
Expression, Delivery and Function of Insecticidal Proteins Expressed by Recombinant Baculoviruses


Abstract
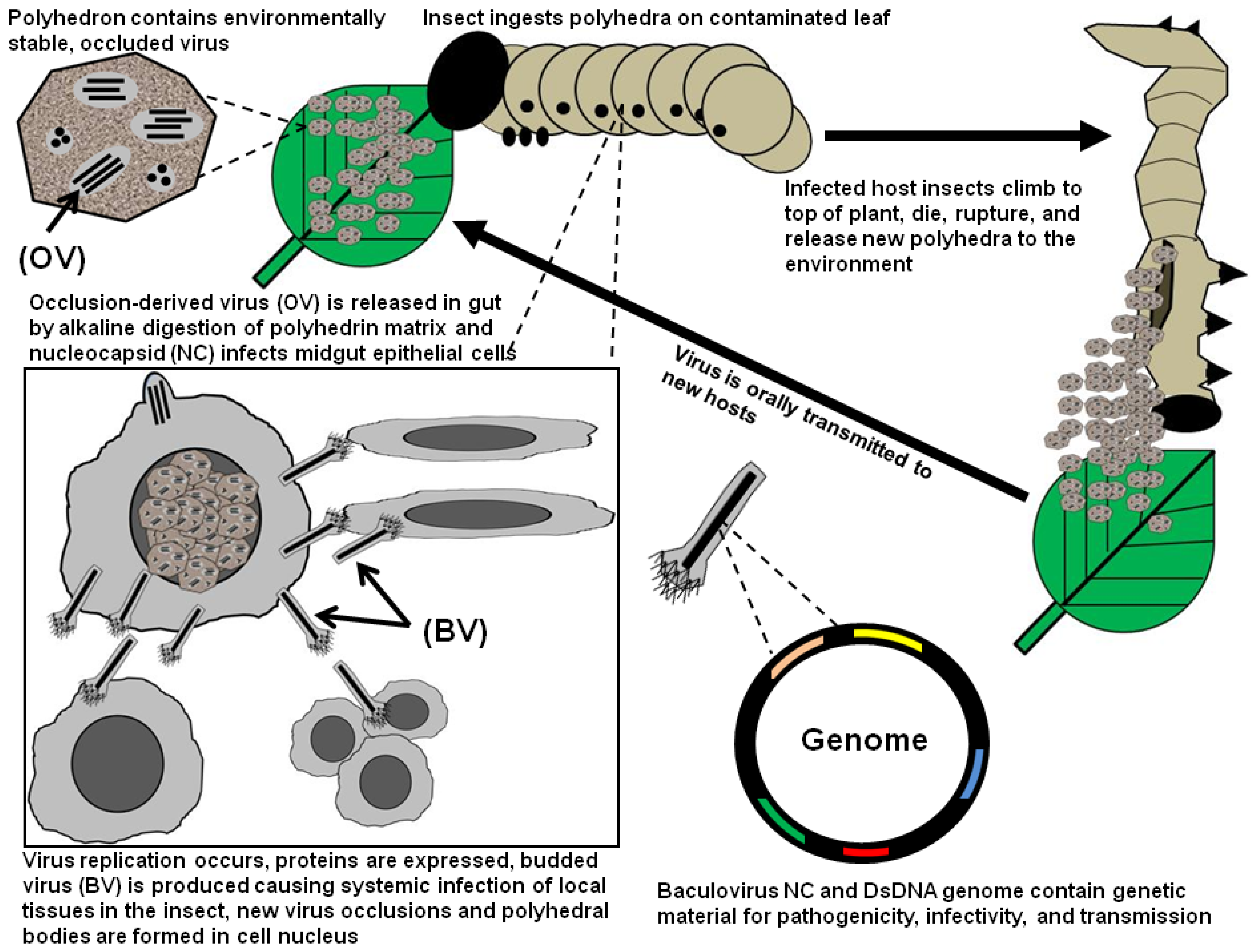
:1. Introduction

2. Insecticidal Peptides Expressed by Recombinant Baculoviruses
2.1. Neurotoxins

{kind=link}
| Recombinant Baculovirus | Insecticidal Peptide | Parental Virus/Host Species 1 | Promoter | Signal Peptide | Improvement vs. Control 2 | Reference 3 |
|---|---|---|---|---|---|---|
| vBeIt-1 | BeIT (scorpion) | AcMNPV/T. ni, G. mellonella | polh | None | No larval paralysis | [24] |
| vBeIt-2 | BeIT | AcMNPV/T. ni, G. mellonella | polh | Human β-interferon | No larval paralysis | [24] |
| vBeIt-3 | Polyhedrin-BeIT fusion | AcMNPV/T. ni, G. mellonella | polh | None | No larval paralysis | [24] |
| BmAaIT | AaIT(scorpion) | BmNPV/B. mori | polh | bombyxin | 40% | [25] |
| AcUW2(B).AaIT [ p10–AaIT; AcAaIT; vAaIT; Ac.AaIT(p10); vAcAaIT.p10] | AaIT | AcMNPV/H. armigera, H. virescens, S. exigua, T. ni | p10 | bombyxin | 12%–50% (n = 23, average 29%) | [26,27,28,29,30,31,32,33,34] |
| AcST-1 | AaIT | AcMNPV/T. ni | p10 | None | No neurotoxin secretion from infected cells | [35] |
| AcST-3 | AaIT | AcMNPV/T. ni | p10 | AcMNPV GP67 | 24%, 25% | [35,36] |
| vEV-Tox34 | TxP-I (mite) | AcMNPV/T. ni | polh (LSXIV) | native | Not estimated | [37] |
| vETL-Tox34 | TxP-I | AcMNPV/T. ni | ETL (AcMNPV ORF ac49) | native | Not estimated | [38] |
| vCappolh-Tox34 | TxP-I | AcMNPV/T. ni | vp39/polh fusion | native | Not estimated | [38] |
| vSp-tox34 | TxP-I | AcMNPV/T. ni, S. frugiperda | synXIV | native | 33%–59% (n = 7, average 44%) | [38,39,40] |
| vSp-tox21A | tox21A gene product (mite) | AcMNPV/T. ni | synXIV | native | 34%; 49% | [39,40] |
| vEV-HA5f10 | Dol m VB (hornet) | AcMNPV/T. ni | polh (LSXIV) | native | No larval paralysis or other effect | [41] |
| vEV-HA5f17 | Dol m VA(hornet) | AcMNPV/T. ni | polh (LSXIV) | native | No larval paralysis or other effect | [41] |
| AcLα22 (vAcLqαIT) | LqhαIT (scorpion) | AcMNPV/H. armigera, H. virescens | polh | native | 27%–41% (n = 3, average 34%) | [42,43] |
| AcNPV/RH1 | AaIT | AcMNPV/S. exigua | p10 | bombyxin | 31% | [27] |
| vSp-tox34m | TxP-I | AcMNPV/T. ni, S. frugiperda | synXIV | modified native | 44%; 54% | [40] |
| vSp-BSigtox34 | TxP-I | AcMNPV/T. ni | synXIV | Sarcophaga sarcotoxin IA | 26% | [40] |
| vSp-tox21A/tox34 | TxP-I | AcMNPV/T. ni | synXIV | tox21A | 36% | [40] |
| vSp-DCtox34 | TxP-I | AcMNPV/T. ni | synXIV | Drosophila cuticle protein II | 47% | [40] |
| vpHSP70tox34 | TxP-I | AcMNPV/T. ni, S. frugiperda | Drosophila hsp70 | native | 41%; 46% | [40] |
| vDA26tox34 | TxP-I | AcMNPV/T. ni, S. frugiperda | DA26 (AcMNPV ORF ac16) | native | 28%; 39% | [40] |
| vp6.9tox34 | TxP-I | AcMNPV/T. ni, S. frugiperda | p6.9 | native | 56%–59% (n = 4, average 58%) | [40] |
| vp6.9tox34m | TxP-I | AcMNPV/T. ni, S. frugiperda | p6.9 | modified native | 53%; 54% | [40] |
| TnNPV-AaIT (clones 1–4) | AaIT | “TnNPV”/T. ni | synXIV | AcMNPV GP67 | 20%–43% | [44] |
| vMAg4p+ | μ-Aga-IV (spider) | AcMNPV/T. ni, S. frugiperda | synXIV | Honeybee melittin | 15%–42% (n = 7, average 27%) | [30,45,46] |
| vSAt2p+ | As II (sea anemone) | AcMNPV/H. virescens, T. ni, S. frugiperda | synXIV | Sarcophaga sarcotoxin IA | 25%–38% (n = 6, average 33%) | [30,45] |
| vSSh1p+ | Sh I (sea anemone) | AcMNPV/T. ni, S. frugiperda | synXIV | Sarcophaga sarcotoxin IA | 35%–42% (n = 3, average 38%) | [30,45] |
| ie1–AaIT | AaIT | AcMNPV/H. virescens | hr5 enhancer/ ie-1 promoter | bombyxin | 10% | [28] |
| vAcTalTX-1 | TalTX-1 (spider) | AcMNPV/T. ni, S. exigua, H. virescens | polh | native | 18%–33% (n = 4, average 24% | [22] |
| vAcDTX9.2 | DTX9.2 (spider) | AcMNPV/T. ni, S. exigua, H. virescens | polh | T. ni AJSP-1 | 7%–24% (n = 4, average 14%) | [22] |
| HzEGTp6.9tox34 | TxP-I (inserted in egt gene) | HzSNPV/H. zea | p6.9 | native | 35% | [47] |
| HzEGThsptox34 | TxP-I (inserted in egt gene) | HzSNPV/H. zea | Drosophila hsp70 | native | 32% | [47] |
| HzEGTDA26tox34 | TxP-I (inserted in egt gene) | HzSNPV/H. zea | DA26 (AcMNPV ORF ac16) | native | 39% | [47] |
| vV8EE6.9tox34 | TxP-I (inserted in egt gene) | AcMNPV-V8/S. frugiperda | p6.9 | native | 38% | [48] |
| vV8EEHSPtox34 | TxP-I (inserted in egt gene) | AcMNPV-V8/S. frugiperda | Drosophila hsp70 | native | 46% | [48] |
| vhsMAg4p+ | μ-Aga-IV | AcMNPV/H. virescens, S. frugiperda, T. ni | Drosophila hsp70 | Honeybee melittin | 20%–71% (n = 11, average 47%) | [30,46] |
| vhsSAt2p+ | As II | AcMNPV/H. viresncens, S. frugiperda, T. ni | Drosophila hsp70 | Sarcophaga sarcotoxin IA | 50%–61% (n = 7, average 55%) | [30,46] |
| vDASAt2 | As II (in ac16-negative mutant) | AcMNPV/S. frugiperda, T. ni | synXIV | Sarcophaga sarcotoxin IA | 37%–40% (n = 4, average 39%) | [46] |
| vDAhsSAt2 | As II (in ac16-negative mutant) | AcMNPV/S. frugiperda, T. ni | Drosophila hsp70 | Sarcophaga sarcotoxin IA | 40%–53% (n = 5, average 47%) | [46] |
| vhsSSh1p+ | Sh I | AcMNPV/H. virescens, S. frugiperda, T. ni | Drosophila hsp70 | Sarcophaga sarcotoxin IA | 45%–55% (n = 4, average 50%) | [30,46] |
| vMAg4SAt2 | μ-Aga-IV + As II | AcMNPV/S. frugiperda, T. ni | synXIV (both toxins) | Honeybee melittin, Sarcophaga sarcotoxin IA | 38%–45% (n = 5; average 41%) | [30,46] |
| vhsMAg4SAt2 | μ-Aga-IV + As II | AcMNPV/S. frugiperda, T. ni | Drosophila hsp70 (both toxins) | Honeybee melittin, Sarcophaga sarcotoxin IA | 43%–57% (n = 5; average 49%) | [46] |
| AcLIT1.p35 | LqhIT1 (scorpion) | AcMNPV/H. armigera | p35 | native | 16% | [49] |
| AcLIT1.p10 (vAcLqIT1) | LqhIT1 | AcMNPV/H. armigera, H. virescens | p10 | native | 24%–28% (n = 3, average 27%) | [43,49] |
| AcLIT2.pol (vAcLqIT2) | LqhIT2 (scorpion) | AcMNPV/H. armigera, H. virescens | polh | native | 17%–32% (n = 3, average 24%) | [43,49] |
| RcHzSNPV (RcHzLqh) | LqhIT2 | HzSNPV/H. zea, H. virescens | hr5 enhancer/ ie-1 promoter | bombyxin | 31%–44% (n = 11, average 38%) | [32,50] |
| AcTOX34.4 | TxP-1 ( tox34.4) | AcMNPV/T. ni | p10 | native | Approx. 30%–50% | [51] |
| BmLqhIT2 | LqhIT2 | BmNPV/B. mori | polh | bombyxin | Not estimated | [52] |
| Ro6.9AaIT | AaIT | RoMNPV/H. virescens, H. zea, O. nubilalis | p6.9 | bombyxin | 19%–37% (n = 3, average 30%) | [53] |
| Ro6.9LqhIT2 | LqhIT2 | RoMNPV/H. virescens, H. zea, O. nubilalis | p6.9 | bombyxin | 27%–42% (n = 3, average 37%) | [53] |
| AcUW21.AaIT | AaIT | AcMNPV/H. virescens | p10 | bombyxin | 27% | [53] |
| AcMLF9.AaIT | AaIT | AcMNPV/H. virescens | p6.9 | bombyxin | 25%, 26% | [53,54] |
| AcUW21.LqhIT2 | LqhIT2 | AcMNPV/H. virescens | p10 | bombyxin | 34% | [53] |
| AcMLF9.LqhIT2 | LqhIT2 | AcMNPV/H. virescens | p6.9 | bombyxin | 33%, 34% | [53,54] |
| HaCXW2 | AaIT (inserted in the egt gene) | HearSNPV/H. armigera | polh | bombyxin | 8%; 13% (relative to egt-negative control virus) | [55,56] |
| vAcLqIT1-IT2 | LqhIT1 and LqhIT2 | AcMNPV/H. armigera, H. virescens | p10 (LqhIT1), polh (LqhIT2) | native | 24%, 41% | [43] |
| vAcLqαIT-IT2 | LqhαIT and LqhIT2 | AcMNPV/H. armigera, H. virescens | polh (both toxins) | native | 29%, 43% | [43] |
| Ac.LqhIT2(hr5/ie1) | LqhIT2 | AcMNPV/H. virescens | hr5 enhancer/ ie-1 promoter | native, Drosophila cuticle protein II, M. sexta adipokinetic hormone, L. cuprina chymotrypsin, AcMNPV GP67, BjIT, AaIT | Not estimated | [33] |
| Ac.LqhIT2(hr5/39K) | LqhIT2 | AcMNPV/H. virescens | hr5 enhancer/39k promoter | bombyxin | Not estimated | [33] |
| Ac.LqhIT2(hr5/lef3) | LqhIT2 | AcMNPV/H. virescens | hr5 enhancer/lef-3 promoter | bombyxin | Not estimated | [33] |
| Ac.LqhIT2(hr5/ie1) | LqhIT2 | AcMNPV/H. virescens, S. exigua, T. ni | hr5 enhancer/ ie-1 promoter | bombyxin | 37%–56% (n = 10, average 47%) | [33] |
| Ac.AaIT(hr5/ie1) | AaIT | AcMNPV/H. virescens, S. exigua, T. ni | hr5 enhancer/ ie-1 promoter | bombyxin | 13%–46% (n = 10, average 31%) | [33] |
| HaSNPV-AaIT | AaIT (inserted in the egt gene) | HearSNPV/H. armigera | Tandem polh and p6.9 promoters | bombyxin | 4%–27% (relative to egt-negative control virus ; n = 9, average 13%); 26% relative to wild-type HearSNPV | [57,58] |
| Px | Poneratoxin (ant) | AcMNPV/S. frugiperda | polh | none | No improvement | [59] |
| SPx | Poneratoxin | AcMNPV/S. frugiperda | polh | AcMNPV GP67 | 16% | [59] |
| AcIT6 | BotIT6 (scorpion) | AcMNPV/S. littoralis | polh | AcMNPV EGT | 33% | [60] |
| vAP10IT2 | LqhIT2 | AcMNPV/P. xylostella, S. exigua, T. ni | p10 | bombyxin | 8%–16% (n = 4, average 12%) | [61,62] |
| vAPcmIT2 | LqhIT2 | AcMNPV/P. xylostella, S. exigua, T. ni | Cytomegalo-virus minimal promoter | bombyxin | 10%–25% (n = 4, average 15%) | [61,62] |
| vAcAaIT.39K | AaIT | AcMNPV/H. armigera | 39k | bombyxin | 22% | [34] |
| ButaIT-NPV | ButaIT (scorpion) | AcMNPV/H. virescens | p10 | bombyxin | 43% | [63] |
| P10IT2 | LqhIT2 | AcMNPV/T. ni | p10 | bombyxin | 28% | [64] |
| Ppag90IT2 | LqhIT2 | AcMNPV/T. ni | HzNV-1 pag90 | bombyxin | 50% | [64] |
| AcMNPV- BmK IT | BmK IT (scorpion) | AcMNPV/S. exigua | ie-1 | native | 13% | [65] |
| SpltNPV-Δegt-BmK | BmK ITa1(inserted in egt gene) | SpltNPV/S. litura | polh | SpltNPV F protein | 15%; 28% | [66] |
2.2. Physiological Regulators
| Recombinant Baculovirus | Physiological Regulator | Parental Virus/Host Species 1 | Promoter | Signal Peptide | Improvement vs. Control 2 | Reference 3 |
|---|---|---|---|---|---|---|
| BmDH5 | M. sexta diuretic hormone | BmNPV/B. mori | polh | Drosophila cuticle protein II | Approximately 20% | [25,69] |
| AcRP23.JHE | H. virescens juvenile hormone esterase (JHE) | AcMNPV/T. ni | polh | native | Not determined | [70] |
| AcUW2(B).JHE (AcJHE) | H. virescens JHE | AcMNPV/H. virescens, T. ni | p10 | native | No improvement | [26,31,71,72,73] |
| vJHEEGTD | H. virescens JHE | AcMNPV (egt deletion mutant)/T. ni | Tandem polh (LSXIV) and synXIV promoters | native | No improvement relative to egt deletion control virus | [74] |
| vEHEGTD | M. sexta eclosion hormone | AcMNPV (egt deletion mutant)/S. frugiperda | Tandem polh (LSXIV) and synXIV promoters | native | No improvement relative to egt deletion control virus | [75] |
| AcJHE-SG | H. virescens JHE-SG mutant | AcMNPV/H. virescens, T. ni | p10 | native | 16%–31% (n = 3, average 25%) 16%; 29%; 31% | [31,72] |
| AcJHE-HK | H. virescens JHE-HK mutant | AcMNPV/ H. virescens | p10 | native | No improvement | [31,72] |
| AcJHE-RH | H. virescens JHE-RH mutant | AcMNPV/ H. virescens | p10 | native | No improvement | [72] |
| AcJHE-DN | H. virescens JHE-DN mutant | AcMNPV/ H. virescens | p10 | native | No improvement | [72] |
| vWTPTTHM | B. mori prothoracicotropic hormone | AcMNPV/S. frugiperda | Tandem polh (LSXIV) and synXIV promoters | Sarcophaga sarcotoxin IA | No improvement | [76] |
| vEGT-PTTHM | B. mori prothoracicotropic hormone | AcMNPV ( egt deletion mutant)/S. frugiperda | Tandem polh (LSXIV) and synXIV promoters | Sarcophaga sarcotoxin IA | No improvement | [76] |
| AcJHE.KK ( p10-JHE-KK; AcUW21.JHE-KK) | H. virescens JHE-KK mutant | AcMNPV/ H. virescens, T. ni | p10 | native | 0%–25% (n = 7, average 13%) | [28,29,31,73] |
| ie1–JHE–KK | H. virescens JHE-KK mutant | AcMNPV/H. virescens | hr5 enhancer/ ie-1 promoter | native | No improvement | [28] |
| AcBX-PBAN | H. zea pheromone biosynthesis activating neuropeptide | AcMNPV/T. ni | polh | bombyxin | 19%; 26% | [77] |
| AcJHE-29 | H. virescens JHE-29 mutant | AcMNPV/ H. virescens , T. ni | p10 | native | No improvement | [73] |
| AcJHE-524 | H. virescens JHE-524 mutant | AcMNPV/ H. virescens , T. ni | p10 | native | No improvement | [73] |
| AcJHE-KSK | H. virescens JHE-KSK mutant | AcMNPV/H. virescens | p10 | native | 17% | [31] |
| AcJHE-KHK | H. virescens JHE-KHK mutant | AcMNPV/H. virescens | p10 | native | No improvement | [31] |
| AcHezK-II S | H. zea helicokinin-II (one copy) | AcMNPV/H. zea | polh | Human placental alkaline phosphatase | Not estimated | [78] |
| AcHezK-II M | H. zea helicokinin-II (two copies) | AcMNPV/H. zea | polh and p10 | Human placental alkaline phosphatase | Not estimated | [78] |
| Recombinant Baculovirus | Degradative Enzyme | Parental Virus/Host Species 1 | Promoter | Signal Peptide | Improvement vs. Control 2 | Reference 3 |
|---|---|---|---|---|---|---|
| vAcMNPV.chi | M. sexta chitinase | AcMNPV/S. frugiperda | polh | native | 22%; 23% | [80] |
| AcIE1TV3.STR1 | Rat stromelysin-1 | AcMNPV/H. virescens | hr5 enhancer/ ie-1 promoter | native | No improvement | [54] |
| AcMLF9.STR1 | Rat stromelysin-1 | AcMNPV/H. virescens | p6.9 | native | No improvement | [54] |
| AcIE1TV3.GEL | Human gelatinase A | AcMNPV/H. virescens | hr5 enhancer/ ie-1 promoter | native | No improvement | [54] |
| AcMLF9.GEL | Human gelatinase A | AcMNPV/H. virescens | p6.9 | native | 12% | [54] |
| AcIE1TV3.ScathL | S. peregrina cathepsin L | AcMNPV/H. virescens | hr5 enhancer/ ie-1 promoter | native | No improvement | [54] |
| AcMLF9.ScathL | S. peregrina cathepsin L | AcMNPV/H. virescens | p6.9 | native | 49%; 51% | [54,81] |
| AcMNPV-enMP2 | MacoNPV-A enhancin | AcMNPV/T. ni | Native | None | No improvement at same effective dose as control | [82] |
| BacVEFPol | TnGV enhancin | AcMNPV/T. ni | p10 | None | 17% | [83] |
| AcMLF9.ScathL.CAT | S. peregrina cathepsin L | AcMNPV/H. virescens | p6.9 | native | 54% | [81] |
| AcMLF9.ScathL.hsp70/lacZ | S. peregrina cathepsin L | AcMNPV/H. virescens | p6.9 | native | 51% | [81] |
| AcBAC–polhvfgf | AcMNPV viral fibroblast growth factor (stimulates protease synthesis/activation) | AcMNPV/S. frugiperda, T. ni | polh | none | 20%; 23% | [84] |
| AcMNPV-GFP-HCB-Polh | H. armigera cathepsin B | AcMNPV/H. armigera | polh | native | 11% | [85] |
| HearNPV-cathL | S. peregrina cathepsin L (inserted in egt gene) | HearSNPV/H. armigera | Tandem polh and p6.9 | native | 26%; 26% | [58] |
| vSynScathL | S. peregrina cathepsin L | AcMNPV/S. frugiperda | synXIV | native | 26%; 65% | [86] |
| vSynKerat | Aspergillus fumigates keratinase | AcMNPV/S. frugiperda | synXIV | native | 33%; 48% | [86] |
2.3. Degradative Enzymes
2.4. Other Insecticidal Proteins
| Recombinant Baculovirus | Insecticidal Protein | Parental Virus/Host Species 1 | Promoter | Signal Peptide | Improvement vs. Control 2 | Reference 3 |
|---|---|---|---|---|---|---|
| Ac(PH+)Bt | B. thuringiensis CryIA(c) | AcMNPV/T. ni | polh | none | Not determined | [99] |
| AcBtm | B. thuringiensis CryIA(b) (full-length) | AcMNPV/H. virescens | polh | none | No improvement | [102] |
| AcBt5 | B. thuringiensis CryIA(b) (N-terminal truncated) | AcMNPV/H. virescens | polh | none | No improvement | [102] |
| AcBt3 | B. thuringiensis CryIA(b) (C-terminal truncated) | AcMNPV/H. virescens | polh | none | No improvement | [102] |
| AcBt5/3 | B. thuringiensis CryIA(b) (N- and C-terminal truncated) | AcMNPV/H. virescens | polh | none | No improvement | [102] |
| BV13T | Z. mays URF13 | AcMNPV/T. ni | polh | native | Not estimated | [106] |
| Bv13.3940 | Z. mays URF13 mutant | AcMNPV/T. ni | polh | native | Not estimated | [106] |
| AcNPV/JM2 | B. thuringiensis CryIA(b) (C-terminal truncated) | AcMNPV/S. exigua | p10 | none | No improvement | [27] |
| AcNPV/FW3 | B. thuringiensis CryIA(b) (C-terminal truncated) | AcMNPV/S. exigua | p10 | H. virescens juvenile hormone esterase | No improvement | [27] |
| vHSA50L | PBCV-1 cv-PDG (pyrimidine dimer-specific glycosylase) | AcMNPV/S. frugiperda, T. ni | Drosophila hsp70 | none | No improvement vs. T. ni; 48% vs. S. frugiperda | [107] |
| vHSA50LORF | PBCV-1 cv-PDG | AcMNPV/S. frugiperda, T. ni | Drosophila hsp70 | none | No improvement vs. T. ni; 41% vs. S. frugiperda | [107] |
| ColorBtrus | AcMNPV polyhedrin-B. thuringiensis Cry1A(c)-green fluorescent protein fusion | AcMNPV/P. xylostella | polh | None | 63% | [105] |
3. Factors Involved in Successful Improvement of Baculovirus Pesticidal Performance with Insectidal Protein Expression
3.1. Choice of Promoter
3.2. Choice of Signal Peptide
3.3. Translational Efficiency and Post-Translational Processing
3.4. Interaction with Host Physiology
4. Looking Forward: Insecticidal RNAs?
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Huber, J. Use of baculoviruses in pest management programs. In The Biology of Baculoviruses. Volume II. Practical Application for Insect Control; Granados, R.R., Federici, B.A., Eds.; CRC Press: Boca Raton, FL, USA, 1986; pp. 181–202. [Google Scholar]
- Herniou, E.A.; Arif, B.M.; Becnel, J.J.; Blissard, G.W.; Bonning, B.; Harrison, R.L.; Jehle, J.A.; Theilmann, D.A.; Vlak, J.M. Baculoviridae. In Virus Taxonomy: Ninth Report of the International Committee on Taxonomy of Viruses; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Elsevier: Oxford, UK, 2011; pp. 163–174. [Google Scholar]
- Harrison, R.L.; Hoover, K. Baculoviruses and other occluded insect viruses. In Insect Pathology, Second Edition; Vega, F.E., Kaya, H.K., Eds.; Academic Press: Boston, MA, USA, 2012; pp. 73–131. [Google Scholar]
- Armenta, R.; Martinez, A.M.; Chapman, J.W.; Magallanes, R.; Goulson, D.; Caballero, P.; Cave, R.D.; Cisneros, J.; Valle, J.; Castillejos, V.; et al. Impact of a nucleopolyhedrovirus bioinsecticide and selected synthetic insecticides on the abundance of insect natural enemies on maize in southern Mexico. J. Econ. Entomol. 2003, 96, 649–661. [Google Scholar] [CrossRef] [PubMed]
- Harrap, K.A. The structure of nuclear polyhedrosis viruses. I. The inclusion body. Virology 1972, 50, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Thompson, C.G.; Scott, D.W.; Wickman, B.E. Long-term persistence of the nuclear polyhedrosis virus of the Douglas-fir tussock moth, Orgyia pseudotsugata (Lepidoptera: Lymantriidae), in forest soil. Environ. Entomol. 1981, 10, 254–255. [Google Scholar] [CrossRef]
- Pritchett, D.W.; Young, S.Y.; Yearian, W.C. Dissolution of Autographa californica nuclear polyhedrosis virus polyhedra by the digestive fluid of Trichoplusia ni (Lepidoptera: Noctuidae) larvae. J. Invert. Path. 1982, 39, 354–361. [Google Scholar] [CrossRef]
- Kawanishi, C.Y.; Summers, M.D.; Stoltz, D.B.; Arnott, H.J. Entry of an insect virus in vivo by fusion of viral envelope and microvillus membrane. J. Invert. Path. 1972, 20, 104–108. [Google Scholar] [CrossRef]
- Granados, R.R.; Lawler, K.A. In vivo pathway of Autographa californica baculovirus invasion and infection. Virology 1981, 108, 297–308. [Google Scholar] [CrossRef] [PubMed]
- Summers, M.D.; Volkman, L.E. Comparison of biophysical and morphological properties of occluded and extracellular nonoccluded baculovirus from in vivo and in vitro host systems. J. Virol. 1976, 17, 962–972. [Google Scholar] [PubMed]
- Goodwin, R.H.; Vaughn, J.L.; Adams, J.R.; Louloudes, S.J. Replication of a nuclear polyhedrosis virus in an established insect cell line. J. Invertebr. Pathol. 1970, 16, 284–288. [Google Scholar] [CrossRef] [PubMed]
- Williams, G.V.; Rohel, D.Z.; Kuzio, J.; Faulkner, P. A cytopathological investigation of Autographa californica nuclear polyhedrosis virus p10 gene functions using insetion/deletion mutants. J. Gen. Virol. 1989, 70, 187–202. [Google Scholar] [CrossRef] [PubMed]
- Hawtin, R.E.; Zarkowska, T.; Arnold, K.; Thomas, C.J.; Gooday, G.W.; King, L.A.; Kuzio, J.A.; Possee, R.D. Liquefaction of Autographa californica nucleopolyhedrovirus-infected insects is dependent on the integrity of virus-encoded chitinase and cathepsin genes. Virology 1997, 238, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Harper, J.D. Food consumption by cabbage loopers infected with nuclear polyhedrosis virus. J. Invert. Path. 1973, 21, 191–197. [Google Scholar] [CrossRef]
- Black, B.C.; Brennan, L.A.; Dierks, P.M.; Gard, I.E. Commercialization of baculoviral insecticides. In The Baculoviruses; Miller, L.K., Ed.; Plenum Press: New York, NY, USA, 1997; pp. 341–387. [Google Scholar]
- Miller, L.K.; Lingg, A.J.; Bulla, L.A., Jr. Bacterial, viral, and fungal insecticides. Science 1983, 219, 715–721. [Google Scholar] [CrossRef] [PubMed]
- O'Reilly, D.R.; Miller, L.K. A baculovirus blocks insect molting by producing ecdysteroid udp-glucosyl transferase. Science 1989, 245, 1110–1112. [Google Scholar] [CrossRef] [PubMed]
- O'Reilly, D.R.; Miller, L.K. Improvement of a baculovirus pesticide by deletion of the egt gene. Biotechnology 1991, 9, 1086–1089. [Google Scholar] [CrossRef]
- Hunter-Fujita, F.R.; Entwistle, P.F.; Evans, H.F.; Crook, N.E. Insect Viruses and Pest Management; John Wiley & Sons: Chichester, UK, 1998. [Google Scholar]
- Kamita, S.G.; Kang, K.-D.; Hammock, B.D.; Inceoglu, A.B. Genetically modified baculoviruses for pest insect control. In Comprehensive Molecular Insect Science; Gilbert, L.I., Iatrou, K., Gill, S.S., Eds.; Pergamon Press: Oxford, UK, 2005; Volume 6, pp. 271–322. [Google Scholar]
- Kamita, S.G.; Kang, K.-D.; Hammock, B.D.; Inceoglu, A.B. Addendum: Genetically modified baculoviruses for pest insect control. In Insect Control: Biological and Synthetic Agents; Gilbert, L.I., Gill, S.S., Eds.; Academic Press: London, UK, 2010; pp. 383–386. [Google Scholar]
- Hughes, P.R.; Wood, H.A.; Breen, J.P.; Simpson, S.F.; Duggan, A.J.; Dybas, J.A. Enhanced bioactivity of recombinant baculoviruses expressing insect-specific spider toxins in lepidopteran crop pests. J. Invertebr. Pathol. 1997, 69, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Van Beek, N.A.M.; Hughes, P.R. The response time of insect larvae infected with recombinant baculoviruses. J. Invertebr. Pathol. 1998, 72, 338–347. [Google Scholar] [CrossRef] [PubMed]
- Carbonell, L.F.; Hodge, M.R.; Tomalski, M.D.; Miller, L.K. Synthesis of a gene coding for an insect-specific scorpion neurotoxin and attempts to express it using baculovirus vectors. Gene 1988, 73, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Maeda, S.; Volrath, S.L.; Hanzlik, T.N.; Harper, S.A.; Majima, K.; Maddox, D.W.; Hammock, B.D.; Fowler, E. Insecticidal effects of an insect-specific neurotoxin expressed by a recombinant baculovirus. Virology 1991, 184, 777–780. [Google Scholar] [CrossRef] [PubMed]
- McCutchen, B.F.; Choudary, P.V.; Crenshaw, R.; Maddox, D.; Kamita, S.G.; Palekar, N.; Volrath, S.; Fowler, E.; Hammock, B.D.; Maeda, S. Development of a recombinant baculovirus expressing an insect-selective neurotoxin: Potential for pest control. Biotechnology 1991, 9, 848–852. [Google Scholar] [CrossRef] [PubMed]
- Martens, J.W.; Knoester, M.; Weijts, F.; Groffen, S.J.; Hu, Z.; Bosch, D.; Vlak, J.M. Characterization of baculovirus insecticides expressing tailored Bacillus thuringiensis CryIA(B) crystal proteins. J. Invert. Path. 1995, 66, 249–257. [Google Scholar] [CrossRef]
- Jarvis, D.L.; Reilly, L.M.; Hoover, K.; Hammock, B.; Guarino, L.A. Construction and characterization of immediate-early baculovirus insecticides. Biol. Control. 1996, 7, 228–235. [Google Scholar] [CrossRef]
- Kunimi, Y.; Fuxa, J.R.; Hammock, B.D. Comparison of wild type and genetically engineered nuclear polyhedrosis viruses of Autographa californica for mortality, virus replication and polyhedra production in Trichoplusia ni larvae. Entomol. Exp. Appl. 1996, 81, 251–257. [Google Scholar] [CrossRef]
- Popham, H.J.R.; Prikhod’ko, G.G.; Felcetto, T.J.; Ostlind, D.A.; Warmke, J.W.; Cohen, C.J.; Miller, L.K. Effect of deltamethrin treatment on lepidopteran larvae infected with baculoviruses expressing insect-selective toxins μ-Aga-IV, As II, or Sh 1. Biol. Control. 1998, 12, 79–87. [Google Scholar] [CrossRef]
- Van Meer, M.M.M.; Bonning, B.C.; Ward, V.K.; Vlak, J.M.; Hammock, B.D. Recombinant, catalytically inactive juvenile hormone esterase enhances efficacy of baculovirus insecticides. Biol. Control. 2000, 19, 191–199. [Google Scholar] [CrossRef]
- Ignoffo, C.M.; Wong, J.F.H.; McIntosh, A.H.; McCutchen, W.F.; Grasela, J.J.; Goodman, C.L.; Saathoff, S.G. Sensitivity of heliothine larvae to wild-type and recombinant strains of a single nucleocapsid HzSNPV) and a multiple nucleocapsid (AcMNPV) baculovirus. Appl. Entomol. Zool. 2001, 36, 127–136. [Google Scholar]
- Van Beek, N.; Lu, A.; Presnail, J.; Davis, D.; Greenamoyer, C.; Joraski, K.; Moore, L.; Pierson, M.; Herrmann, R.; Flexner, L.; et al. Effect of signal sequence and promoter on the speed of action of a genetically modified Autographa californica nucleopolyhedrovirus expressing the scorpion toxin LqhIT2. Biol. Control. 2003, 27, 53–64. [Google Scholar] [CrossRef]
- Regev, A.; Rivkin, H.; Gurevitz, M.; Chejanovsky, N. New measures of insecticidal efficacy and safety obtained with the 39k promoter of a recombinant baculovirus. FEBS Lett. 2006, 580, 6777–6782. [Google Scholar] [CrossRef] [PubMed]
- Stewart, L.M.; Hirst, M.; Lopez Ferber, M.; Merryweather, A.T.; Cayley, P.J.; Possee, R.D. Construction of an improved baculovirus insecticide containing an insect-specific toxin gene. Nature 1991, 352, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Crespo, P.; Sait, S.M.; Hails, R.S.; Cory, J.S. Behavior of a recombinant baculovirus in lepidopteran hosts with different susceptibilities. Appl. Environ. Microbiol. 2001, 67, 1140–1146. [Google Scholar] [CrossRef] [PubMed]
- Tomalski, M.D.; Miller, L.K. Insect paralysis by baculovirus-mediated expression of a mite neurotoxin gene. Nature 1991, 352, 82–85. [Google Scholar] [CrossRef] [PubMed]
- Tomalski, M.D.; Miller, L.K. Expression of a paralytic neurotoxin gene to improve insect baculoviruses as biopesticides. Biotechnology 1992, 10, 545–549. [Google Scholar] [CrossRef]
- Tomalski, M.D.; Hutchinson, K.; Todd, J.; Miller, L.K. Identification and characterization of tox21a: A mite cDNA encoding a paralytic neurotoxin related to TxP-I. Toxicon 1993, 31, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Lu, A.; Seshagiri, S.; Miller, L.K. Signal sequence and promoter effects on the efficacy of toxin-expressing baculoviruses as biopesticides. Biol. Control. 1996, 7, 320–332. [Google Scholar] [CrossRef]
- Tomalski, M.D.; King, T.P.; Miller, L.K. Expression of hornet genes encoding venom allergen antigen 5 in insects. Arch. Insect Biochem. Physiol. 1993, 22, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Chejanovsky, N.; Zilberberg, N.; Rivkin, H.; Zlotkin, E.; Gurevitz, M. Functional expression of an alpha anti-insect scorpion neurotoxin in insect cells and lepidopterous larvae. FEBS Lett. 1995, 376, 181–184. [Google Scholar] [CrossRef] [PubMed]
- Regev, A.; Rivkin, H.; Inceoglu, B.; Gershburg, E.; Hammock, B.D.; Gurevitz, M.; Chejanovsky, N. Further enhancement of baculovirus insecticidal efficacy with scorpion toxins that interact cooperatively. FEBS Lett. 2003, 537, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Yao, B.; Pang, Y.; Fan, Y.; Zhao, R.; Yang, Y.; Wang, T. Construction of an insecticidal baculovirus expressing insect-specific neurotoxin AaIT. Sci. China C Life Sci. 1996, 39, 199–206. [Google Scholar] [PubMed]
- Prikhod’ko, G.G.; Robson, M.; Warmke, J.W.; Cohen, C.J.; Smith, M.M.; Wang, P.; Warren, V.; Kaczorowski, G.; van der Ploeg, L.H.T.; Miller, L.K. Properties of three baculovirus-expressing genes that encode insect-selective toxins: μ-Aga-IV, As II, and Sh I. Biol. Control. 1996, 7, 236–244. [Google Scholar] [CrossRef]
- Prikhod’ko, G.G.; Popham, H.J.R.; Felcetto, T.J.; Ostlind, D.A.; Warren, V.A.; Smith, M.M.; Garsky, V.M.; Warmke, J.W.; Cohen, C.J.; Miller, L.K. Effects of simultaneous expression of two sodium channel toxin genes on the properties of baculoviruses as biopesticides. Biol. Control. 1998, 12, 66–78. [Google Scholar] [CrossRef]
- Popham, H.J.R.; Li, Y.; Miller, L.K. Genetic improvement of Helicoverpa zea nuclear polyhedrosis virus as a biopesticide. Biol. Control. 1997, 10, 83–91. [Google Scholar] [CrossRef]
- Popham, H.J.R.; Pellock, B.J.; Robson, M.; Dierks, P.M.; Miller, L.K. Characterization of a variant of Autographa californica nuclear polyhedrosis virus with a nonfunctional orf 603. Biol. Control. 1998, 12, 223–230. [Google Scholar] [CrossRef]
- Gershburg, E.; Stockholm, D.; Froy, O.; Rashi, S.; Gurevitz, M.; Chejanovsky, N. Baculovirus-mediated expression of a scorpion depressant toxin improves the insecticidal efficacy achieved with excitatory toxins. FEBS Lett. 1998, 422, 132–136. [Google Scholar] [CrossRef] [PubMed]
- Ignoffo, C.M.; Wong, J.F.; Saathoff, S.G. Mortality and feeding of a mid-stadium larvae of Helicoverpa zea and Heliothis virescens fed a wild strain or a recombinant strain of Baculovirus heliothis expressing an insect-specific toxin of the scorpion Leirus quinquestriatus hebraeus. Appl. Entomol. Zool. 1999, 34, 279–283. [Google Scholar]
- Burden, J.P.; Hails, R.S.; Windass, J.D.; Suner, M.M.; Cory, J.S. Infectivity, speed of kill, and productivity of a baculovirus expressing the itch mite toxin TxP-1 in second and fourth instar larvae of Trichoplusia ni. J. Invertebr. Pathol. 2000, 75, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Imai, N.; Ali, S.E.; El-Singaby, N.R.; Iwanaga, M.; Matsumoto, S.; Iwabuchi, K.; Maeda, S. Insecticidal effects of a recombinant baculovirus expressing scorption toxin LqhIT2. J. Seric. Sci. Jpn. 2000, 69, 197–205. [Google Scholar]
- Harrison, R.L.; Bonning, B.C. Use of scorpion neurotoxins to improve the insecticidal activity of Rachiplusia ou multicapsid nucleopolyhedrovirus. Biol. Control. 2000, 17, 191–201. [Google Scholar] [CrossRef]
- Harrison, R.L.; Bonning, B.C. Use of proteases to improve the insecticidal activity of baculoviruses. Biol. Control. 2001, 20, 199–209. [Google Scholar] [CrossRef]
- Chen, X.; Sun, X.; Hu, Z.; Li, M.; O’Reilly, D.R.; Zuidema, D.; Vlak, J.M. Genetic engineering of Helicoverpa. armigera single-nucleocapsid nucleopolyhedrovirus as an improved pesticide. J. Invertebr. Pathol. 2000, 76, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Chen, X.; Zhang, Z.; Wang, H.; Bianchi, F.J.; Peng, H.; Vlak, J.M.; Hu, Z. Bollworm responses to release of genetically modified Helicoverpa armigera nucleopolyhedroviruses in cotton. J. Invertebr. Pathol. 2002, 81, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.L.; Wang, H.L.; Sun, X.C.; Chen, X.W.; Peng, C.M.; Pan, D.M.; Jehle, J.A.; van der Werf, W.; Vlak, J.M.; Hu, Z.H. Biological activity and field efficacy of a genetically modified Helicoverpa armigera SNPV expressing an insect-selective toxin from a chimeric promoter. Biol. Control. 2004, 29, 124–137. [Google Scholar] [CrossRef]
- Sun, X.; Wua, D.; Sun, X.; Jin, L.; Mab, Y.; Bonning, B.C.; Peng, H.; Hu, Z. Impact of Helicoverpa armigera nucleopolyhedroviruses expressing a cathepsin l-like protease on target and nontarget insect species on cotton. Biol. Control. 2009, 49, 77–83. [Google Scholar] [CrossRef]
- Szolajska, E.; Poznanski, J.; Ferber, M.L.; Michalik, J.; Gout, E.; Fender, P.; Bailly, I.; Dublet, B.; Chroboczek, J. Poneratoxin, a neurotoxin from ant venom. Structure and expression in insect cells and construction of a bio-insecticide. Eur. J. Biochem. 2004, 271, 2127–2136. [Google Scholar] [CrossRef] [PubMed]
- Bel Haj Rhouma, R.; Cerutti-Duonor, M.; Benkhadir, K.; Goudey-Perriere, F.; El Ayeb, M.; Lopez-Ferber, M.; Karoui, H. Insecticidal effects of Buthus occitanus tunetanus BotIT6 toxin expressed in Escherichia coli and baculovirus/insect cells. J. Insect Physiol. 2005, 51, 1376–1383. [Google Scholar] [CrossRef] [PubMed]
- Tuan, S.J.; Hou, R.F.; Kao, S.S.; Lee, C.F.; Chao, Y.C. Improved plant protective efficacy of a baculovirus using an early promoter to drive insect-specific neurotoxin expression. Bot. Bull. Acad. Sin. 2005, 46, 11–20. [Google Scholar]
- Tuan, S.J.; Hou, R.F.; Lee, C.F.; Chao, Y.C. High level production of polyhedra in a scorpion toxin containing recombinant baculovirus for better control of insect pests. Bot. Stud. 2007, 48, 273–281. [Google Scholar]
- Rajendra, W.; Hackett, K.J.; Buckley, E.; Hammock, B.D. Functional expression of lepidopteran-selective neurotoxin in baculovirus: Potential for effective pest management. Biochim. Biophys. Acta 2006, 1760, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Jinn, T.R.; Tu, W.C.; Lu, C.I.; Tzen, J.T. Enhancing insecticidal efficacy of baculovirus by early expressing an insect neurotoxin, lqhit2, in infected Trichoplusia ni larvae. Appl. Microbiol. Biotechnol. 2006, 72, 1247–1253. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Zheng, B.; Fu, Y.; Sun, Y.; Liang, A. Baculovirus-mediated expression of a chinese scorpion neurotoxin improves insecticidal efficacy. Chin. Sci. Bull. 2008, 53, 1855–1860. [Google Scholar] [CrossRef]
- Tang, X.X.; Sun, X.L.; Pu, G.Q.; Wang, W.B.; Zhang, C.X.; Zhu, J. Expression of a neurotoxin gene improves the insecticidal activity of Spodoptera litura nucleopolyhedrovirus (SpltNPV). Virus Res. 2011, 159, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Sun, X.; van der Werf, W.; Vlak, J.M.; Hu, Z. Field inactivation of wild-type and genetically modified Helicoverpa armigera single nucleocapsid nucleopolyhedrovirus in cotton. Biocontrol. Sci. Technol. 2004, 14, 185–192. [Google Scholar] [CrossRef]
- Zlotkin, E.; Kadouri, D.; Gordon, D.; Pelhate, M.; Martin, M.F.; Rochat, H. An excitatory and a depressant insect toxin from scorpion venom both affect sodium conductance and possess a common binding site. Arch. Biochem. Biophys. 1985, 240, 877–887. [Google Scholar] [CrossRef] [PubMed]
- Maeda, S. Increased insecticidal effect by a recombinant baculovirus carrying a synthetic diuretic hormone gene. Biochem. Biophys. Res. Commun. 1989, 165, 1177–1183. [Google Scholar] [CrossRef] [PubMed]
- Hammock, B.D.; Bonning, B.C.; Possee, R.D.; Hanzlik, T.N.; Maeda, S. Expression and effects of the juvenile hormone esterase in a baculovirus vector. Nature 1990, 344, 458–461. [Google Scholar] [CrossRef]
- Bonning, B.C.; Hirst, M.; Possee, R.D.; Hammock, B.D. Further development of a recombinant baculovirus insecticide expressing the enzyme juvenile hormone esterase from Heliothis virescens. Insect Biochem. Mol. Biol. 1992, 22, 453–458. [Google Scholar] [CrossRef]
- Bonning, B.C.; Hoover, K.; Booth, T.F.; Duffey, S.; Hammock, B.D. Development of a recombinant baculovirus expressing a modified juvenile hormone esterase with potential for insect control. Arch. Insect Biochem. Physiol. 1995, 30, 177–194. [Google Scholar] [CrossRef]
- Bonning, B.C.; Possee, R.D.; Hammock, B.D. Insecticidal efficacy of a recombinant baculovirus expressing JHE-KK, a modified juvenile hormone esterase. J. Invertebr. Pathol. 1999, 73, 234–236. [Google Scholar] [CrossRef] [PubMed]
- Eldridge, R.; O'Reilly, D.R.; Hammock, B.D.; Miller, L.K. Insecticidal properties of genetically engineered baculoviruses expressing an insect juvenile hormone esterase gene. Appl. Environ. Microbiol. 1992, 58, 1583–1591. [Google Scholar] [PubMed]
- Eldridge, R.; O’Reilly, D.R.; Miller, L.K. Efficacy of a baculovirus pesticide expressing an eclosion hormone gene. Biol. Control. 1992, 2, 104–110. [Google Scholar] [CrossRef]
- O'Reilly, D.R.; Kelly, T.J.; Masler, E.P.; Thyagaraja, B.S.; Robson, R.M.; Shaw, T.C.; Miller, L.K. Overexpression of Bombyx mori prothoracicotropic hormone using baculovirus vectors. Insect Biochem. Mol. Biol. 1995, 25, 475–485. [Google Scholar] [CrossRef] [PubMed]
- Ma, P.W.; Davis, T.R.; Wood, H.A.; Knipple, D.C.; Roelofs, W.L. Baculovirus expression of an insect gene that encodes multiple neuropeptides. Insect Biochem. Mol. Biol. 1998, 28, 239–249. [Google Scholar] [CrossRef] [PubMed]
- Raina, A.K.; Vakharia, V.N.; Leclerc, R.F.; Blackburn, M.B. Engineering a recombinant baculovirus with a peptide hormone gene and its effect on the corn earworm, Helicoverpa zea. Biopestic. Int. 2007, 3, 43–52. [Google Scholar]
- Hegedus, D.; Erlandson, M.; Gillott, C.; Toprak, U. New insights into peritrophic matrix synthesis, architecture, and function. Annu. Rev. Entomol. 2009, 54, 285–302. [Google Scholar] [CrossRef] [PubMed]
- Gopalakrishnan, K.; Muthukrishnan, S.; Kramer, K.J. Baculovirus-mediated expression of a Manduca sexta chitinase gene: Properties of the recombinant protein. Insect Biochem. Mol. Biol. 1995, 25, 255–265. [Google Scholar] [CrossRef]
- Li, H.; Tang, H.; Harrison, R.L.; Bonning, B.C. Impact of a basement membrane-degrading protease on dissemination and secondary infection of Autographa californica multiple nucleopolyhedrovirus in Heliothis virescens (Fabricus). J. Gen. Virol. 2007, 88, 1109–1119. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Li, L.; Moore, K.; Donly, C.; Theilmann, D.A.; Erlandson, M. Characterization of Mamestra configurata nucleopolyhedrovirus enhancin and its functional analysis via expression in an Autographa californica m nucleopolyhedrovirus recombinant. J. Gen. Virol. 2003, 84, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Del Rincón-Castro, M.C.; Ibarra, J.E. Effect of a nuclepolyhedrovirus of Autographa californica expressing the enhancin gene of Trichoplusia ni granulovirus on T. ni larvae. Biocontrol. Sci. Technol. 2005, 15, 701–710. [Google Scholar] [CrossRef]
- Detvisitsakun, C.; Cain, E.L.; Passarelli, A.L. The Autographa californica M nucleopolyhedrovirus fibroblast growth factor accelerates host mortality. Virology 2007, 365, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Hong-Lian, S.; Du-Juan, D.; Jin-Dong, H.; Jin-Xin, W.; Xiao-Fan, Z. Construction of the recombinant baculovirus AcMNPV with cathepsin B-like proteinase and its insecticidal activity against Helicoverpa armigera. Pestic. Biochem. Physiol. 2008, 91, 141–146. [Google Scholar] [CrossRef]
- Gramkow, A.W.; Perecmanis, S.; Sousa, R.L.; Noronha, E.F.; Felix, C.R.; Nagata, T.; Ribeiro, B.M. Insecticidal activity of two proteases against Spodoptera frugiperda larvae infected with recombinant baculoviruses. Virol. J. 2010, 7, 143. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, Y.; Corsaro, B.G.; Granados, R.R. Location and nucleotide sequence of the gene encoding the viral enhancing factor of the Trichoplusia ni granulosis virus. J. Gen. Virol. 1991, 72, 2645–2651. [Google Scholar] [CrossRef] [PubMed]
- Lepore, L.S.; Roelvink, P.R.; Granados, R.R. Enhancin, the granulosis virus protein that facilitates nucleopolyhedrovirus (NPV) infections, is a metalloprotease. J. Invertebr. Pathol. 1996, 68, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Granados, R.R. An intestinal mucin is the target substrate for a baculovirus enhancin. Proc. Natl. Acad. Sci. USA 1997, 94, 6977–6982. [Google Scholar] [CrossRef] [PubMed]
- Hynes, R.O. The evolution of metazoan extracellular matrix. J. Cell Biol. 2012, 196, 671–679. [Google Scholar] [CrossRef] [PubMed]
- Smith-Johannsen, H.; Witkiewicz, H.; Iatrou, K. Infection of silkmoth follicular cells with Bombyx mori nuclear polyhedrosis virus. J. Invert. Path. 1986, 48, 74–84. [Google Scholar] [CrossRef]
- Hess, R.T.; Falcon, L.A. Temporal events in the invasion of the codling moth, Cydia pomonella, by a granulosis virus. J. Invertebr. Pathol. 1987, 50, 85–105. [Google Scholar] [CrossRef]
- Tang, H.; Li, H.; Lei, S.M.; Harrison, R.L.; Bonning, B.C. Tissue specificity of a baculovirus-expressed, basement membrane-degrading protease in larvae of Heliothis virescens. Tissue Cell 2007, 39, 431–443. [Google Scholar] [CrossRef] [PubMed]
- Engelhard, E.K.; Kam-Morgan, L.N.; Washburn, J.O.; Volkman, L.E. The insect tracheal system: A conduit for the systemic spread of Autographa californica m nuclear polyhedrosis virus. Proc. Natl. Acad. Sci. USA 1994, 91, 3224–3227. [Google Scholar] [CrossRef] [PubMed]
- Passarelli, A.L. Barriers to success: How baculoviruses establish efficient systemic infections. Virology 2011, 411, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Means, J.C.; Passarelli, A.L. Viral fibroblast growth factor, matrix metalloproteases, and caspases are associated with enhancing systemic infection by baculoviruses. Proc. Natl. Acad. Sci. USA 2010, 107, 9825–9830. [Google Scholar] [CrossRef] [PubMed]
- Katsuma, S.; Horie, S.; Daimon, T.; Iwanaga, M.; Shimada, T. In vivo and in vitro analyses of a Bombyx mori nucleopolyhedrovirus mutant lacking functional vfgf. Virology 2006, 355, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Tang, H.; Sivakumar, S.; Philip, J.; Harrison, R.L.; Gatehouse, J.A.; Bonning, B.C. Insecticidal activity of a basement membrane-degrading protease against Heliothis virescens (Fabricius) and Acyrthosiphon pisum (Harris). J. Insect Physiol. 2008, 54, 777–789. [Google Scholar] [CrossRef] [PubMed]
- Merryweather, A.T.; Weyer, U.; Harris, M.P.; Hirst, M.; Booth, T.; Possee, R.D. Construction of genetically engineered baculovirus insecticides containing the Bacillus thuringiensis subsp. Kurstaki HD-73 delta endotoxin. J. Gen. Virol. 1990, 71, 1535–1544. [Google Scholar] [CrossRef] [PubMed]
- Martens, J.W.; Honee, G.; Zuidema, D.; van Lent, J.W.; Visser, B.; Vlak, J.M. Insecticidal activity of a bacterial crystal protein expressed by a recombinant baculovirus in insect cells. Appl. Environ. Microbiol. 1990, 56, 2764–2770. [Google Scholar] [PubMed]
- Lilley, M.; Ruffell, R.N.; Somerville, H.J. Purification of the insecticidal toxin in crystals of Bacillus thuringiensis. J. Gen. .Microbiol. 1980, 118, 1–11. [Google Scholar] [PubMed]
- Ribeiro, B.M.; Crook, N.E. Expression of full-length and truncated forms of crystal protein genes from Bacillus thuringiensis subsp. Kurstaki in a baculovirus and pathogenicity of the recombinant viruses. J. Invertebr. Pathol. 1993, 62, 121–130. [Google Scholar] [CrossRef] [PubMed]
- El-Menofy, W.; Osman, G.; Assaeedi, A.; Salama, M. A novel recombinant baculovirus overexpressing a Bacillus thuringiensis Cry1AB toxin enhances insecticidal activity. Biol. Proced. Online 2014, 16, 7. [Google Scholar] [CrossRef] [PubMed]
- Je, Y.H.; Jin, B.R.; Park, H.W.; Roh, J.Y.; Chang, J.H.; Seo, S.J.; Olszewski, J.A.; O’Reilly, D.R.; Kang, S.K. Baculovirus expression vectors that incorporate the foreign protein into viral occlusion bodies. Biotechniques 2003, 34, 81–87. [Google Scholar] [PubMed]
- Chang, J.H.; Choi, J.Y.; Jin, B.R.; Roh, J.Y.; Olszewski, J.A.; Seo, S.J.; O’Reilly, D.R.; Je, Y.H. An improved baculovirus insecticide producing occlusion bodies that contain Bacillus thuringiensis insect toxin. J. Invertebr. Pathol. 2003, 84, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Korth, K.L.; Levings, C.S.D. Baculovirus expression of the maize mitochondrial protein URF13 confers insecticidal activity in cell cultures and larvae. Proc. Natl. Acad. Sci. USA 1993, 90, 3388–3392. [Google Scholar] [CrossRef] [PubMed]
- Petrik, D.T.; Iseli, A.; Montelone, B.A.; van Etten, J.L.; Clem, R.J. Improving baculovirus resistance to UV inactivation: Increased virulence resulting from expression of a DNA repair enzyme. J. Invertebr. Pathol. 2003, 82, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Ross, L.; Guarino, L.A. Cycloheximide inhibition of delayed early gene expression in baculovirus-infected cells. Virology 1997, 232, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Passarelli, A.L.; Guarino, L.A. Baculovirus late and very late gene regulation. Curr. Drug Targets 2007, 8, 1103–1115. [Google Scholar] [CrossRef] [PubMed]
- Rohel, D.Z.; Cochran, M.A.; Faulkner, P. Characterization of two abundant mRNAs of Autographa. californica nuclear polyhedrosis virus present late in infection. Virology 1983, 124, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.E.; Vlak, J.M.; Summers, M.D. Physical analysis of Autographa californica nuclear polyhedrosis virus transcripts for polyhedrin and a 10,000-molecular-weight protein. J. Virol. 1983, 45, 215–225. [Google Scholar] [PubMed]
- Jarvis, D.L.; Summers, M.D. Glycosylation and secretion of human tissue plasminogen activator in recombinant baculovirus-infected insect cells. Mol. Cell Biol. 1989, 9, 214–223. [Google Scholar] [PubMed]
- Roelvink, P.W.; van Meer, M.M.; de Kort, C.A.; Possee, R.D.; Hammock, B.D.; Vlak, J.M. Dissimilar expression of Autographa californica multiple nucleocapsid nuclear polyhedrosis virus polyhedrin and p10 genes. J. Gen. Virol. 1992, 73, 1481–1489. [Google Scholar] [CrossRef] [PubMed]
- Bonning, B.C.; Roelvink, P.W.; Vlak, J.M.; Possee, R.D.; Hammock, B.D. Superior expression of juvenile hormone esterase and beta-galactosidase from the basic protein promoter of Autographa californica nuclear polyhedrosis virus compared to the p10 protein and polyhedrin promoters. J. Gen. Virol. 1994, 75, 1551–1556. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Z.; Ooi, B.G.; Miller, L.K. Baculovirus vectors for multiple gene expression and for occluded virus production. Gene 1991, 100, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Tessier, D.C.; Thomas, D.Y.; Khouri, H.E.; Laliberte, F.; Vernet, T. Enhanced secretion from insect cells of a foreign protein fused to the honeybee melittin signal peptide. Gene 1991, 98, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Perlak, F.J.; Fuchs, R.L.; Dean, D.A.; McPherson, S.L.; Fischhoff, D.A. Modification of the coding sequence enhances plant expression of insect control protein genes. Proc. Natl. Acad. Sci. USA 1991, 88, 3324–3328. [Google Scholar] [CrossRef] [PubMed]
- Ooi, B.G.; Miller, L.K. Regulation of host RNA levels during baculovirus infection. Virology 1988, 166, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.; Qiao, N.; Zhang, W.; Cheng, R.L.; Zhang, X.Q.; Bao, Y.Y.; Xu, Y.P.; Gu, L.Z.; Han, J.D.; Zhang, C.X. Dynamic interactions between Bombyx mori nucleopolyhedrovirus and its host cells revealed by transcriptome analysis. J. Virol. 2012, 86, 7345–7359. [Google Scholar] [CrossRef] [PubMed]
- Taniai, K.; Inceoglu, A.B.; Hammock, B.D. Expression efficiency of a scorpion neurotoxin, AahIT, using baculovirus in insect cells. Appl. Entomol. Zool. 2002, 37, 225–232. [Google Scholar] [CrossRef]
- Harrison, R.L.; Jarvis, D.L. Protein N-glycosylation in the baculovirus-insect cell expression system and engineering of insect cells to produce “mammalianized” recombinant glycoproteins. Adv. Virus Res. 2006, 68, 159–191. [Google Scholar] [PubMed]
- Fishman, L.; Kagan, M.L.; Zlotkin, E. Accessibility of the insect nervous system to a neurotoxic polypeptide. J. Exp. Zool. 1991, 257, 10–23. [Google Scholar] [CrossRef]
- Elazar, M.; Levi, R.; Zlotkin, E. Targeting of an expressed neurotoxin by its recombinant baculovirus. J. Exp. Biol. 2001, 204, 2637–2645. [Google Scholar] [PubMed]
- Hoover, K.; Schultz, C.M.; Lane, S.S.; Bonning, B.C.; Duffey, S.S.; McCutchen, B.F.; Hammock, B.D. Reduction in damage to cotton plants by a recombinant baculovirus that knocks moribund larvae of Heliothis virescens off the plant. Biol. Control. 1995, 5, 419–426. [Google Scholar] [CrossRef]
- Georgievska, L.; Joosten, N.; Hoover, K.; Cory, J.S.; Vlak, J.M.; van der Werf, W. Effects of single and mixed infections with wild type and genetically modified Helicoverpa armigera nucleopolyhedrovirus on movement behaviour of cotton bollworm larvae. Entomol. Exp. Appl. 2010, 135, 56–67. [Google Scholar] [CrossRef]
- D’Amico, V.; Elkinton, J.S. Rainfall effects on transmission of gypsy moth (Lepidoptera: Lymantriidae) nuclear polyhedrosis virus. Environ. Entomol. 1995, 24, 1144–1149. [Google Scholar] [CrossRef]
- Bonning, B.C.; Booth, T.F.; Hammock, B.D. Mechanistic studies of the degradation of juvenile hormone esterase in Manduca sexta. Arch. Insect Biochem. Physiol. 1997, 34, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Bonning, B.C.; Ward, V.K.; van Meer, M.M.; Booth, T.F.; Hammock, B.D. Disruption of lysosomal targeting is associated with insecticidal potency of juvenile hormone esterase. Proc. Natl. Acad. Sci. USA 1997, 94, 6007–6012. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.L.; Sun, X.C.; Bai, B.K.; van der Werf, W.; Vlak, J.M.; Hu, Z.H. Production of polyhedral inclusion bodies from Helicoverpa armigera larvae infected with wild-type and recombinant HaSNPV. Biocontrol. Sci. Technol. 2005, 15, 353–366. [Google Scholar] [CrossRef]
- Van Beek, N.; Davis, D.C. Baculovirus insecticide production in insect larvae. Meth. Mol. Biol. 2007, 388, 367–378. [Google Scholar]
- Lee, S.Y.; Qu, X.; Chen, W.; Poloumienko, A.; MacAfee, N.; Morin, B.; Lucarotti, C.; Krause, M. Insecticidal activity of a recombinant baculovirus containing an antisense c-myc fragment. J. Gen. Virol. 1997, 78, 273–281. [Google Scholar] [PubMed]
- Hajos, J.P.; Vermunt, A.M.; Zuidema, D.; Kulcsar, P.; Varjas, L.; de Kort, C.A.; Zavodszky, P.; Vlak, J.M. Dissecting insect development: Baculovirus-mediated gene silencing in insects. Insect Mol. Biol. 1999, 8, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Carthew, R.W.; Sontheimer, E.J. Origins and mechanisms of miRNAs and siRNAs. Cell 2009, 136, 642–655. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.P.; Chen, K.P.; Yao, Q.; Lin, M.H.; Gao, G.T.; Zhao, Y. Identification and characterization of an NPV infection-related gene bmsop2 in Bombyx mori l. J. Appl. Entomol. 2005, 129, 425–431. [Google Scholar] [CrossRef]
- Kang, L.; Shi, H.; Liu, X.; Zhang, C.; Yao, Q.; Wang, Y.; Chang, C.; Shi, J.; Cao, J.; Kong, J.; et al. Arginine kinase is highly expressed in a resistant strain of silkworm (Bombyx mori, lepidoptera): Implication of its role in resistance to Bombyx mori nucleopolyhedrovirus. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2011, 158, 230–234. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, Q.; Nielsen, L.K.; Reid, S. Genome scale transcriptomics of baculovirus-insect interactions. Viruses 2013, 5, 2721–2747. [Google Scholar] [CrossRef] [PubMed]
- Washburn, J.O.; Kirkpatrick, B.A.; Volkman, L.E. Insect protection against viruses. Nature 1996, 383, 767. [Google Scholar] [CrossRef]
- Washburn, J.O.; Haas-Stapleton, E.J.; Tan, F.F.; Beckage, N.E.; Volkman, L.E. Co-infection of Manduca sexta larvae with polydnavirus from Cotesia congregata increases susceptibility to fatal infection by Autographa californica M nucleopolyhedrovirus. J. Insect Physiol. 2000, 46, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Rivkin, H.; Kroemer, J.A.; Bronshtein, A.; Belausov, E.; Webb, B.A.; Chejanovsky, N. Response of immunocompetent and immunosuppressed Spodoptera littoralis larvae to baculovirus infection. J. Gen. Virol. 2006, 87, 2217–2225. [Google Scholar] [CrossRef] [PubMed]
- McNeil, J.; Cox-Foster, D.; Gardner, M.; Slavicek, J.; Thiem, S.; Hoover, K. Pathogenesis of Lymantria dispar multiple nucleopolyhedrovirus in L. dispar and mechanisms of developmental resistance. J. Gen. Virol. 2010, 91, 1590–1600. [Google Scholar] [CrossRef] [PubMed]
- Burand, J.P.; Hunter, W.B. RNAi: Future in insect management. J. Invertebr. Pathol. 2013, 112 Suppl., S68–S74. [Google Scholar] [PubMed]
- Terenius, O.; Papanicolaou, A.; Garbutt, J.S.; Eleftherianos, I.; Huvenne, H.; Kanginakudru, S.; Albrechtsen, M.; An, C.; Aymeric, J.L.; Barthel, A.; et al. RNA interference in lepidoptera: An overview of successful and unsuccessful studies and implications for experimental design. J. Insect Physiol. 2011, 57, 231–245. [Google Scholar] [CrossRef] [PubMed]
- Bautista, M.A.; Miyata, T.; Miura, K.; Tanaka, T. RNA interference-mediated knockdown of a cytochrome p450, cyp6bg1, from the diamondback moth, Plutella xylostella, reduces larval resistance to permethrin. Insect Biochem. Mol. Biol. 2009, 39, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Peng, H.; Yao, Q.; Chen, H.; Xie, Q.; Tang, B.; Zhang, W. Developmental control of a lepidopteran pest Spodoptera exigua by ingestion of bacteria expressing dsRNA of a non-midgut gene. PLoS One 2009, 4, e6225. [Google Scholar] [CrossRef] [PubMed]
- Di Lelio, I.; Varricchio, P.; Di Prisco, G.; Marinelli, A.; Lasco, V.; Caccia, S.; Casartelli, M.; Giordana, B.; Rao, R.; Gigliotti, S.; et al. Functional analysis of an immune gene of Spodoptera littoralis by RNAi. J. Insect Physiol. 2014, 64, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Yu, N.; Christiaens, O.; Liu, J.; Niu, J.; Cappelle, K.; Caccia, S.; Huvenne, H.; Smagghe, G. Delivery of dsRNA for RNAi in insects: An overview and future directions. Insect Sci. 2013, 20, 4–14. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kroemer, J.A.; Bonning, B.C.; Harrison, R.L. Expression, Delivery and Function of Insecticidal Proteins Expressed by Recombinant Baculoviruses. Viruses 2015, 7, 422-455. https://doi.org/10.3390/v7010422
Kroemer JA, Bonning BC, Harrison RL. Expression, Delivery and Function of Insecticidal Proteins Expressed by Recombinant Baculoviruses. Viruses. 2015; 7(1):422-455. https://doi.org/10.3390/v7010422
Chicago/Turabian StyleKroemer, Jeremy A., Bryony C. Bonning, and Robert L. Harrison. 2015. "Expression, Delivery and Function of Insecticidal Proteins Expressed by Recombinant Baculoviruses" Viruses 7, no. 1: 422-455. https://doi.org/10.3390/v7010422
APA StyleKroemer, J. A., Bonning, B. C., & Harrison, R. L. (2015). Expression, Delivery and Function of Insecticidal Proteins Expressed by Recombinant Baculoviruses. Viruses, 7(1), 422-455. https://doi.org/10.3390/v7010422