
Role of gga-miR-221 and gga-miR-222 during Tumour Formation in Chickens Infected by Subgroup J Avian Leukosis Virus


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Virus and Cell Lines
2.2. Cell Transfections
2.3. Cell Proliferation Assay
2.4. Colony Formation Assay
2.5. Wound Healing Assay
2.6. Analysis of Cell Apoptosis
2.7. Vector Construction
2.8. Luciferase Activity Assay
2.9. Western Blot
2.10. Animal Infection Assay
2.11. Real-Time Quantitative RT-PCR
2.12. Data Analysis
2.13. Ethics Statement
2.14. Accession Number
3. Results
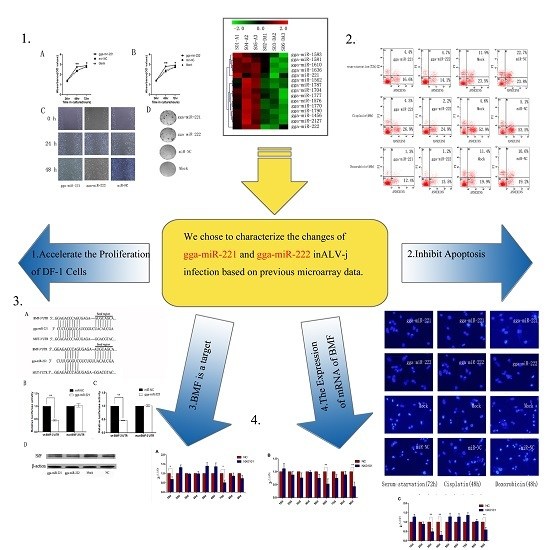
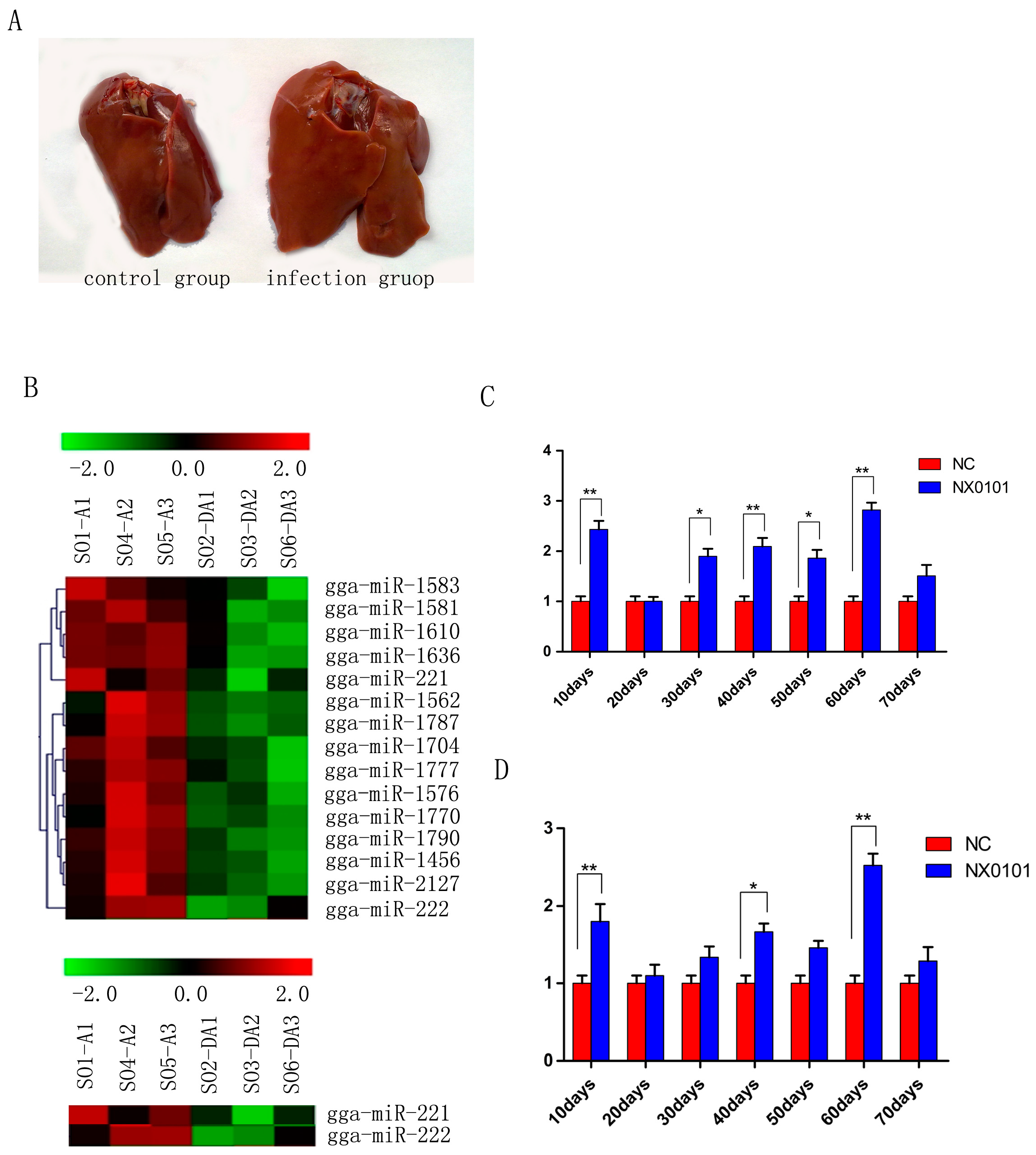
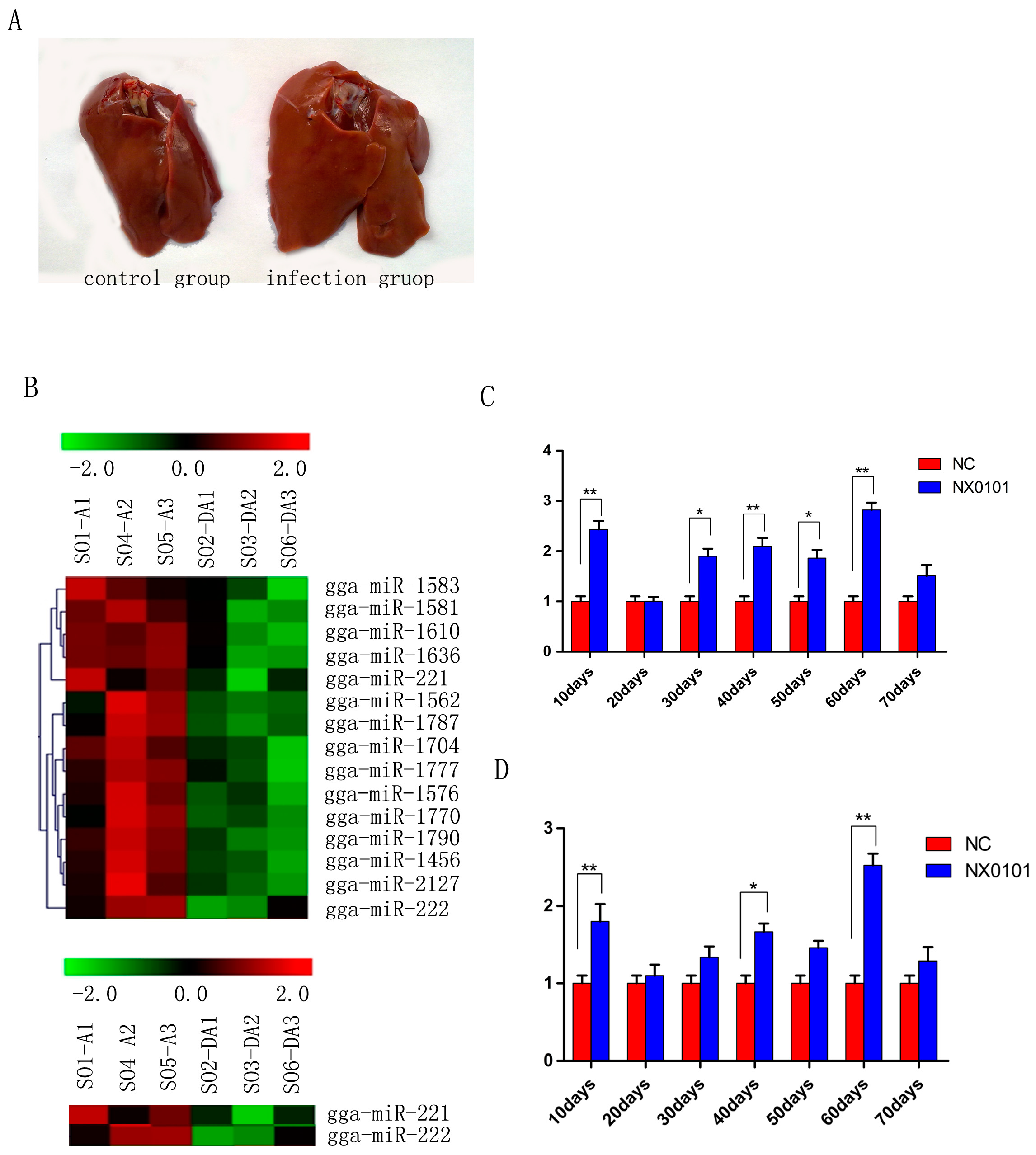
3.1. Expression of gga-miR-221 and gga-miR-222 in the Liver and Marrow of Pullets Infected with ALV-J
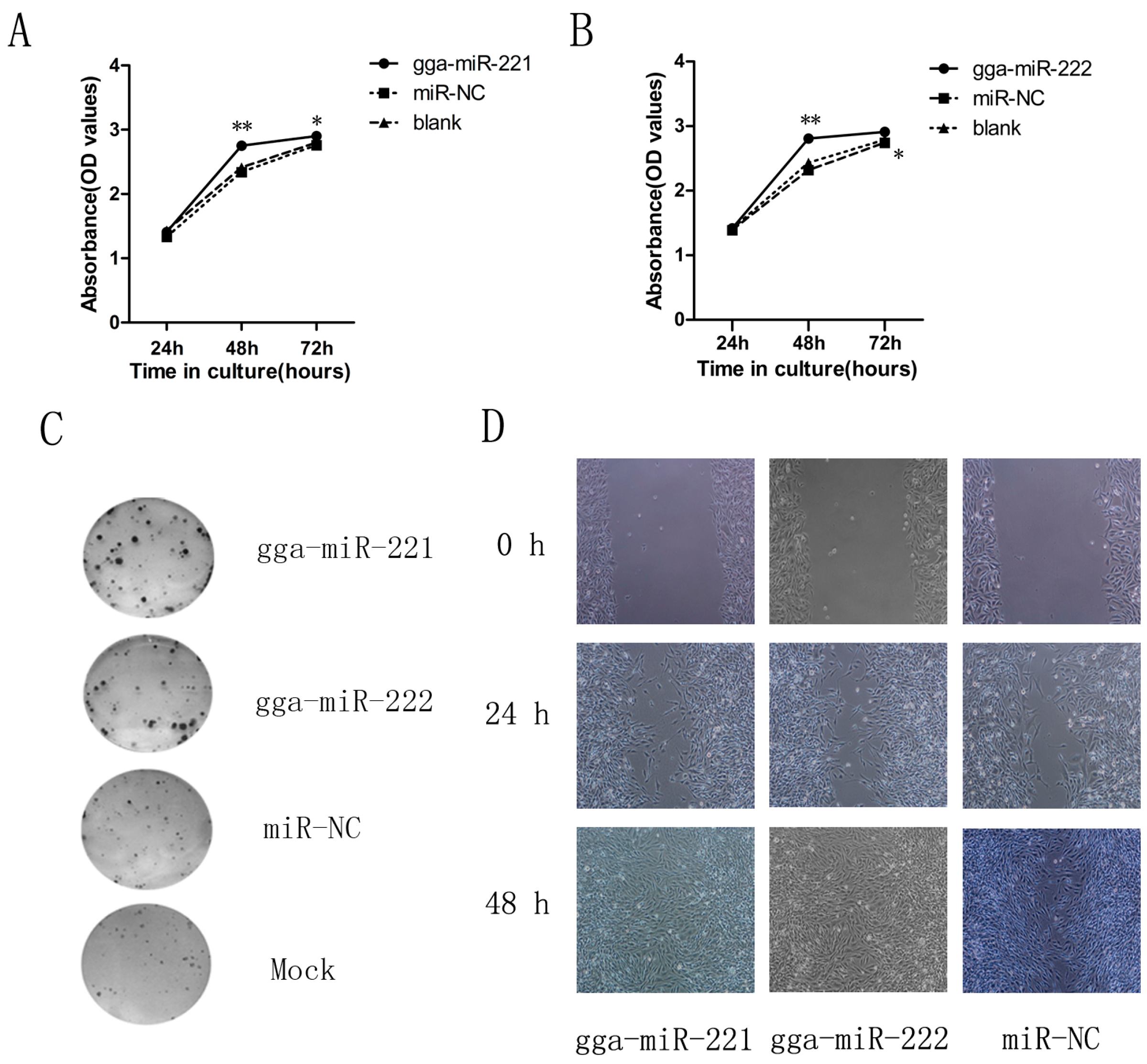
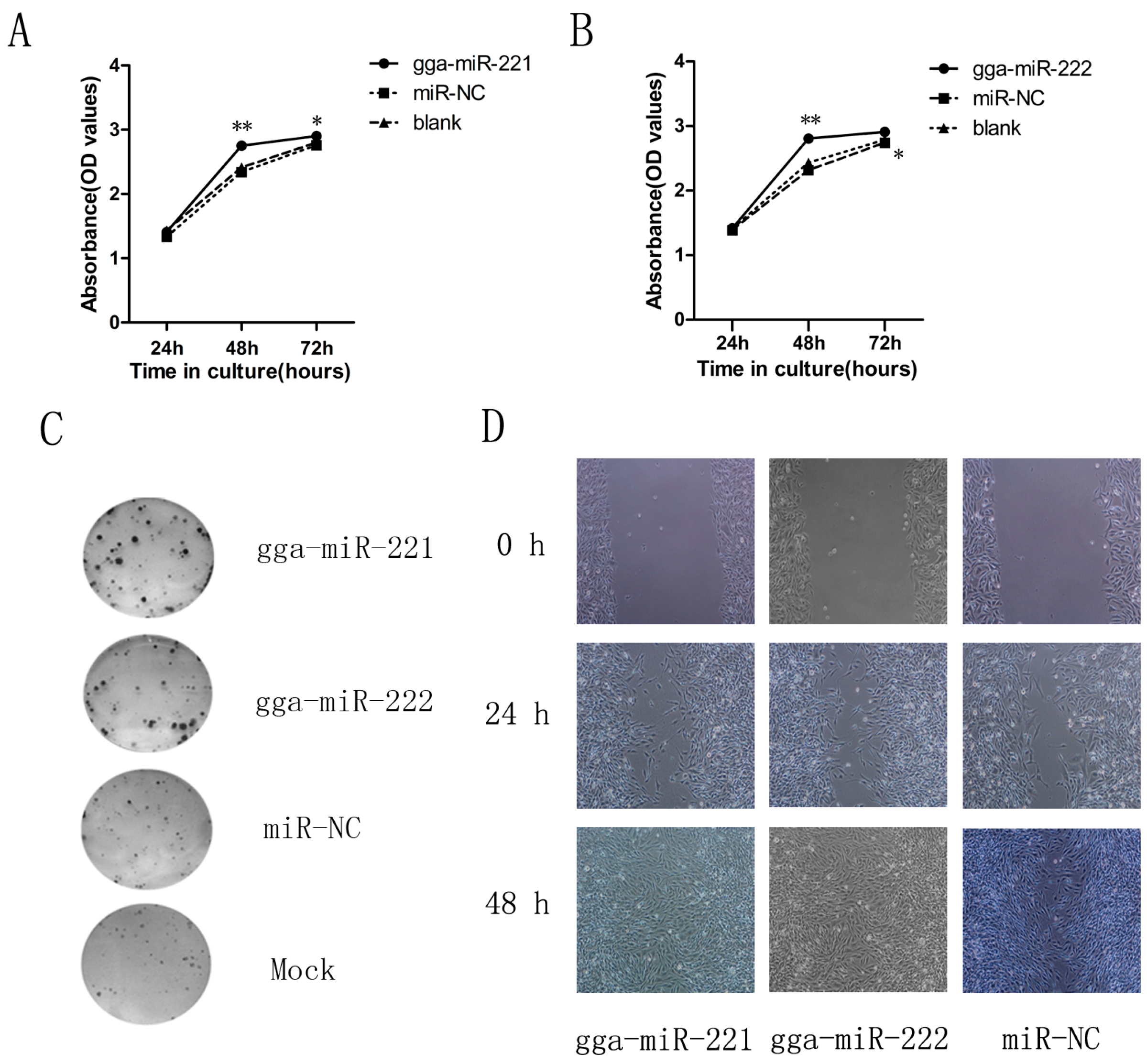
3.2. gga-miR-221 and gga-miR-222 Accelerate the Proliferation, Growth, and Migration of DF-1 Cells
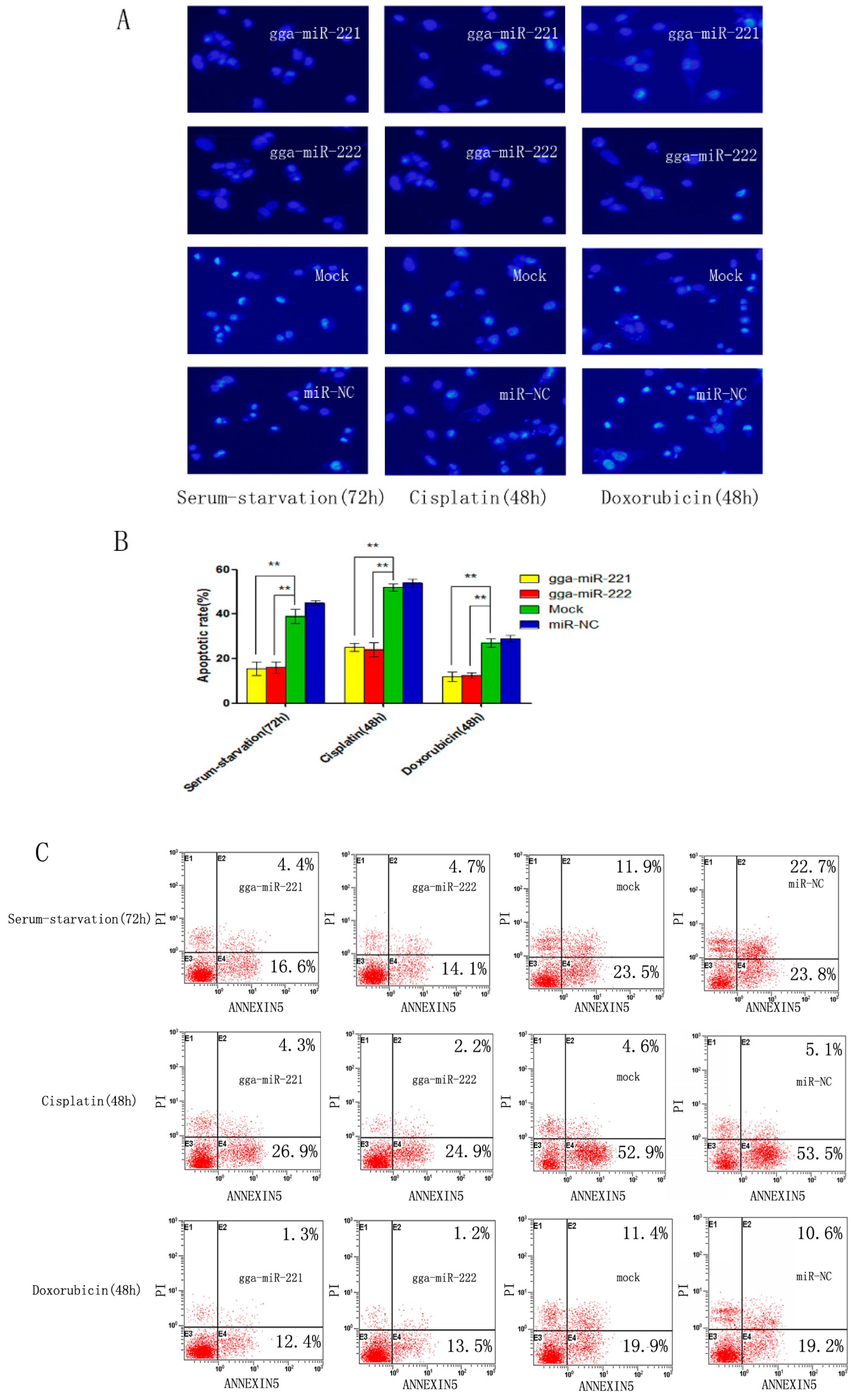
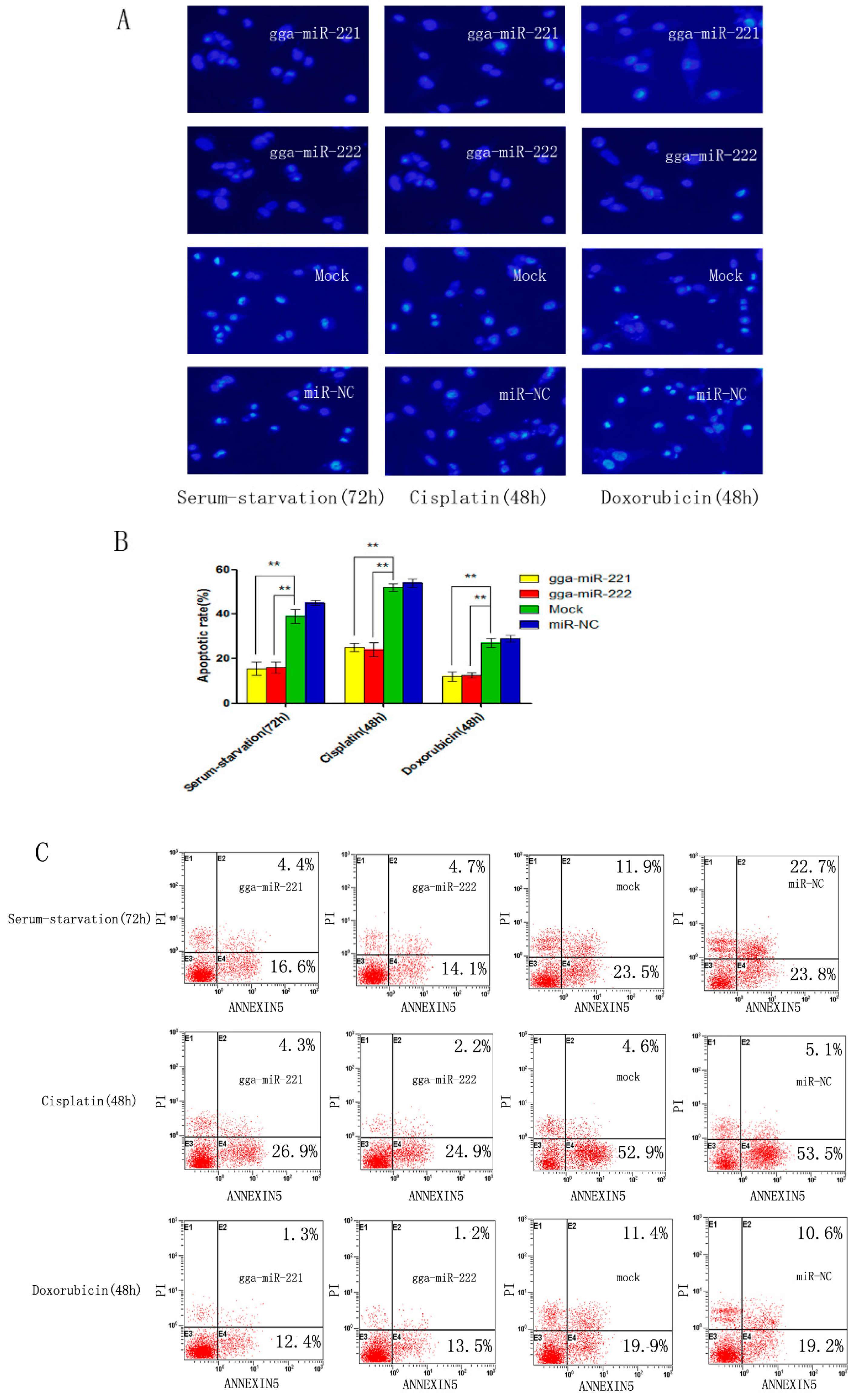
3.3. gga-miR-221 and gga-miR-222 Inhibit Apoptosis
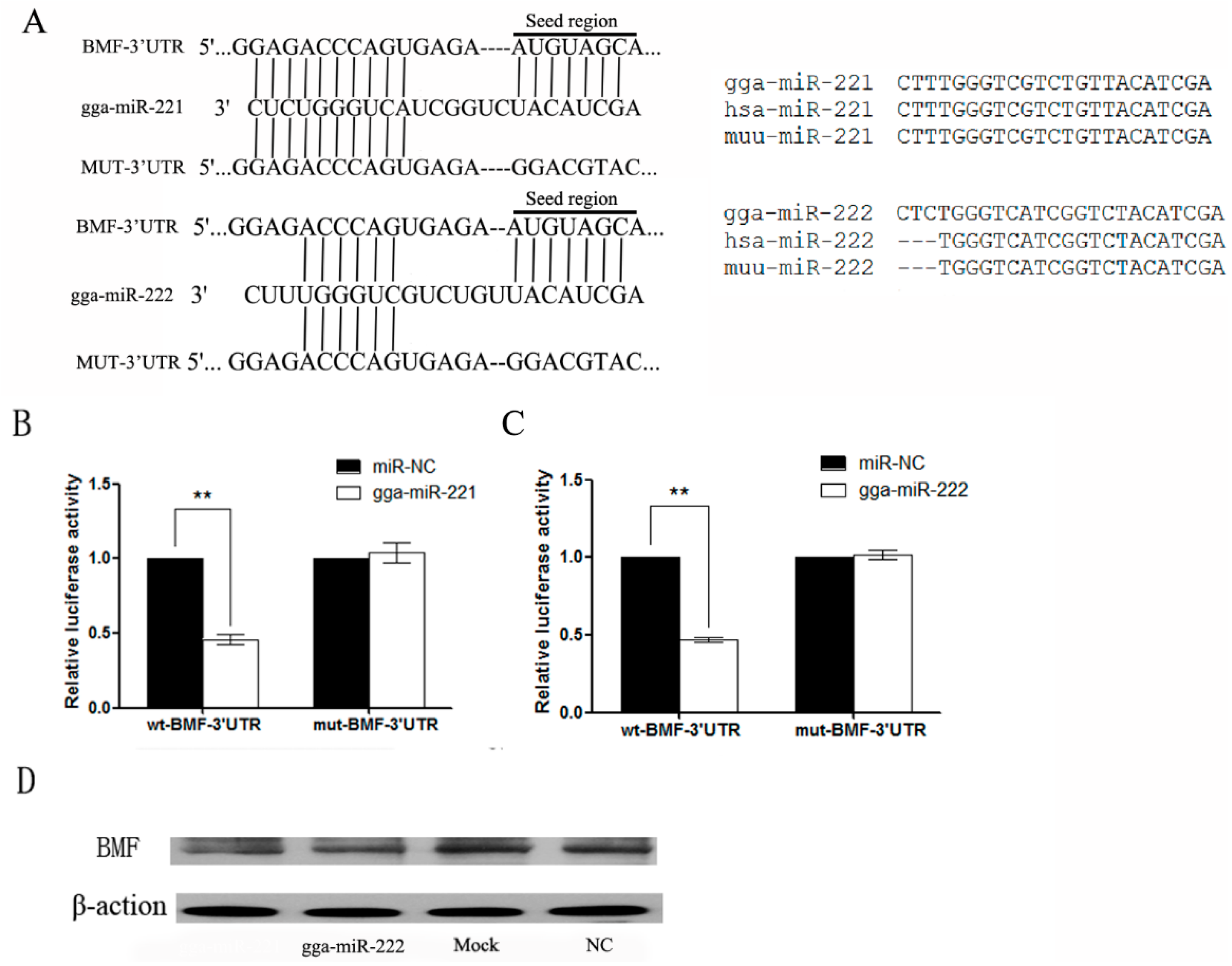
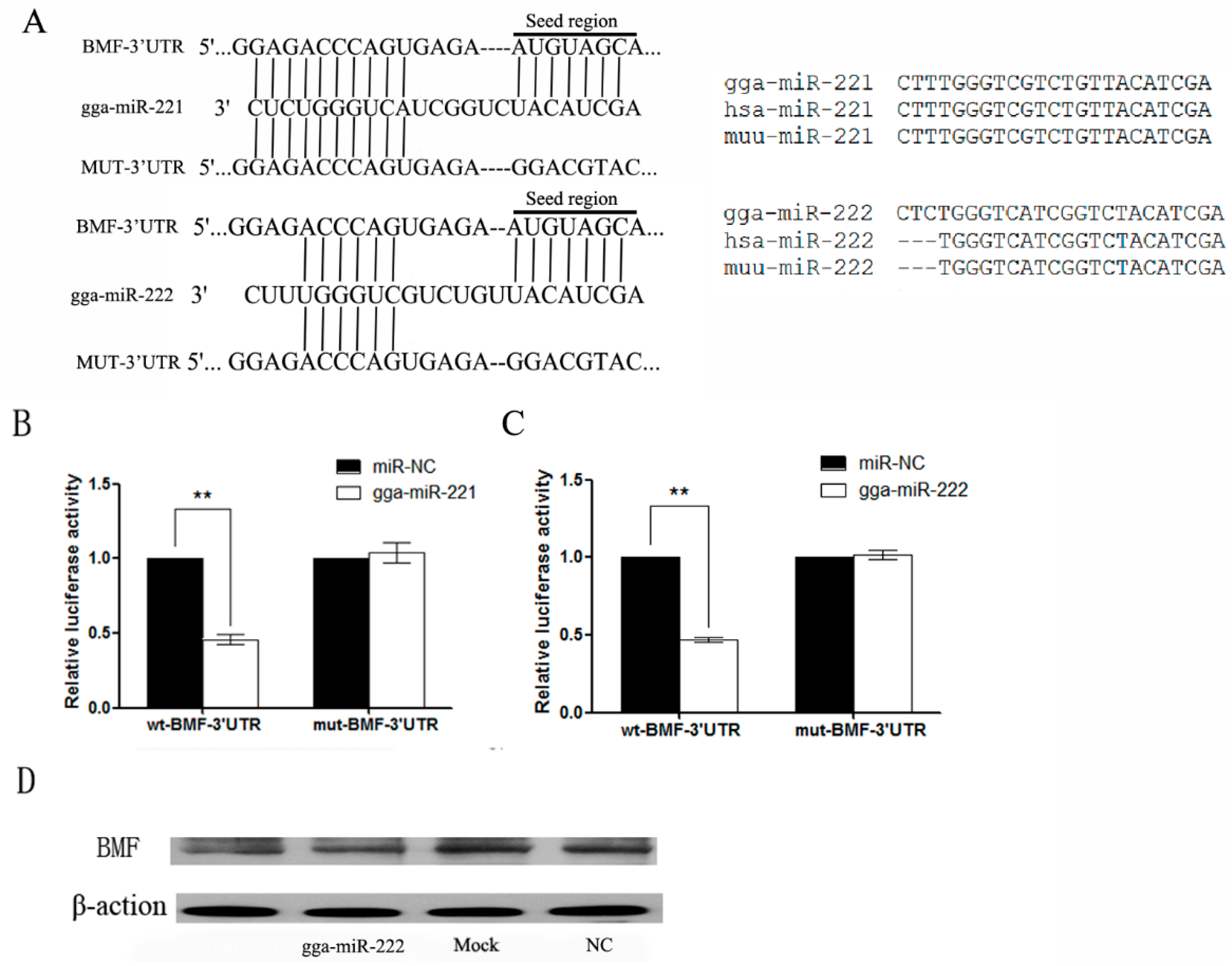
3.4. gga-miR-221 and gga-miR-222 Inhibit the Expression of BMF by Binding to the 3′-Untranslated Region (UTR)
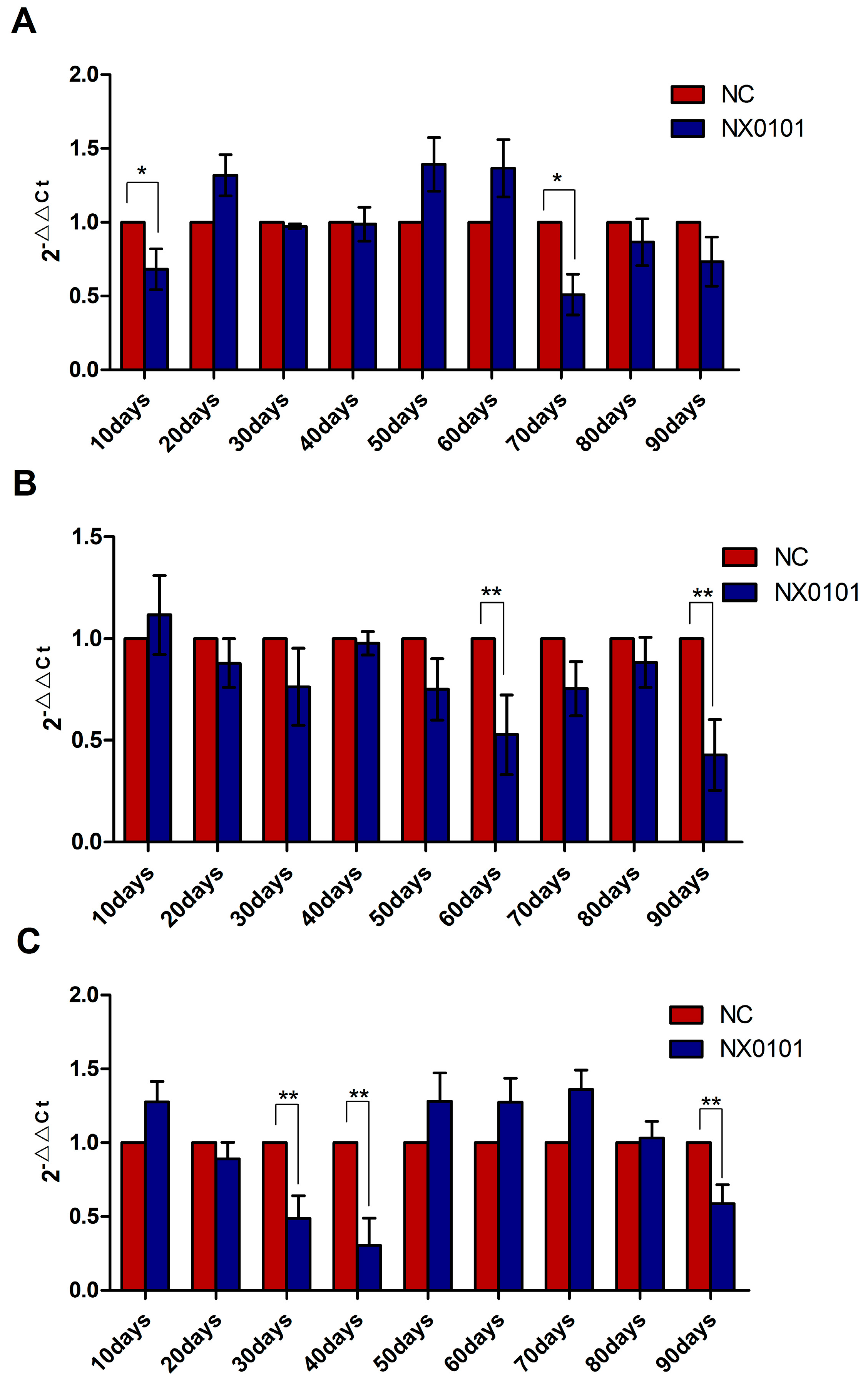
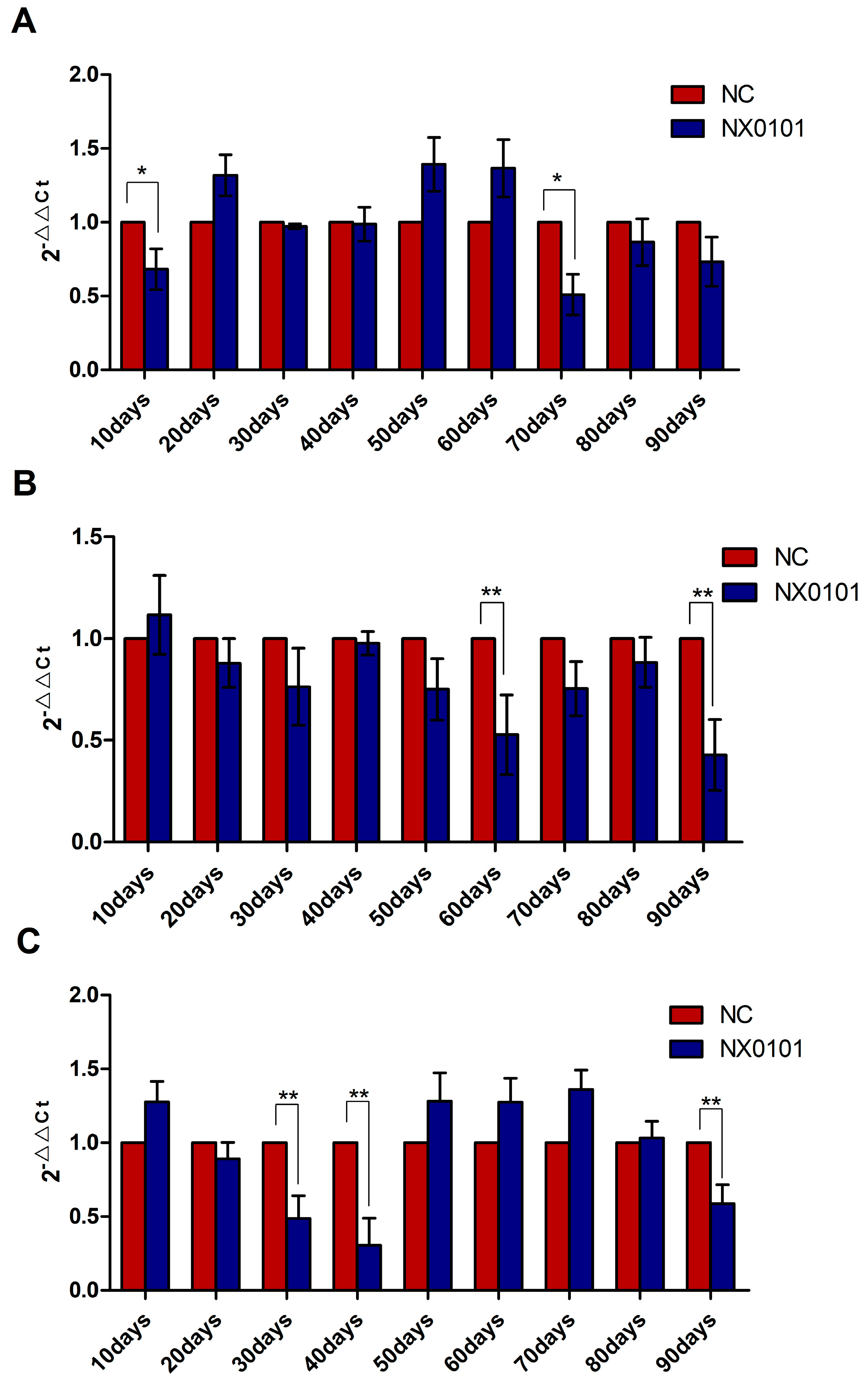
3.5. The Expression of mRNA of BMF on Liver, Blood, and Spleen
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Payne, L.N.; Nair, V. The long view: 40 Years of avian leukosis research. Avian Pathol. 2012, 41, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Ji, J.; Li, H.; Zhang, H.; Xie, Q.; Chang, S.; Shang, H.; Ma, J.; Bi, Y. Complete genome sequence of an avian leukosis virus isolate associated with hemangioma and myeloid leukosis in egg-type and meat-type chickens. J. Virol. 2012, 86, 10907–10908. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Xue, C.; Ji, J.; Chang, S.; Shang, H.; Zhang, L.; Ma, J.; Bi, Y.; Xie, Q. Complete genome sequence of a J subgroup avian leukosis virus isolated from local commercial broilers. J. Virol. 2012, 86, 11937–11938. [Google Scholar] [CrossRef] [PubMed]
- Neumann, U. Morphological evidence for avian leukosis virus (ALV) subgroup j avian myelocytomatosis observed in broiler parent flocks in Costa Rica. In Proceedings of the International Symposium on ALV-J and other Avian Retroviruses, Rauischolzhauzen, Germany, 5–8 June 2000; pp. 181–183.
- Bagust, T.J. Avian leukosis virus-J in Australia: Scenarios and options for control. In Proceedings of the International Symposium on ALV-J and Other Avian Retroviruses, Rauischolzhauzen, Germany, 5–8 June 2000; pp. 234–239.
- Bagust, T.J.; Fenton, S.P.; Reddy, M.R. Detection of subgroup j avian leukosis virus infection in Australian meat-type chickens. Aust. Vet. J. 2004, 82, 701–706. [Google Scholar] [CrossRef] [PubMed]
- Otsuka, K.; Ochiya, T. Genetic networks lead and follow tumor development: MicroRNA regulation of cell cycle and apoptosis in the p53 pathways. Biomed. Res. Int. 2014, 2014, 749724. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, D.W.; Thornberry, N.A. Apoptosis:Life and death decisions. Science 2014, 299, 214–215. [Google Scholar] [CrossRef] [PubMed]
- Antonsson, B. Bax and other pro-apoptotic Bcl-2 family “Killer-Proteins” and their victim the mitochondrion. Cell Tissue Res. 2001, 306, 347–361. [Google Scholar] [CrossRef] [PubMed]
- Cepero, E.; Johnson, B.W.; Boise, L.H. Cloning and analysis of Bcl-2 family genes. Methods Cell Biol. 2001, 66, 29–47. [Google Scholar] [PubMed]
- Weerasinghe, P.; Hallock, S.; Tang, S.; Liepins, A. Role of Bcl-2 family proteins and Caspase-3 in sanguinarine-induced bimodal cell death. Cell Biol. Toxicol. 2001, 17, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Zhang, J.; Zhang, A.; Wang, Y.; Han, L.; You, Y.; Pu, P.; Kang, C. PUMA is a novel target of miR-221/222 in human epithelial cancers. Int. J. Oncol. 2010, 37, 1621–1626. [Google Scholar] [PubMed]
- Marzo, I.; Brenner, C.; Zamzami, N.; Jürgensmeier, J.M.; Susin, S.A.; Vieira, H.L.; Prévost, M.C.; Xie, Z.; Matsuyama, S.; Reed, J.C.; et al. Bax and adenine nucleotide translocator cooperate in the mitochondrial control of apoptosis. Science 1998, 281, 2027–2031. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Tu, H.C.; Ren, D.; Takeuchi, O.; Jeffers, J.R.; Zambetti, G.P.; Hsieh, J.J.; Cheng, E.H. Stepwise Activation of BAX and BAK by tBID, BIM, and PUMA Initiates Mitochondrial Apoptosis. Mol. Cell 2009, 36, 487–499. [Google Scholar] [CrossRef] [PubMed]
- Ren, D.; Tu, H.C.; Kim, H.; Wang, G.X.; Bean, G.R.; Takeuchi, O.; Jeffers, J.R.; Zambetti, G.P.; Hsieh, J.J.; Cheng, E.H. BID, BIM, and PUMA are essential for activation of the BAX and BAK-dependent cell death program. Science 2010, 330, 1390–1393. [Google Scholar] [CrossRef] [PubMed]
- Letai, A. Puma Strikes Bax. J. Cell Biol. 2009, 185, 189–191. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Willis, S.N.; Wei, A.; Smith, B.J.; Fletcher, J.I.; Hinds, M.G.; Colman, P.M.; Day, C.L.; Adams, J.M.; Huang, D.C. Differential targeting of prosurvival Bcl-2 proteins by their BH3-only ligands allows complementary apoptotic function. Mol. Cell 2005, 17, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Gérecová, G.; Kopanicová, J.; Jaká, P.; Běhalová, L.; Juhásová, B.; Bhatia-Kiššová, I.; Forte, M.; Polčic, P.; Mentel, M. BH3-only proteins Noxa, Bik, Bmf, and Bid activate Bax and Bak indirectly when studied in yeast model. FEMS Yeast Res. 2013, 13, 747–754. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Esquer, F.; Palomar, M.A.; Rivas, I.; Delcan, J.; Linares, R.; Diaz-Gil, G. Characterization of the BH3 protein Bmf in Gallus gallus: Identification of a novel chicken-specific isoform. Gene 2008, 407, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Egle, A.; Harris, A.W.; Bouillet, P.; Cory, S. Bim is a suppressor of Myc-induced mouse B cell leukemia. Proc. Natl. Acad. Sci. USA 2004, 101, 6164–6169. [Google Scholar] [CrossRef] [PubMed]
- Tagawa, H.; Karnan, S.; Suzuki, R.; Matsuo, K.; Zhang, X.; Ota, A.; Morishima, Y.; Nakamura, S.; Seto, M. Genome-wide array-based CGH for mantle cell lymphoma: Identification of homozygous deletions of the proapoptotic gene BIM. Oncogene 2005, 24, 1348–1358. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Shukla, G.C.; Singh, J.; Barik, S. MicroRNAs: Processing, maturation, target recognition and regulatory functions. Mol. Cell Pharmacol. 2011, 3, 83–92. [Google Scholar] [PubMed]
- Calin, G.A.; Dumitru, C.D.; Shimizu, M.; Bichi, R.; Zupo, S.; Noch, E.; Aldler, H.; Rattan, S.; Keating, M.; Rai, K.; et al. Frequent deletions and down-regulation of micro-RNA genes miR15 and miR16 at 13Q14 in chronic lymphocytic leukemia. Proc. Natl. Acad. Sci. USA 2002, 99, 15524–15529. [Google Scholar] [CrossRef] [PubMed]
- Cimmino, A.; Calin, G.A.; Fabbri, M.; Iorio, M.V.; Ferracin, M.; Shimizu, M.; Wojcik, S.E.; Aqeilan, R.I.; Zupo, S.; Dono, M.; et al. MiR-15 and miR-16 induce apoptosis by targeting BCL2. Proc. Natl. Acad. Sci. USA 2005, 102, 13944–13949. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.F.; Dewson, G.; Evangelista, M.; Pettikiriarachchi, A.; Gold, G.J.; Zhu, H.; Colman, P.M.; Fairlie, W.D. The functional differences between pro-survival and pro-apoptotic B cell lymphoma 2 (Bcl-2) proteins depend on structural differences in their Bcl-2 Homology 3 (BH3) domains. J. Biol. Chem. 2014, 289, 36001–36017. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Shang, H.; Shu, D.; Zhang, H.; Ji, J.; Sun, B.; Xie, Q. Gga-miR-375 plays a key role in tumorigenesis post subgroup J avian leukosis virus infection. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Garofalo, M.; Quintavalle, C.; Romano, G.; Croce, C.M.; Condorelli, G. MiR221/222 in cancer: Their role in tumor progression and response to therapy. Curr. Mol. Med. 2012, 12, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Ji, J.; Xie, Q.; Shang, H.; Zhang, H.; Xin, X.; Chen, F.; Sun, B.; Xue, C.; Ma, J. Aberrant expression of liver micoRNA in chickens infected with subgroup J avian leukosis virus. Virus Res. 2012, 169, 268–271. [Google Scholar] [CrossRef] [PubMed]
- Frenquelli, M.; Muzio, M.; Scielzo, C.; Fazi, C.; Scarfò, L.; Rossi, C.; Ferrari, G.; Ghia, P.; Caligaris-Cappio, F. MicroRNA and proliferation control in chronic lymphocytic leukemia: Functional relationship between miR-221/222 cluster and P27. Blood 2010, 115, 3949–3959. [Google Scholar] [CrossRef] [PubMed]
- Rommer, A.; Steinleitner, K.; Hackl, H.; Schneckenleithner, C.; Engelmann, M.; Scheideler, M.; Vlatkovic, I.; Kralovics, R.; Cerny-Reiterer, S.; Valent, P.; et al. Overexpression of primary microRNA 221/222 in acute myeloid leukemia. BMC Cancer 2013, 13, 364. [Google Scholar] [CrossRef] [PubMed]
- Miller, T.E.; Ghoshal, K.; Ramaswamy, B.; Roy, S.; Datta, J.; Shapiro, C.L.; Vlatkovic, I.; Kralovics, R.; Cerny-Reiterer, S.; Valent, P.; et al. MicroRNA-221/222 confers tamoxifen resistance in breast cancer by targeting p27Kip1. J. Biol. Chem. 2008, 283, 29897–29903. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.; Cui, F.; Bu, Q.; Yan, C. The expression and clinical significance of GTP-binding RAS-like 3 (ARHI) and microRNA 221 and 222 in prostate cancer. J. Int. Med. Res. 2011, 39, 1870–1875. [Google Scholar] [CrossRef] [PubMed]
- Chun-Zhi, Z.; Lei, H.; An-Ling, Z.; Yan-Chao, F.; Xiao, Y.; Guang-Xiu, W.; Zhi-Fan, J.; Pei-Yu, P.; Qing-Yu, Z.; Chun-Sheng, K. MicroRNA-221 and microRNA-222 regulate gastric carcinoma cell proliferation and radioresistance by targeting PTEN. BMC Cancer 2010, 10. [Google Scholar] [CrossRef]
- Galardi, S.; Mercatelli, N.; Giorda, E.; Massalini, S.; Frajese, G.V.; Ciafrè, S.A.; Farace, M.G. MiR-221 and miR-222 expression affects the proliferation potential of human prostate carcinoma cell lines by targeting p27Kip1. J. Biol. Chem. 2007, 282, 23716–23724. [Google Scholar] [CrossRef] [PubMed]
- Felli, N.; Fontana, L.; Pelosi, E.; Botta, R.; Bonci, D.; Facchiano, F.; Liuzzi, F.; Lulli, V.; Morsilli, O.; Peschle, C.; et al. MicroRNAs 221 and 222 inhibit normal erythropoiesis and erythroleukemic cell growth via kit receptor down-modulation. Proc. Natl. Acad. Sci. USA 2005, 13, 18081–18086. [Google Scholar] [CrossRef] [PubMed]
- Gramantieri, L.; Fornari, F.; Ferracin, M.; Veronese, A.; Sabbioni, S.; Calin, G.A.; Grazi, G.L.; Croce, C.M.; Bolondi, L.; Negrini, M. MicroRNA-221 targets Bmf in hepatocellular carcinoma and correlates with tumor multifocality. Clin. Cancer Res. 2009, 15, 5073–5081. [Google Scholar] [CrossRef] [PubMed]
- Lambeth, L.S.; Yao, Y.; Smith, L.P.; Zhao, Y.; Nair, V. MicroRNAs 221 and 222 Target p27Kip1 in Marek’s disease virus-transformed tumour cell line MSB-1. J. Gen. Virol. 2009, 90, 1164–1171. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.; Sun, S.; Wang, J. Reduced serologic response to newcastle disease virus in broiler chickens exposed to a Chinese field strain of subgroup J avian leukosis virus. Avian Dis. 2006, 50, 191–195. [Google Scholar] [CrossRef] [PubMed]
- Kepp, O.; Gottschalk, K.; Churin, Y.; Rajalingam, K.; Brinkmann, V.; Machuy, N.; Kroemer, G.; Rudel, T. Bim and Bmf synergize to induce apoptosis in neisseria gonorrhoeae infection. PLoS Pathog. 2009, 3, e1000348. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Sun, A.J.; Teng, M.; Zhou, H.; Cui, Z.Z.; Qu, L.H.; Zhang, G.P. Expression profiles of microRNAs encoded by the oncogenic Marek’s Disease virus reveal two distinct expression patterns in vivo during different phases of disease. J. Gen. Virol. 2011, 92, 608–620. [Google Scholar] [CrossRef] [PubMed]
- Tian, F.; Luo, J.; Zhang, H.; Chang, S.; Song, J. MiRNA expression signatures induced by Marek’s disease virus infection in chickens. Genomics 2012, 99, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Dabrowska, M.J.; Dybkaer, K.; Johnsen, H.E. Loss of MicroRNA targets in the 3’ untranslated region as a mechanism of retroviral insertional activation of growth factor independence 1. J. Virol. 2009, 83, 8051–8061. [Google Scholar] [CrossRef] [PubMed]
- Chesters, P.M.; Howes, K.; McKay, J.C.; Payne, L.N.; Venugopal, K. Acutely transforming avian leukosis virus subgroup J strain 966: Defective genome encodes a 72-kilodalton Gag-Myc fusion protein. J. Virol. 2001, 75, 4219–4225. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dai, Z.; Ji, J.; Yan, Y.; Lin, W.; Li, H.; Chen, F.; Liu, Y.; Chen, W.; Bi, Y.; Xie, Q. Role of gga-miR-221 and gga-miR-222 during Tumour Formation in Chickens Infected by Subgroup J Avian Leukosis Virus. Viruses 2015, 7, 6538-6551. https://doi.org/10.3390/v7122956
Dai Z, Ji J, Yan Y, Lin W, Li H, Chen F, Liu Y, Chen W, Bi Y, Xie Q. Role of gga-miR-221 and gga-miR-222 during Tumour Formation in Chickens Infected by Subgroup J Avian Leukosis Virus. Viruses. 2015; 7(12):6538-6551. https://doi.org/10.3390/v7122956
Chicago/Turabian StyleDai, Zhenkai, Jun Ji, Yiming Yan, Wencheng Lin, Hongxin Li, Feng Chen, Yang Liu, Weiguo Chen, Yingzuo Bi, and Qingmei Xie. 2015. "Role of gga-miR-221 and gga-miR-222 during Tumour Formation in Chickens Infected by Subgroup J Avian Leukosis Virus" Viruses 7, no. 12: 6538-6551. https://doi.org/10.3390/v7122956
APA StyleDai, Z., Ji, J., Yan, Y., Lin, W., Li, H., Chen, F., Liu, Y., Chen, W., Bi, Y., & Xie, Q. (2015). Role of gga-miR-221 and gga-miR-222 during Tumour Formation in Chickens Infected by Subgroup J Avian Leukosis Virus. Viruses, 7(12), 6538-6551. https://doi.org/10.3390/v7122956