
Factors That Improve RT-QuIC Detection of Prion Seeding Activity


 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Recombinant Prion Protein Purification
2.2. RT-QuIC
3. Results
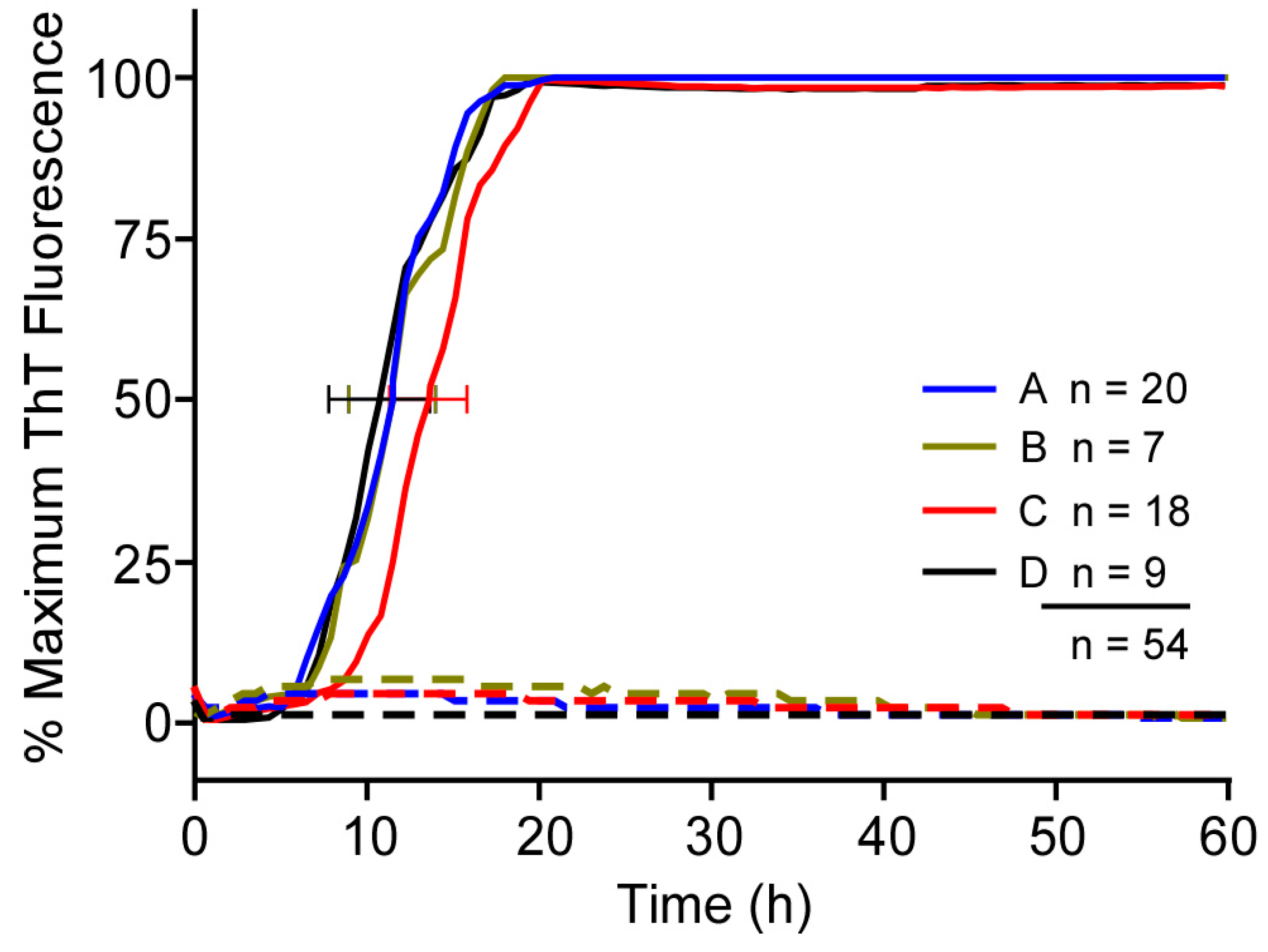
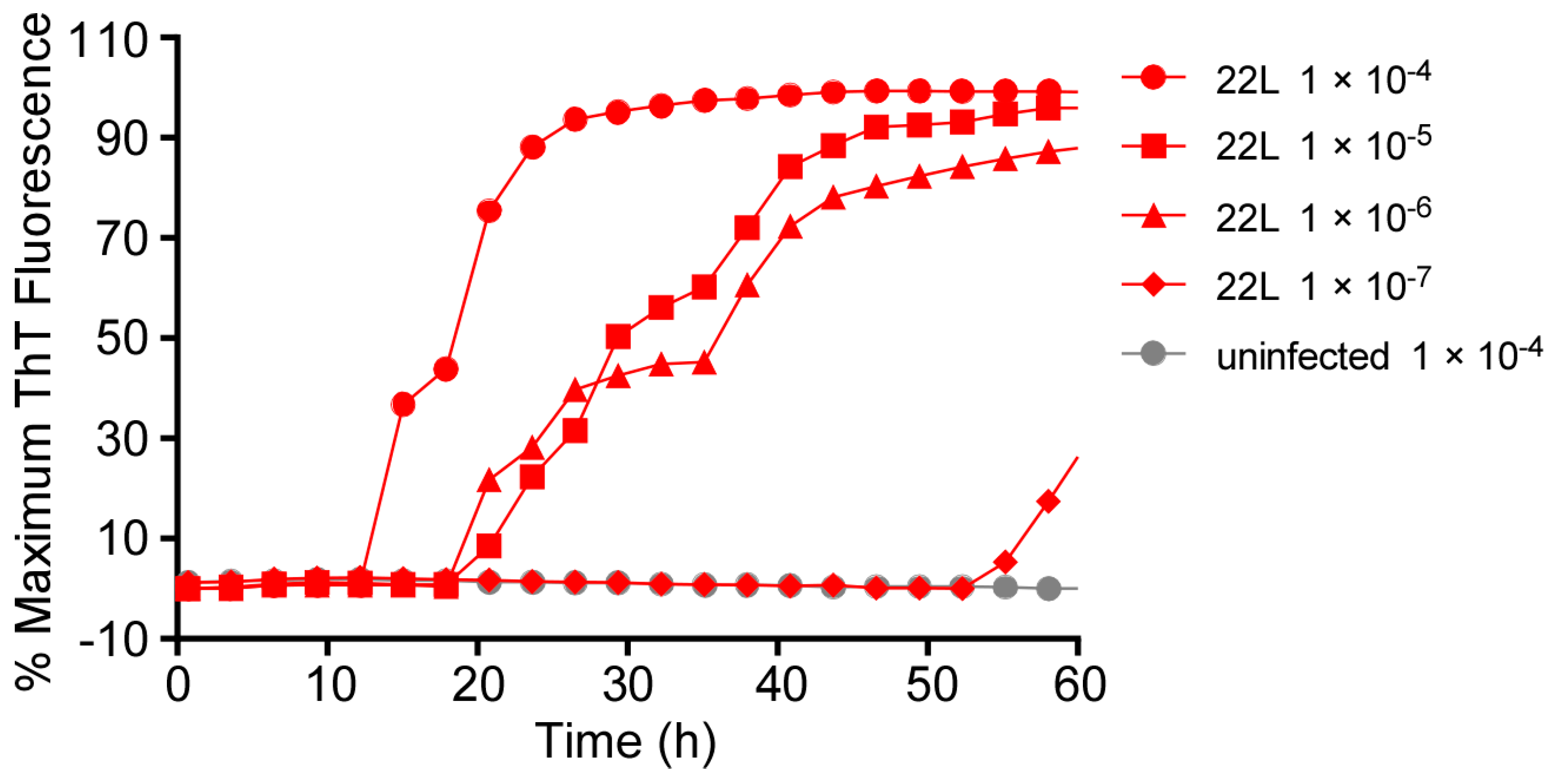
3.1. Comparison of RT-QuIC Kinetics Using Multiple Batches of a Given Type of Recombinant PrP Substrate
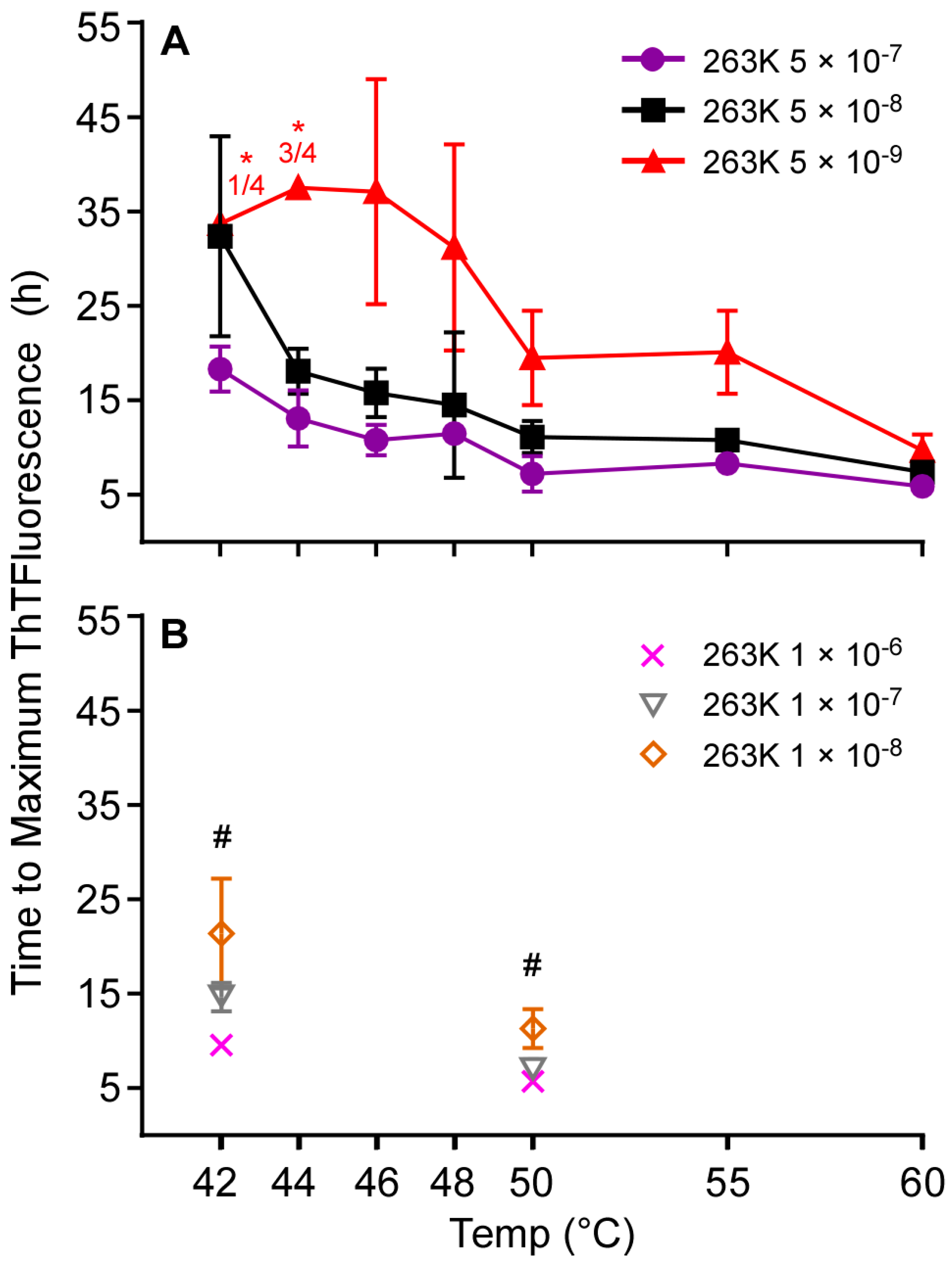
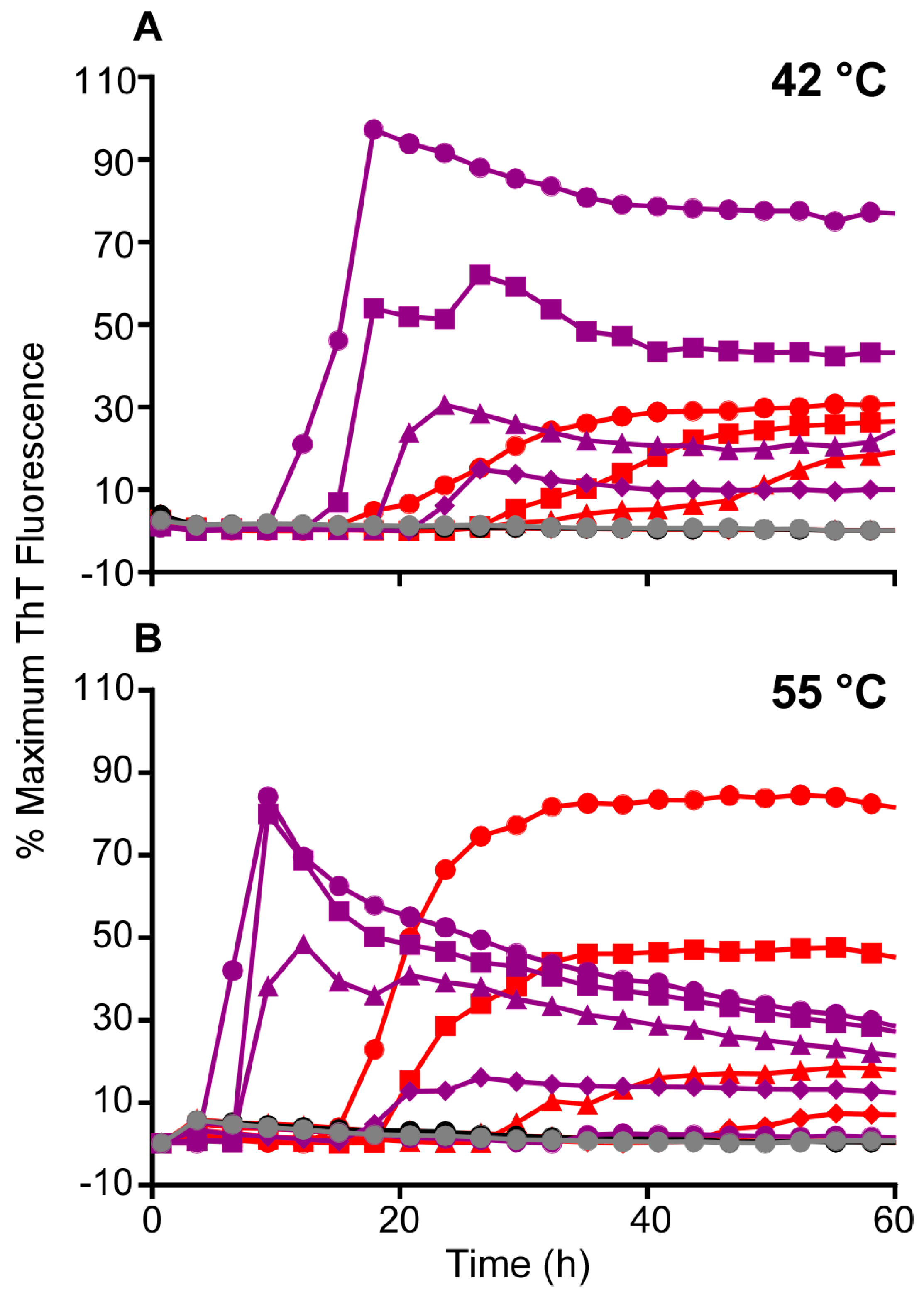
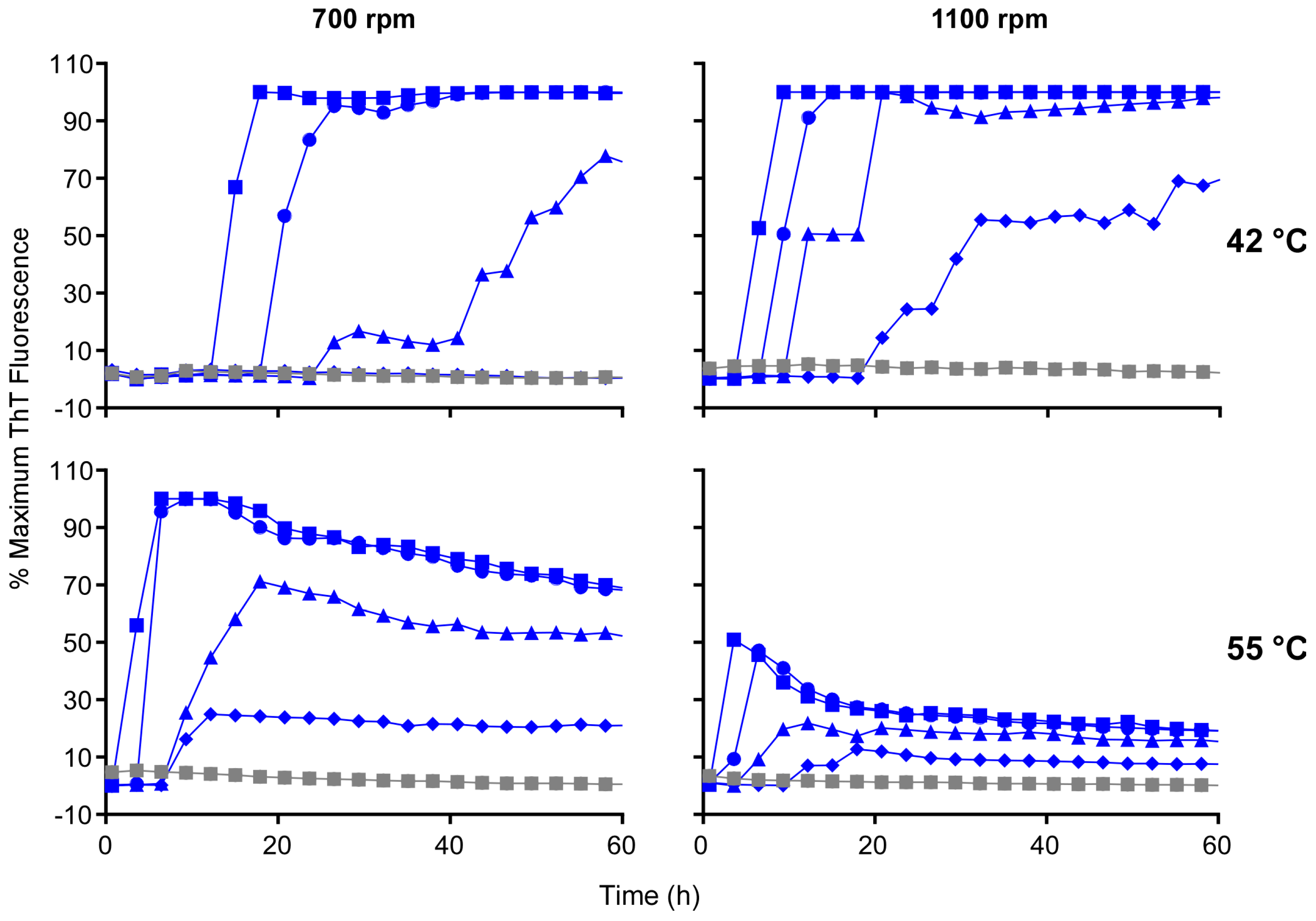
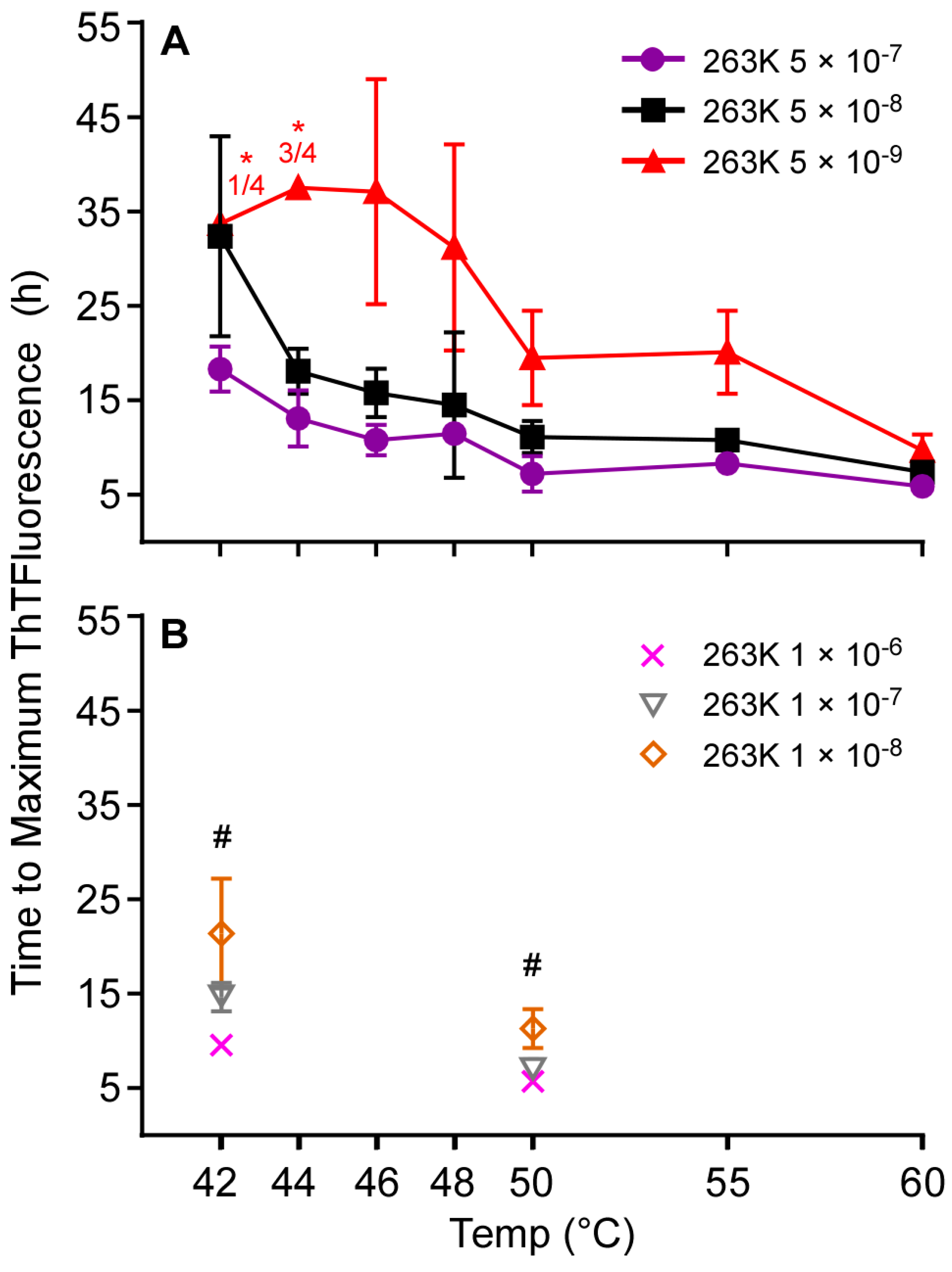
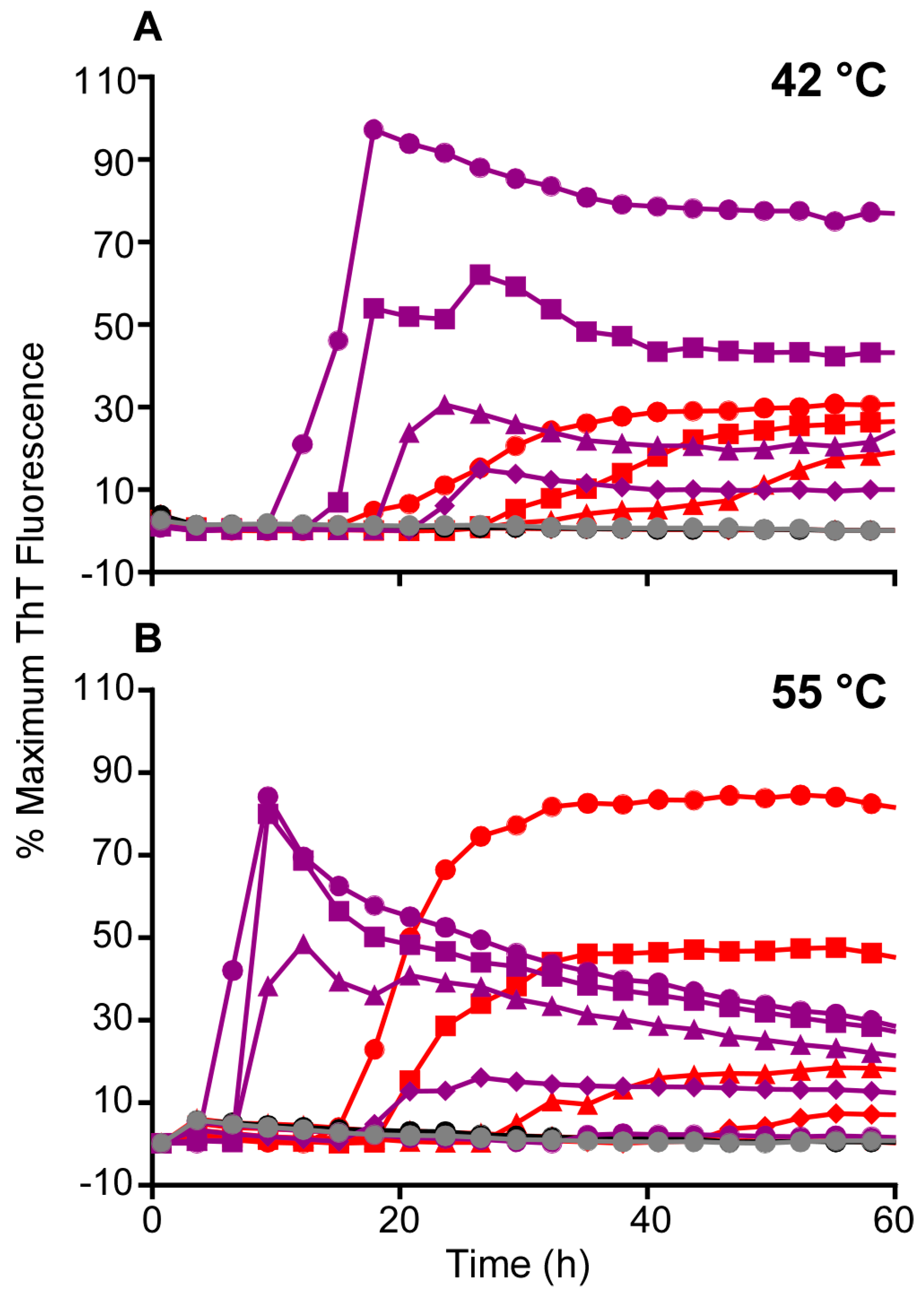
3.2. Higher Temperatures Promote Faster RT-QuIC Reactions
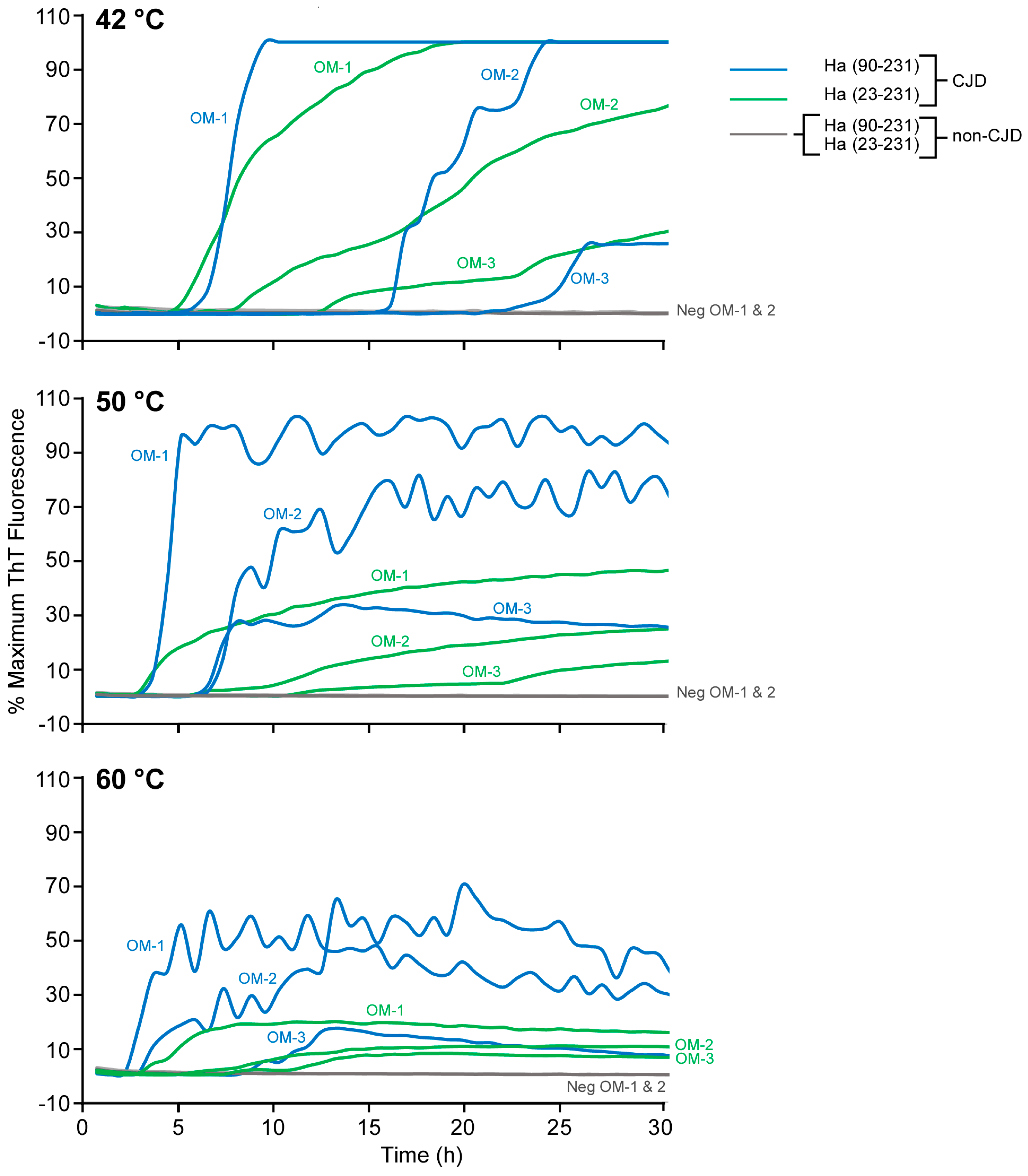
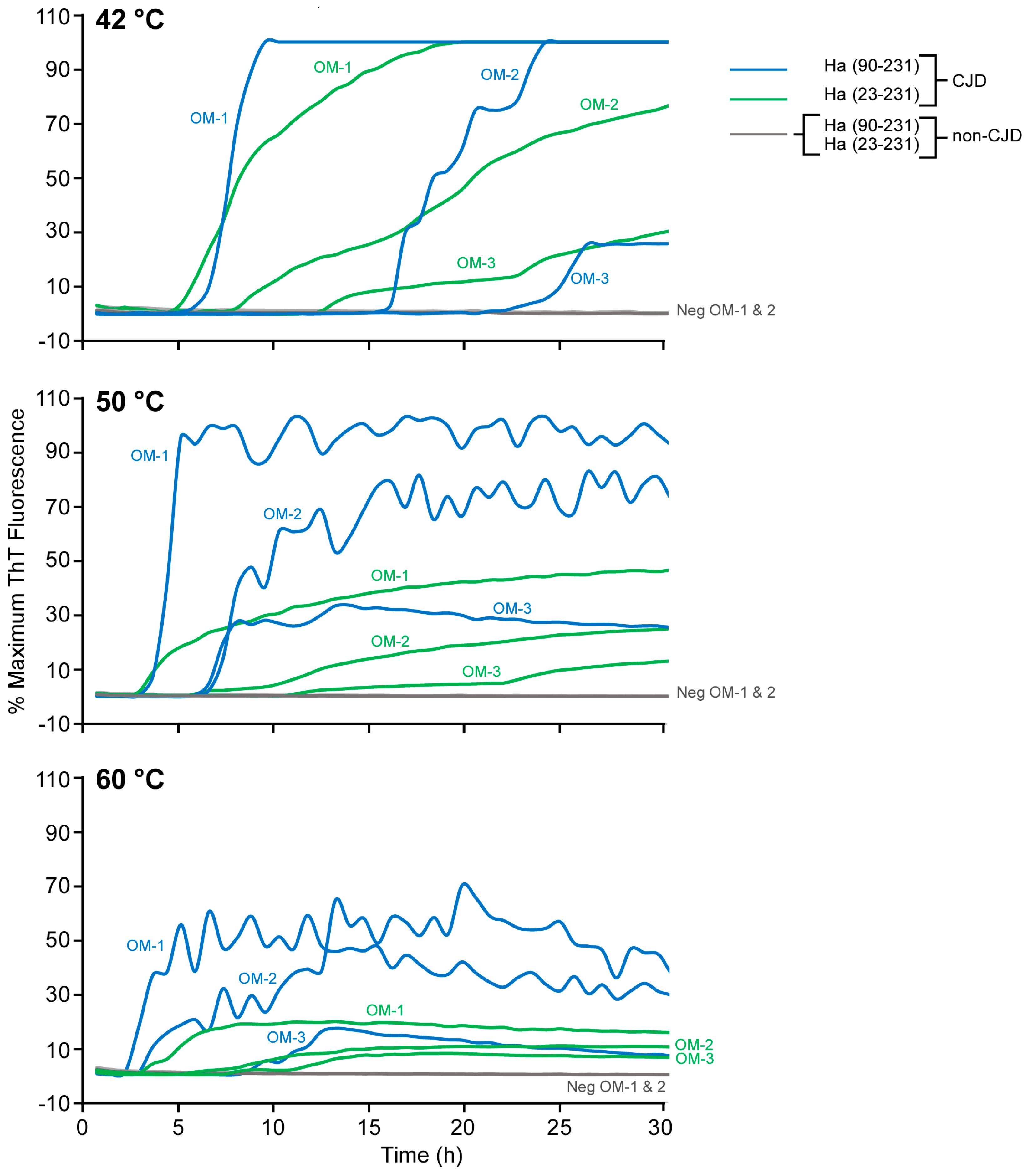
3.3. Improved Detection of Human sCJD in Nasal Brushings
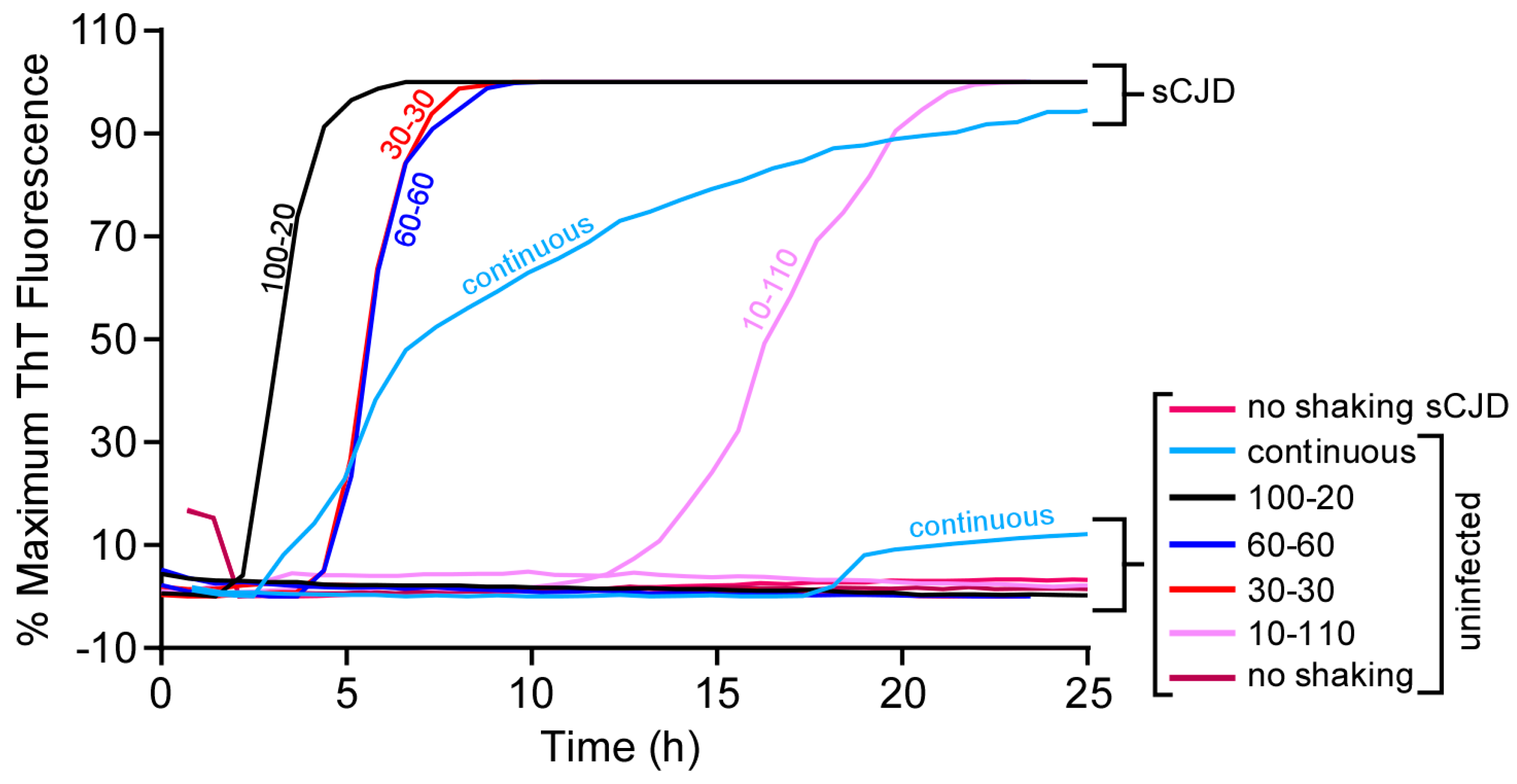
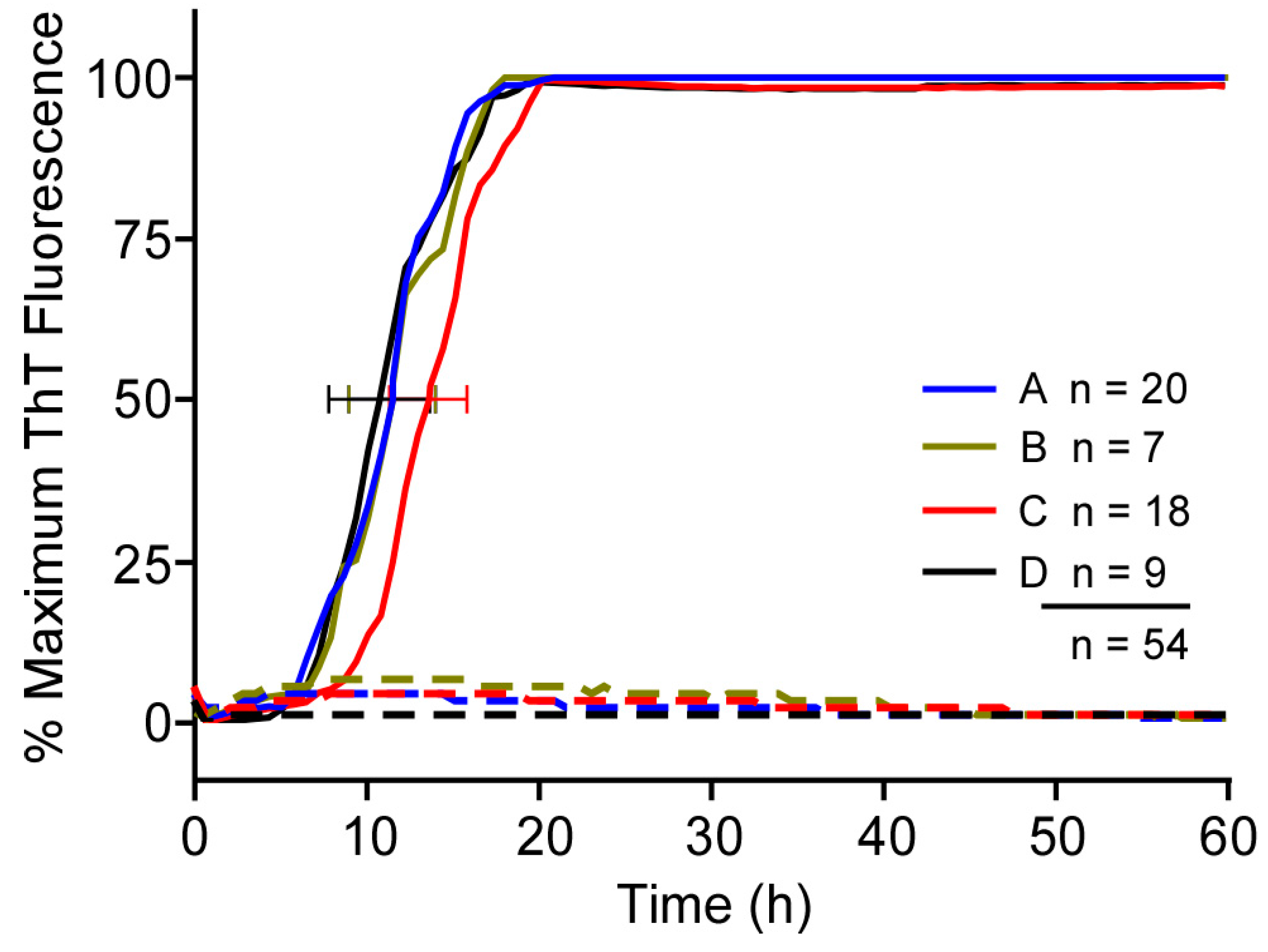
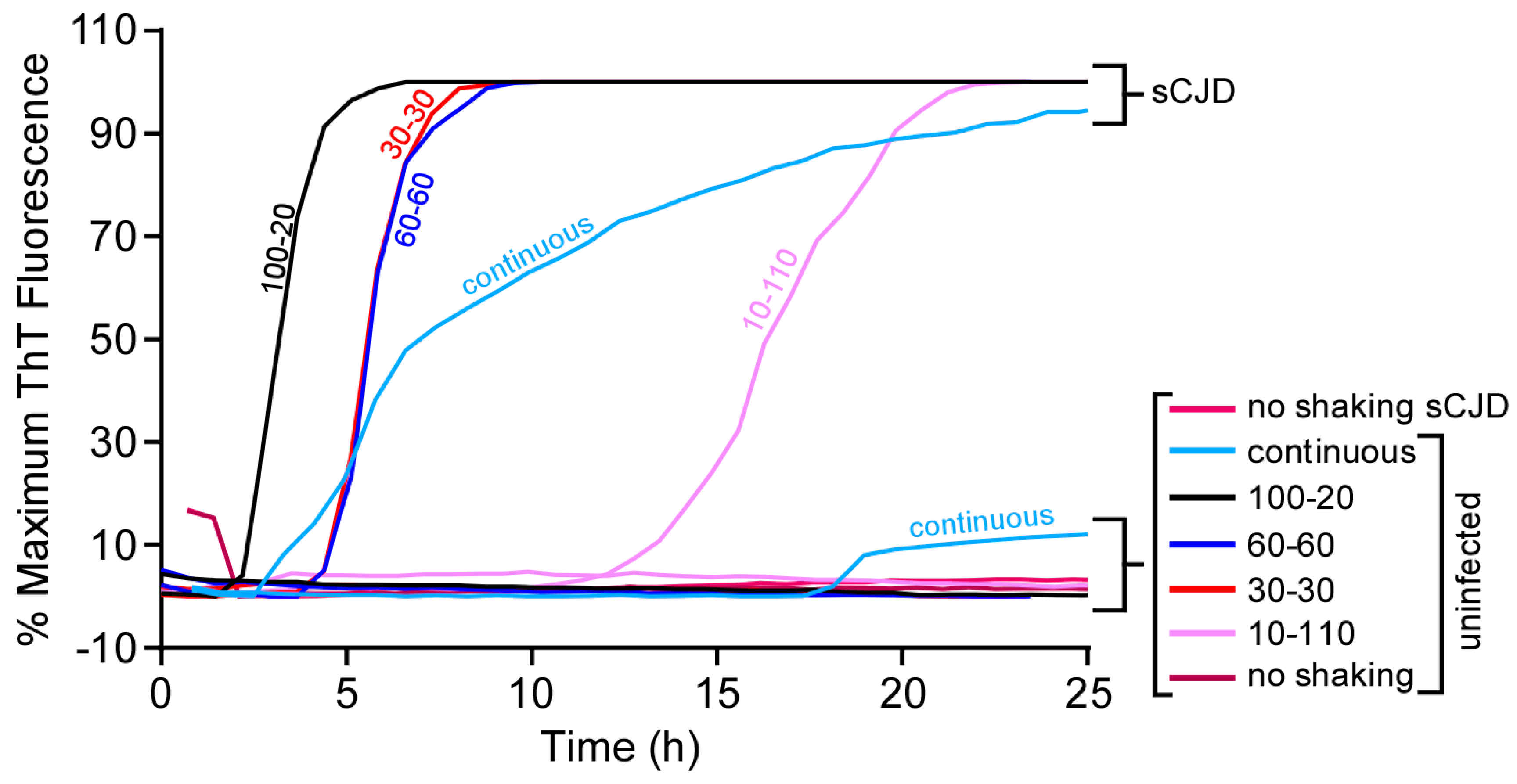
3.4. Longer Shaking Intervals Promote Faster RT-QuIC Kinetics
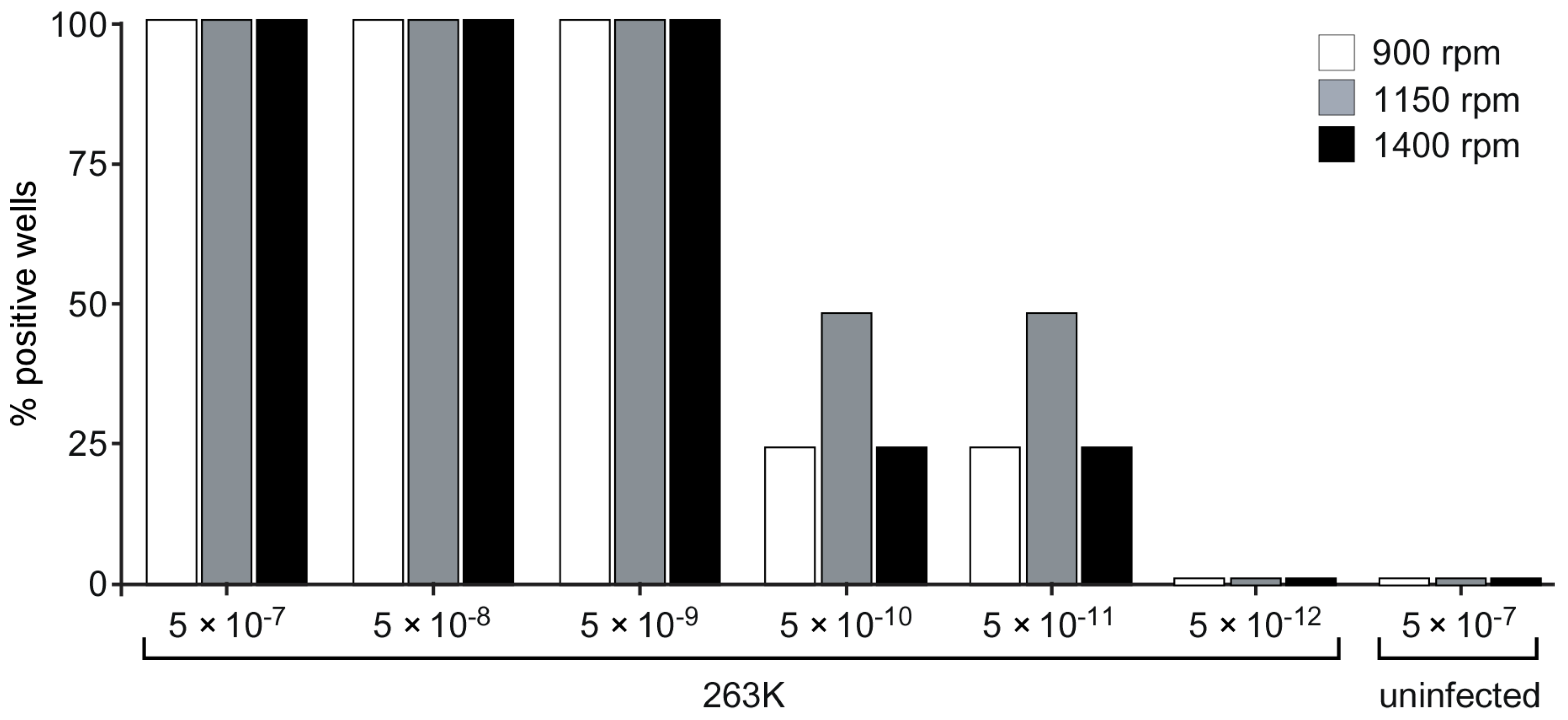
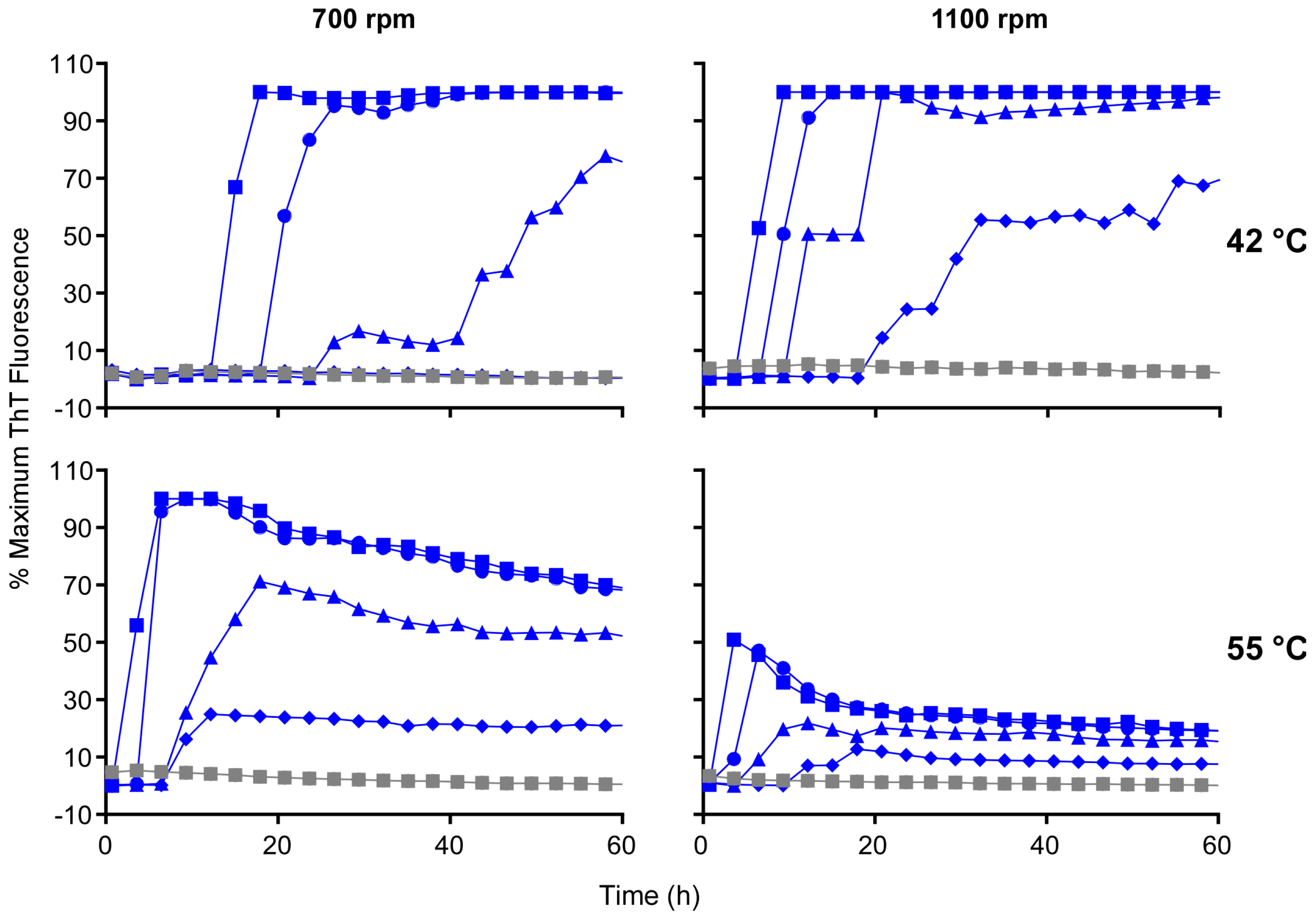
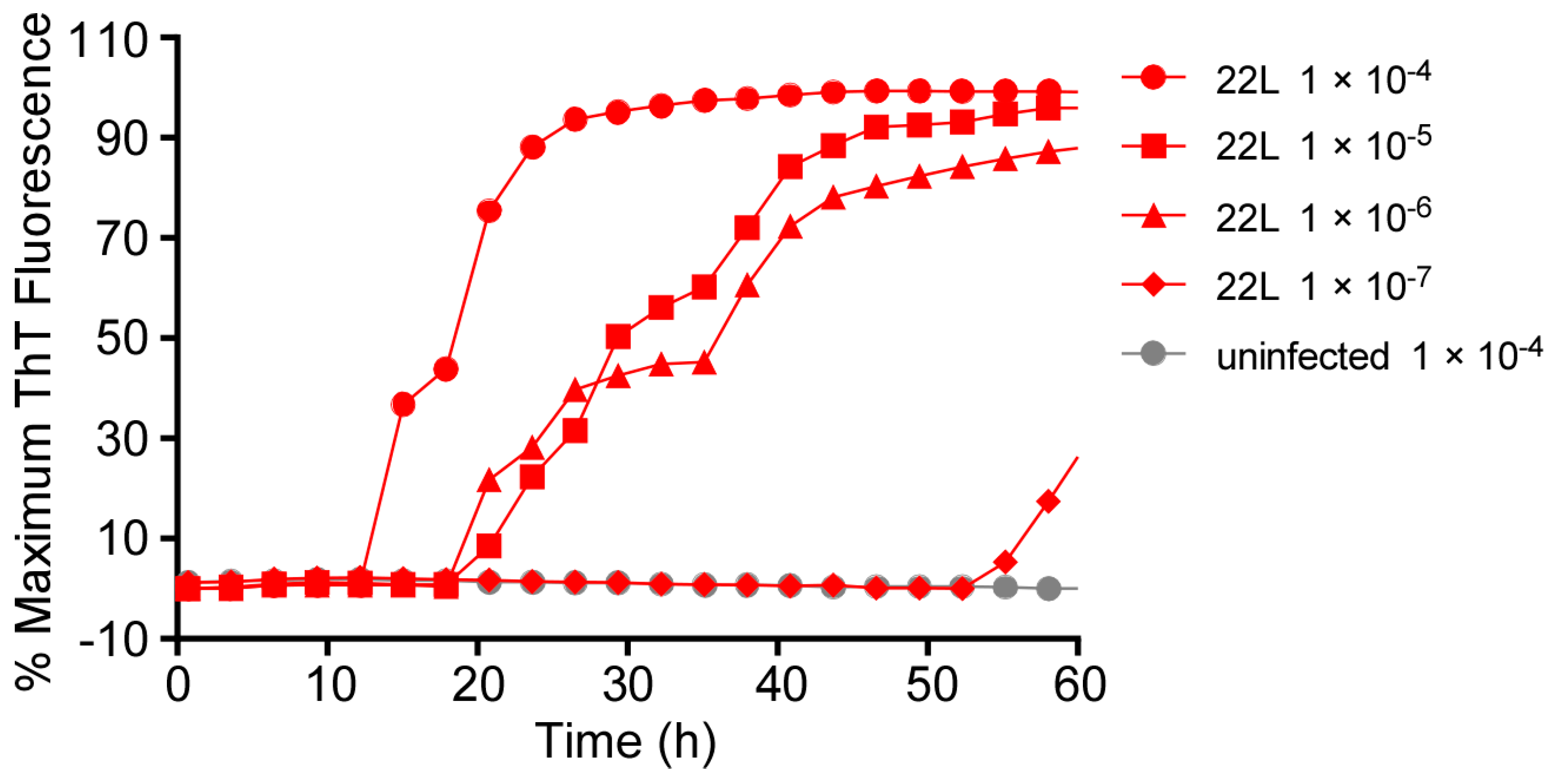
3.5. Acceleration of RT-QuIC with Higher Shaking Speeds
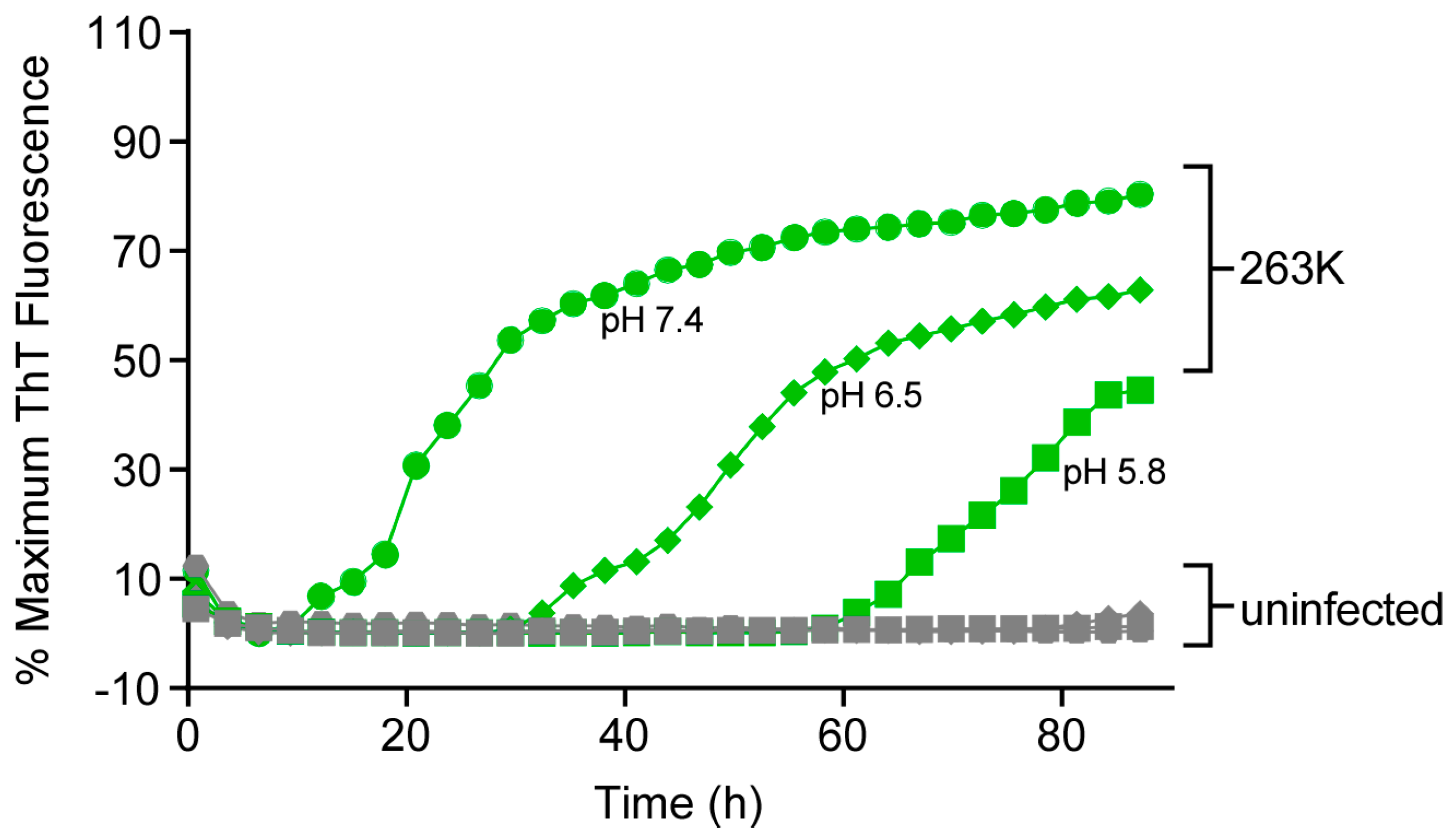
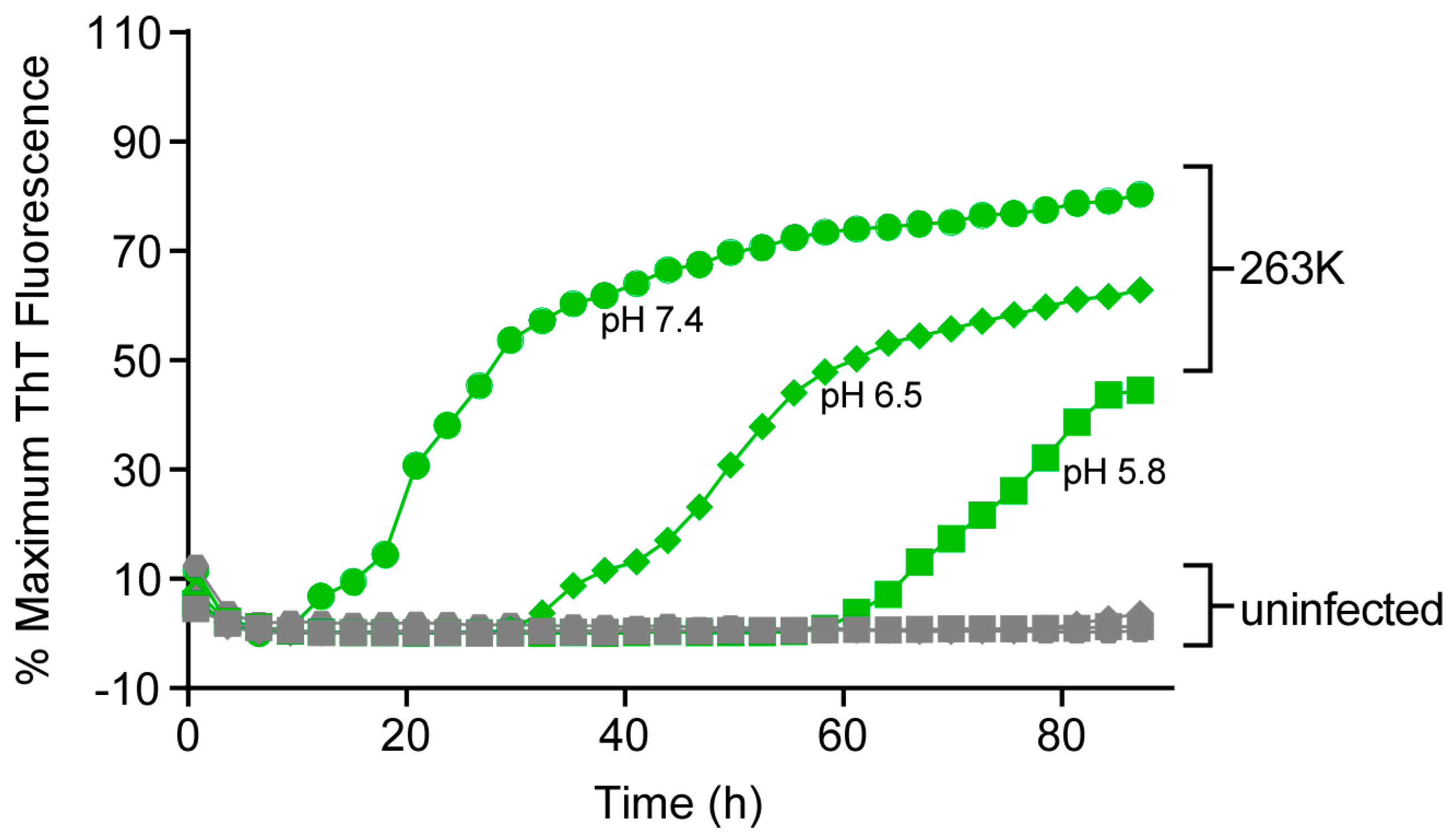
3.6. Effect of pH on RT-QuIC Amplification Kinetics
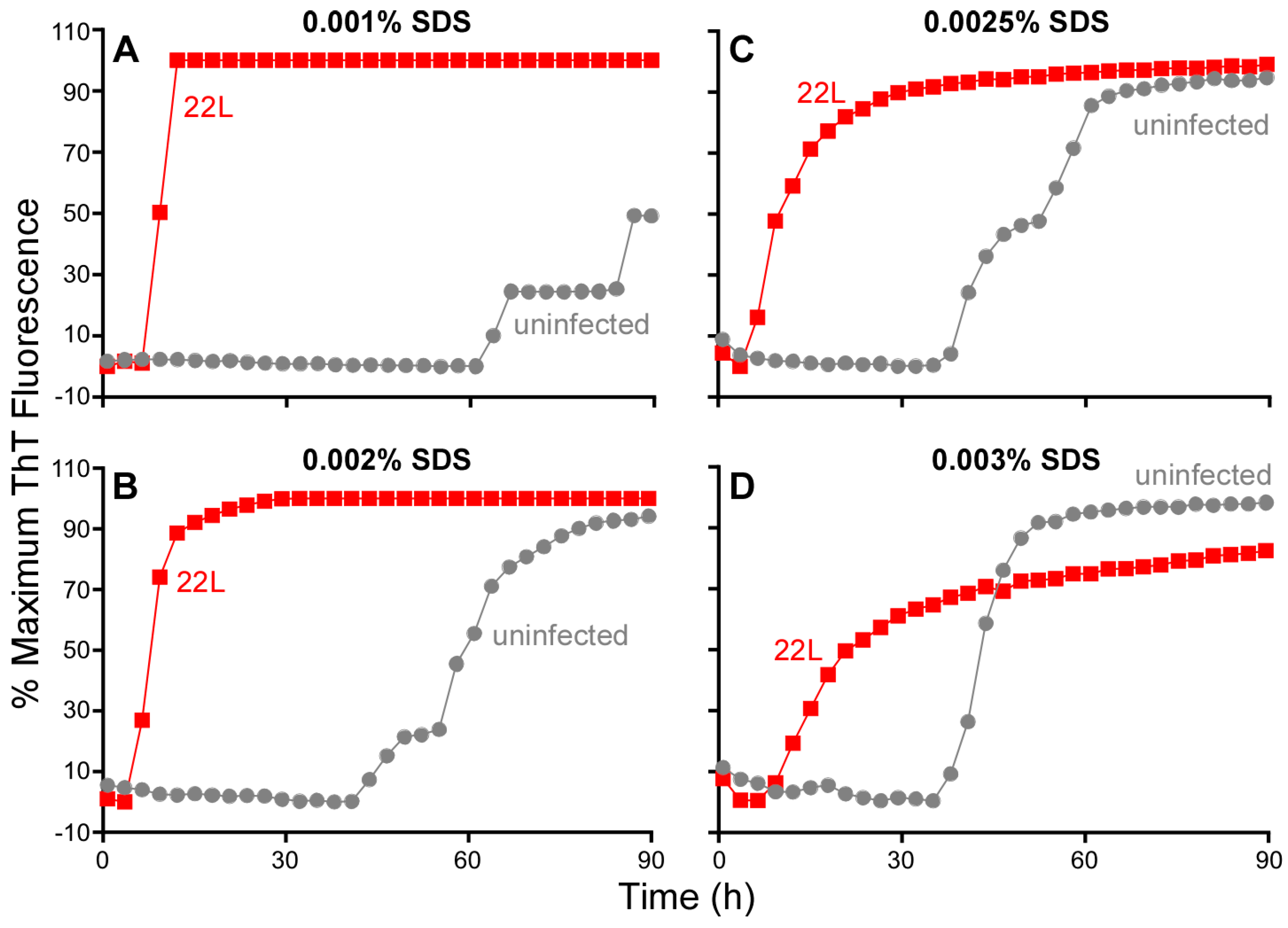
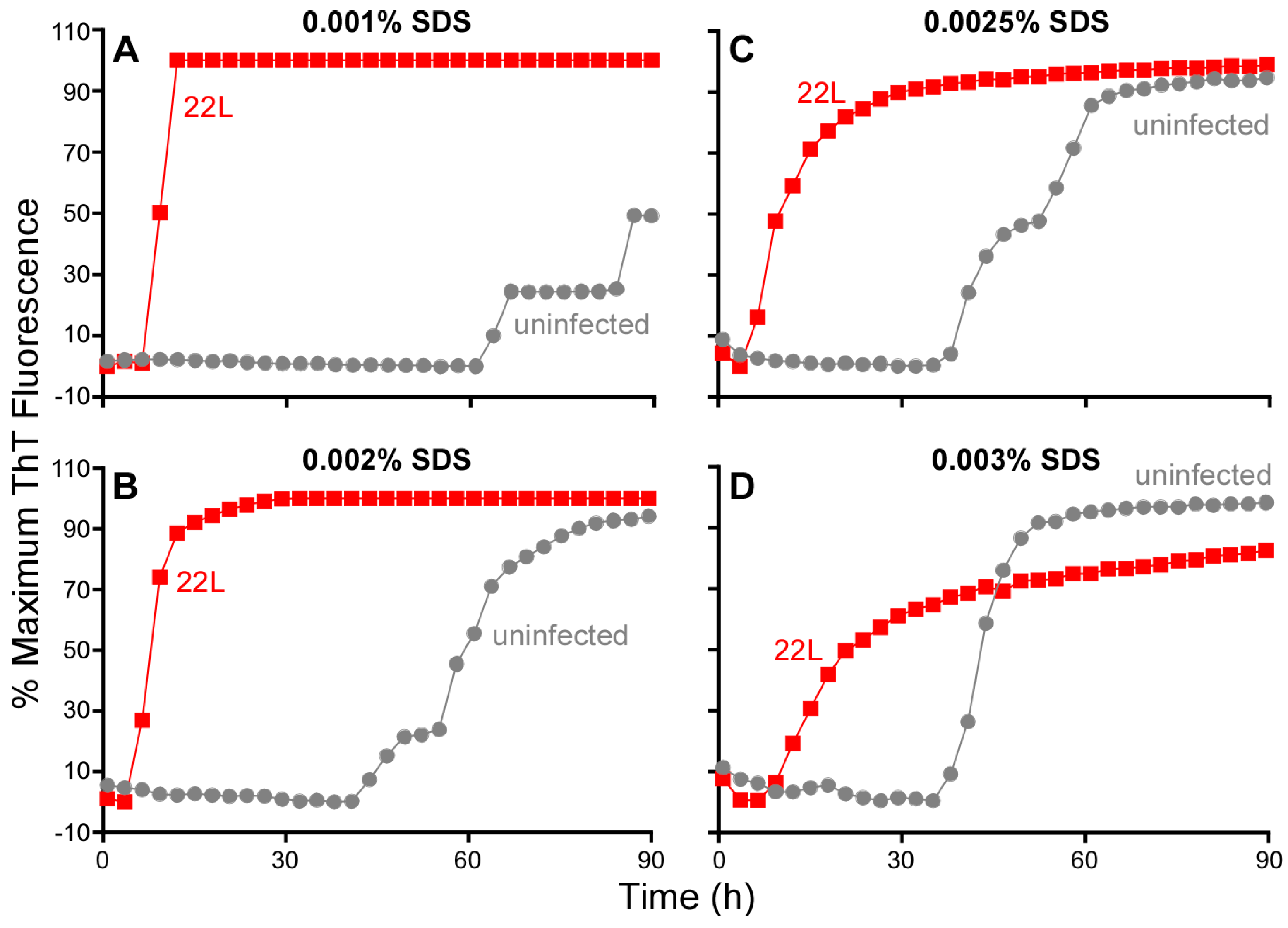
3.7. Influence of SDS on RT-QuIC Amplification Kinetics
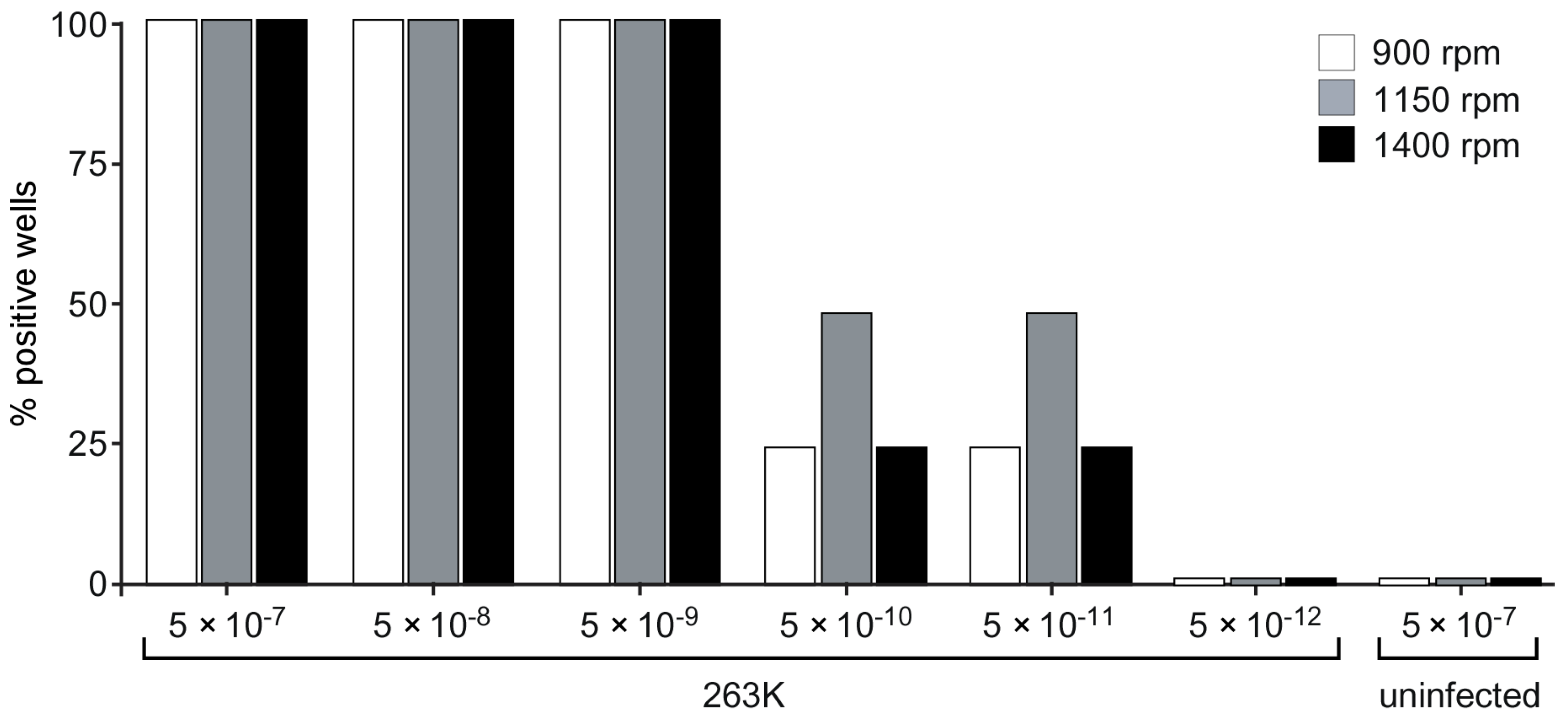
3.8. Prion Seed Detection Using a Separate Shaker-Incubator and Single-Timepoint Readings on a Fluorimeter
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kraus, A.; Groveman, B.R.; Caughey, B. Prions and the potential transmissibility of protein misfolding diseases. Annu. Rev. Microbiol. 2013, 67, 543–564. [Google Scholar] [CrossRef] [PubMed]
- Colby, D.W.; Zhang, Q.; Wang, S.; Groth, D.; Legname, G.; Riesner, D.; Prusiner, S.B. Prion detection by an amyloid seeding assay. Proc. Natl. Acad. Sci. USA 2007, 104, 20914–20919. [Google Scholar] [CrossRef] [PubMed]
- Wilham, J.M.; Orrú, C.D.; Bessen, R.A.; Atarashi, R.; Sano, K.; Race, B.; Meade-White, K.D.; Taubner, L.M.; Timmes, A.; Caughey, B. Rapid end-point quantitation of prion seeding activity with sensitivity comparable to bioassays. PLoS Pathog. 2010, 6, e1001217. [Google Scholar] [CrossRef] [PubMed]
- Atarashi, R.; Satoh, K.; Sano, K.; Fuse, T.; Yamaguchi, N.; Ishibashi, D.; Matsubara, T.; Nakagaki, T.; Yamanaka, H.; Shirabe, S.; et al. Ultrasensitive human prion detection in cerebrospinal fluid by real-time quaking-induced conversion. Nat. Med. 2011, 17, 175–178. [Google Scholar] [CrossRef] [PubMed]
- Orru, C.D.; Wilham, J.M.; Raymond, L.D.; Kuhn, F.; Schroeder, B.; Raeber, A.J.; Caughey, B. Prion disease blood test using immunoprecipitation and improved quaking-induced conversion. MBio 2011, 2, e00078-11. [Google Scholar] [CrossRef] [PubMed]
- Vascellari, S.; Orru, C.D.; Hughson, A.G.; King, D.; Barron, R.; Wilham, J.M.; Baron, G.S.; Race, B.; Pani, A.; Caughey, B. Prion seeding activities of mouse scrapie strains with divergent PrPSc protease sensitivities and amyloid plaque content using RT-QuIC and equic. PLoS ONE 2012, 7, e48969. [Google Scholar] [CrossRef] [PubMed]
- Elder, A.M.; Henderson, D.M.; Nalls, A.V.; Wilham, J.M.; Caughey, B.W.; Hoover, E.A.; Kincaid, A.E.; Bartz, J.C.; Mathiason, C.K. In vitro detection of prionemia in TSE-infected cervids and hamsters. PLoS ONE 2013, 8, e80203. [Google Scholar] [CrossRef] [PubMed]
- Henderson, D.M.; Manca, M.; Haley, N.J.; Denkers, N.D.; Nalls, A.V.; Mathiason, C.K.; Caughey, B.; Hoover, E.A. Rapid antemortem detection of CWD prions in deer saliva. PLoS ONE 2013, 8, e74377. [Google Scholar]
- Peden, A.H.; McGuire, L.I.; Appleford, N.E.; Mallinson, G.; Wilham, J.M.; Orru, C.D.; Caughey, B.; Ironside, J.W.; Knight, R.S.; Will, R.G.; et al. Sensitive and specific detection of sporadic Creutzfeldt-Jakob disease brain prion protein using real-time quaking induced conversion. J. Gen. Virol. 2012, 93, 438–449. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.; Mitteregger-Kretzschmar, G.; Giese, A.; Kretzschmar, H.A. Establishing quantitative real-time quaking-induced conversion (qRT-QuIC) for highly sensitive detection and quantification of PrPSc in prion-infected tissues. Acta Neuropathol Commun. 2013, 1. [Google Scholar] [CrossRef] [PubMed]
- Henderson, D.M.; Davenport, K.A.; Haley, N.J.; Denkers, N.D.; Mathiason, C.K.; Hoover, E.A. Quantitative assessment of prion infectivity in tissues and body fluids by real-time quaking-induced conversion. J. Gen. Virol. 2015, 96, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Orru, C.D.; Groveman, B.R.; Hughson, A.G.; Zanusso, G.; Coulthart, M.B.; Caughey, B. Rapid and sensitive Rt-QuIC detection of human Creutzfeldt-Jakob disease using cerebrospinal fluid. MBio 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- McGuire, L.I.; Peden, A.H.; Orru, C.D.; Wilham, J.M.; Appleford, N.E.; Mallinson, G.; Andrews, M.; Head, M.W.; Caughey, B.; Will, R.G.; et al. RT-QuIC analysis of cerebrospinal fluid in sporadic Creutzfeldt-Jakob disease. Ann. Neurol. 2012, 72, 278–285. [Google Scholar] [CrossRef] [PubMed]
- Cramm, M.; Schmitz, M.; Karch, A.; Mitrova, E.; Kuhn, F.; Schroeder, B.; Raeber, A.; Varges, D.; Kim, Y.S.; Satoh, K.; et al. Stability and reproducibility underscore utility of RT-QuIC for diagnosis of Creutzfeldt-Jakob disease. Mol. Neurobiol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Cramm, M.; Schmitz, M.; Karch, A.; Zafar, S.; Varges, D.; Mitrova, E.; Schroeder, B.; Raeber, A.; Kuhn, F.; Zerr, I. Characteristic CSF prion seeding efficiency in humans with prion diseases. Mol. Neurobiol. 2015, 51, 396–405. [Google Scholar] [CrossRef] [PubMed]
- Orru, C.D.; Bongianni, M.; Tonoli, G.; Ferrari, S.; Hughson, A.G.; Groveman, B.R.; Fiorini, M.; Pocchiari, M.; Monaco, S.; Caughey, B.; et al. A test for Creutzfeldt-Jakob disease using nasal brushings. N. Engl. J. Med. 2014, 371, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Zanusso, G.; Bongianni, M.; Caughey, B. A test for Creutzfeldt-Jakob disease using nasal brushings. N. Engl. J. Med. 2014, 371, 1842–1843. [Google Scholar] [PubMed]
- Orru, C.D.; Groveman, B.R.; Raymond, L.D.; Hughson, A.G.; Nonno, R.; Zou, W.; Ghetti, B.; Gambetti, P.; Caughey, B. Correction: Bank vole prion protein as an apparently universal substrate for RT-QuIC-based detection and discrimination of prion strains. PLoS Pathog. 2015, 11, e1005117. [Google Scholar] [CrossRef] [PubMed]
- Orru, C.D.; Favole, A.; Corona, C.; Mazza, M.; Manca, M.; Groveman, B.R.; Hughson, A.G.; Acutis, P.L.; Caramelli, M.; Zanusso, G.; et al. Detection and discrimination of classical and atypical L-type bovine spongiform encephalopathy by real-time quaking-induced conversion. J. Clin. Microbiol. 2015, 53, 1115–1120. [Google Scholar] [CrossRef] [PubMed]
- Haley, N.J.; Siepker, C.; Hoon-Hanks, L.L.; Mitchell, G.; Walter, W.D.; Manca, M.; Monello, R.J.; Powers, J.G.; Wild, M.A.; Hoover, E.A.; et al. Seeded amplification of chronic wasting disease prions in nasal brushings and recto-anal mucosa associated lymphoid tissues from elk by real time quaking-induced conversion. J. Clin. Microbiol. 2016, 54, 1117–1126. [Google Scholar] [CrossRef] [PubMed]
- Orru, C.D.; Groveman, B.R.; Raymond, L.D.; Hughson, A.G.; Nonno, R.; Zou, W.; Ghetti, B.; Gambetti, P.; Caughey, B. Bank Vole Prion Protein As an Apparently Universal Substrate for RT-QuIC-Based Detection and Discrimination of Prion Strains. PLoS Pathog. 2015, 11, e1004983. [Google Scholar]
- Masujin, K.; Orru, C.D.; Miyazawa, K.; Groveman, B.R.; Raymond, L.D.; Hughson, A.G.; Caughey, B. Detection of Atypical H-Type Bovine Spongiform Encephalopathy and Discrimination of Bovine Prion Strains by Real-Time Quaking-Induced Conversion. J. Clin. Microbiol. 2016, 54, 676–686. [Google Scholar] [CrossRef] [PubMed]
- Dassanayake, R.P.; Orru, C.D.; Hughson, A.G.; Caughey, B.; Graca, T.; Zhuang, D.; Madsen-Bouterse, S.A.; Knowles, D.P.; Schneider, D.A. Sensitive and specific detection of classical scrapie prions in the brains of goats by real-time quaking-induced conversion. J. Gen. Virol. 2016, 97, 803–812. [Google Scholar] [CrossRef] [PubMed]
- Cheng, K.; Sloan, A.; Avery, K.M.; Coulthart, M.; Carpenter, M.; Knox, J.D. Exploring physical and chemical factors influencing the properties of recombinant prion protein and the real-time quaking-induced conversion (RT-QuIC) assay. PLoS ONE 2014, 9, e84812. [Google Scholar] [CrossRef] [PubMed]
- Groveman, B.R.; Kraus, A.; Raymond, L.D.; Dolan, M.A.; Anson, K.J.; Dorward, D.W.; Caughey, B. Charge neutralization of the central lysine cluster in prion protein (PrP) promotes PrP(Sc)-like folding of recombinant PrP amyloids. J. Biol. Chem. 2015, 290, 1119–1128. [Google Scholar] [CrossRef] [PubMed]
- Kraus, A.; Anson, K.J.; Raymond, L.D.; Martens, C.; Groveman, B.R.; Dorward, D.W.; Caughey, B. Prion protein prolines 102 and 105 and the surrounding lysine cluster impede amyloid formation. J. Biol. Chem. 2015, 290, 21510–21522. [Google Scholar] [CrossRef] [PubMed]
- Groveman, B.R.; Dolan, M.A.; Taubner, L.M.; Kraus, A.; Wickner, R.B.; Caughey, B. Parallel in-register intermolecular beta-sheet architectures for prion-seeded prion protein (PrP) amyloids. J. Biol. Chem. 2014, 289, 24129–24142. [Google Scholar] [CrossRef] [PubMed]
- Tahiri-Alaoui, A.; Sim, V.L.; Caughey, B.; James, W. Molecular heterosis of prion protein beta-oligomers. A potential mechanism of human resistance to disease. J. Biol. Chem. 2006, 281, 34171–34178. [Google Scholar] [CrossRef] [PubMed]
- Murray, A.N.; Palhano, F.L.; Bieschke, J.; Kelly, J.W. Surface adsorption considerations when working with amyloid fibrils in multiwell plates and eppendorf tubes. Protein Sci. 2013, 22, 1531–1541. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.G.; Graham, C.; Dudas, S.; Paxman, E.; Vuong, B.; Czub, S. Defining and assessing analytical performance criteria for transmissible spongiform encephalopathy-detecting amyloid seeding assays. J. Mol. Diagn. 2016, 18, 454–467. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Accession Number | Amino Acid Residues | Genotype | SDS (%) | NaCl (mM) | Temperature (°C) | Prion Types Detected | References |
|---|---|---|---|---|---|---|---|---|
| Hamster | K02234 | 23–231 | NA | 0.002 | 300, 324 | 42 | 263K, sCJD, vCJD, L-BSE | [3,9,19] |
| 90–231 | NA | 0.002 | 300, 350 | 42, 55 | 263K, sCJD, CWD, HY-TME, L-BSE | [3,8,9,12,19,20] | ||
| Hamster-Sheep Chimera | Hamster K02234 | Hamster 23–137 | NA | 0.002 | 300 | 42 | vCJD, C-BSE, L-BSE | [5,19] |
| Sheep AY907689 | Sheep 141–234 | NA | ||||||
| Bank vole | AF367624 | 23–230 | M109 | 0.001 | 300 | 42 | sCJD, vCJD, iCJD, GSS, gCJD, FFI, sFI, 6-octarepeat insert, CS, AS, C-, L- and H-type BSE, CWD, 263K, HY- and DW-TME, Chandler, ME7, 87V, 22L WT and GPI− | [21,22] |
| 90–230 | M109 | 0.001 | 300 | 55 | H-BSE, L-BSE | [22] | ||
| 23–230 | I109 | 0.001 | 300 | 42 | H-BSE, L-BSE | [22] | ||
| Mouse | M13685 | 23–231 | NA | 0.001 | 130 | 42 | RML, RML GPI−, ME7, ME7 GPI−, 22L, 22L GPI−, C-BSE, H-BSE, L-BSE | ([6] Figure 9 and Figure 10 **), [22] |
| Sheep | AJ567988 | 25–234 | VRQ | 0.002 | 200, 300, 400 | 42 | Sheep scrapie (VRQ), goat scrapie | [3,23] |
| AY907689 | 25–234 | ARQ | 0.002 | 200 | 42 | Goat scrapie | [23] | |
| HM639748 | 25–234 | ARR | 0.001 | 300 | 42 | H-BSE, L-BSE | [22] | |
| Human | M13899 | 23–231 | M129 | 0.002 | 130 | 42 | sCJD, vCJD, H-BSE, L-BSE | [9,19,22] |
| 23–231 | V129 | 0.002 | 130 | 42 | sCJD, vCJD | [9] | ||
| Human-Bank vole Chimera | Human M13899 | Human 23–165 | M129 | 0.001 | 300 | 42 | H-BSE, L-BSE | [22] |
| Bank vole AF367624 | Bank vole 166–230 | NA | ||||||
| Deer | AF156185 | 24–234 | GMSSQ | 0.002 | 300 | 42 | CWD | [3] |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Orrú, C.D.; Hughson, A.G.; Groveman, B.R.; Campbell, K.J.; Anson, K.J.; Manca, M.; Kraus, A.; Caughey, B. Factors That Improve RT-QuIC Detection of Prion Seeding Activity. Viruses 2016, 8, 140. https://doi.org/10.3390/v8050140
Orrú CD, Hughson AG, Groveman BR, Campbell KJ, Anson KJ, Manca M, Kraus A, Caughey B. Factors That Improve RT-QuIC Detection of Prion Seeding Activity. Viruses. 2016; 8(5):140. https://doi.org/10.3390/v8050140
Chicago/Turabian StyleOrrú, Christina D., Andrew G. Hughson, Bradley R. Groveman, Katrina J. Campbell, Kelsie J. Anson, Matteo Manca, Allison Kraus, and Byron Caughey. 2016. "Factors That Improve RT-QuIC Detection of Prion Seeding Activity" Viruses 8, no. 5: 140. https://doi.org/10.3390/v8050140
APA StyleOrrú, C. D., Hughson, A. G., Groveman, B. R., Campbell, K. J., Anson, K. J., Manca, M., Kraus, A., & Caughey, B. (2016). Factors That Improve RT-QuIC Detection of Prion Seeding Activity. Viruses, 8(5), 140. https://doi.org/10.3390/v8050140