakibara, S.; Boldbaatar, D.; Fujisaki, K. Cloning and characterization of an autophagy-related gene, ATG12, from the three-host tick Haemaphysalis longicornis. Insect Biochem. Mol. Biol. 2007, 37, 975–984. [Google Scholar] [CrossRef] [PubMed]
- Anisuzzamman, I.M.; Alim, M.; Miyoshi, T.; Hatta, T.; Yamaji, K.; Matsumoto, Y.; Fujisaki, K.; Tsuji, N. Longistatin is an unconventional serine protease and induces protective immunity against tick infestation. Mol. Biochem. Parasitol. 2012, 182, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Holbrook, M.R.; Aronson, J.F.; Campbell, G.A.; Jones, S.; Feldmann, H.; Barrett, A.D. An animal model for the tickborne flavivirus-Omsk hemorrhagic fever virus. J. Infect. Dis. 2005, 191, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Seamer, J.; Randles, W.J. The course of Langat virus infection in mice. Br. J. Exp. Pathol. 1967, 48, 403–410. [Google Scholar] [PubMed]
- Labuda, M.; Nuttall, P.A. Tick-borne viruses. Parasitology 2004, 129, S221–S245. [Google Scholar] [CrossRef] [PubMed]
- Booth, T.F.; Davies, C.R.; Jones, L.D.; Staunton, D.; Nuttall, P.A. Anatomical basis of Thogoto virus infection in BHK cell culture and the ixodid tick vector, Rhipicephalus appendiculatus. J. Gen. Virol. 1989, 70, 1093–1104. [Google Scholar] [CrossRef] [PubMed]
- Lindblom, P.; Wilhelmsson, P.; Fryland, L.; Sjöwall, J.; Haglund, M.; Matussek, A.; Ernerudh, J.; Vene, S.; Nyman, D.; Andreassen, A. Tick-borne encephalitis virus in ticks detached from humans and follow-up of serological and clinical response. Ticks Tick-Borne Dis. 2013, 5, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, W.R.; Nuttall, P.A. Rhipicephalus appendiculatus (Acari: Ixodidae): Dynamics of Thogoto virus infection in female ticks during feeding on guinea pigs. Exp. Parasitol. 2003, 104, 20–25. [Google Scholar] [CrossRef]
- Kozuch, O.; Labuda, M.; Lysy, J.; Weismann, P.; Krippel, E. Longitudinal study of natural foci of central European encephalitis virus in west Slovakia. Acta Virol. 1990, 34, 537–544. [Google Scholar] [PubMed]
- Randolph, S.E. Transmission of tick-borne pathogens between co-feeding ticks: Milan Labuda’s enduring paradigm. Ticks Tick-Borne Dis. 2011, 2, 179–182. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| Primer Name | Primer Sequence |
|---|---|
| LGTV Pre-M Forward | GGATGGATTGTTGCCCAGGA |
| LGTV Pre-M Reverse | CCCAGCTCGAGAACCAATGT |
| LGTV Neg. Forward | GTCTCCGGTTGCAGGACTGT |
| LGTV Neg. Reverse | CTCGGTCAGTAGGATGGTGTTG |
| H. longicornis L23 Forward | CACACTCGTGTTCATCGTCC |
| H. longicornis L23 Reverse | ATGAGTGTGTTCACGTTGGC |
| Mouse β-actin Forward | TTCTTTGCAGCTCCTTCGTT |
| Mouse β-actin Reverse | ATGGAGGGGAATACAGCCC |
| Inoculum | LGTV Detection | |
|---|---|---|
| Absolute Value | Percentage | |
| EMEM | 0/20 | 0 |
| LGTV | 20/20 | 100% |
| Tissue | No. Positive/Total (%) | Mean Titer ± Standard Deviation (log10 ffu/Tick) |
|---|---|---|
| Midgut | 20/20 (100) | 3.37 ± 0.16 |
| Salivary Gland | 0/20 (0) | - |
| Carcass | 3/20 (15) | 2.53 ± 0.85 |
| Treatment | Moribund Mice a | Survivors | |||
|---|---|---|---|---|---|
| Mortality (%) (Death/Total) | Viral RNA in Brain b | Seroconversion (%) (Positive/Total) | Viral RNA in Brain b | Viral RNA in Blood b | |
| EMEM-injected ticks | 0 (0/5) | N.A. | 0 (0/5) | − | − |
| LGTV-injected ticks | 10 (2/20) | + (2/2) | 88.8 (16/18) | − | − |
| LGTV inoculated mice | 20 (1/5) | + (1/1) | 100 (4/4) | − | − |
| Sample | Group | LGTV Detection | |
|---|---|---|---|
| Absolute Value | Percentage | ||
| Eggs | EMEM | 0/5 | 0 |
| LGTV | 0/17 | 0 | |
| Larvae | EMEM | 0/5 | 0 |
| LGTV | 0/17 | 0 | |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Talactac, M.R.; Yoshii, K.; Hernandez, E.P.; Kusakisako, K.; Galay, R.L.; Fujisaki, K.; Mochizuki, M.; Tanaka, T. Synchronous Langat Virus Infection of Haemaphysalis longicornis Using Anal Pore Microinjection. Viruses 2017, 9, 189. https://doi.org/10.3390/v9070189
Talactac MR, Yoshii K, Hernandez EP, Kusakisako K, Galay RL, Fujisaki K, Mochizuki M, Tanaka T. Synchronous Langat Virus Infection of Haemaphysalis longicornis Using Anal Pore Microinjection. Viruses. 2017; 9(7):189. https://doi.org/10.3390/v9070189
Chicago/Turabian StyleTalactac, Melbourne Rio, Kentaro Yoshii, Emmanuel Pacia Hernandez, Kodai Kusakisako, Remil Linggatong Galay, Kozo Fujisaki, Masami Mochizuki, and Tetsuya Tanaka. 2017. "Synchronous Langat Virus Infection of Haemaphysalis longicornis Using Anal Pore Microinjection" Viruses 9, no. 7: 189. https://doi.org/10.3390/v9070189
APA StyleTalactac, M. R., Yoshii, K., Hernandez, E. P., Kusakisako, K., Galay, R. L., Fujisaki, K., Mochizuki, M., & Tanaka, T. (2017). Synchronous Langat Virus Infection of Haemaphysalis longicornis Using Anal Pore Microinjection. Viruses, 9(7), 189. https://doi.org/10.3390/v9070189