Seroprevalence Dynamics of European Bat Lyssavirus Type 1 in a Multispecies Bat Colony


Abstract
:1. Introduction
2. Experimental Materials and Methods
2.1. Study Area

2.2. Data Collection
2.3. Detection of EBLV-1 Neutralizing Antibodies
2.4. Statistical Analyses
3. Results
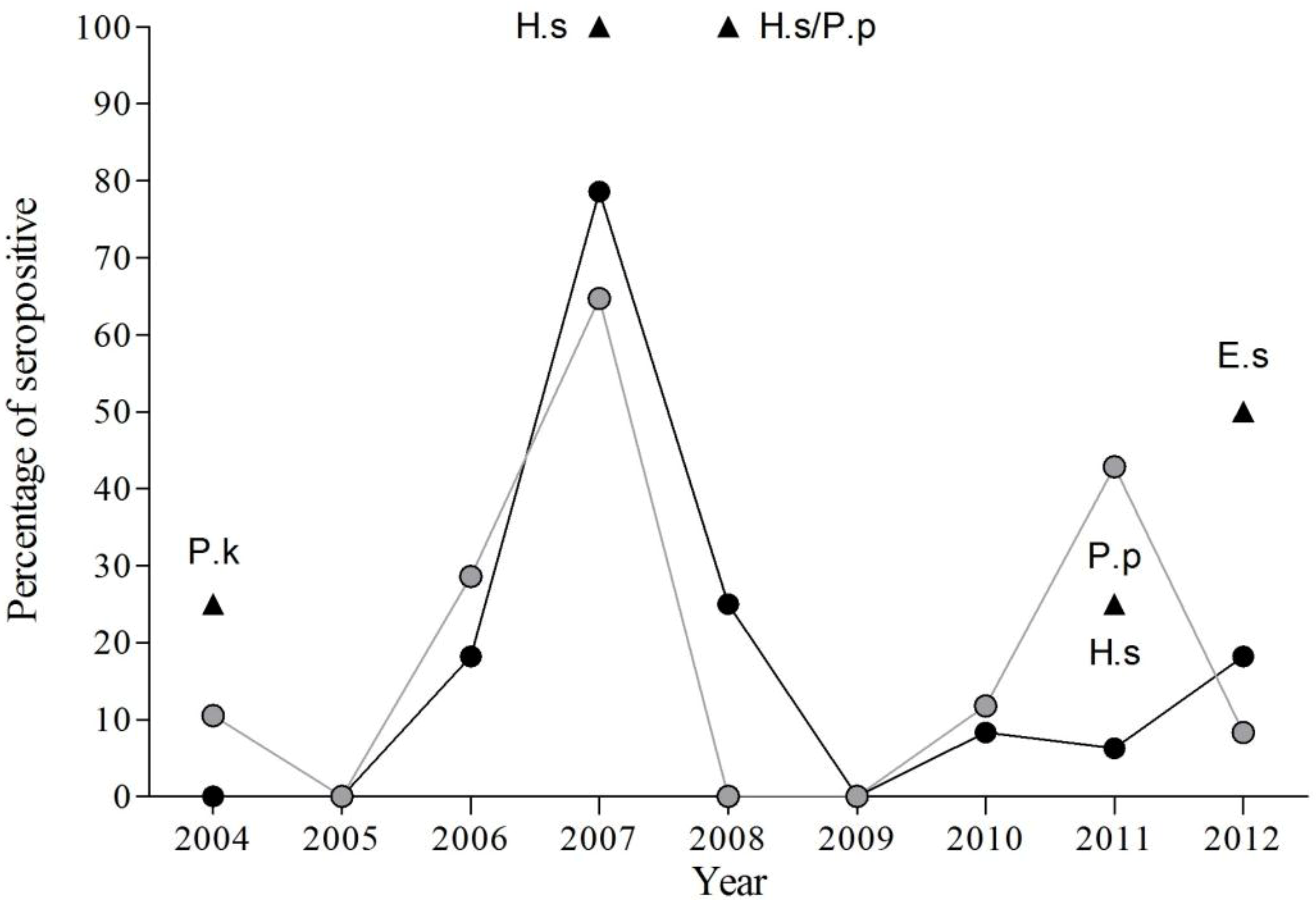
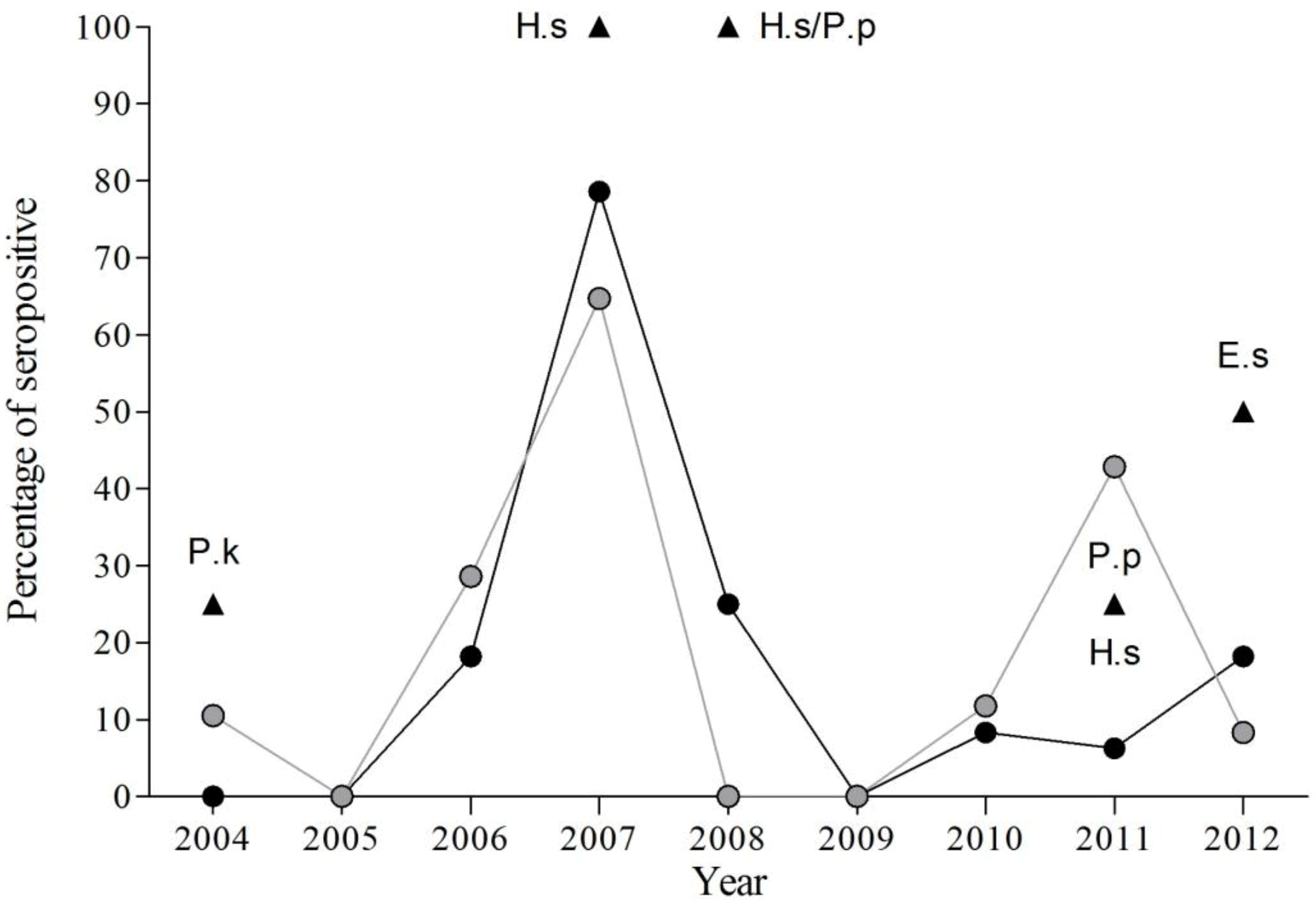
3.1. Presence of EBLV-1 Antibodies

{kind=link}
{kind=link}
{kind=link}
| Species | Females | Males | Total | ||||||
|---|---|---|---|---|---|---|---|---|---|
| n | n+ | % (95 CI) | n | n+ | % (95 CI) | n | n+ | % (95 CI) | |
| E. serotinus | nd | nd | nd | 9 | 1 | 11.11 (0.3–48.2) | 9 | 1 | 11.11 (0.3–48.2) |
| H. savii | 7 | 0 | 0 | 15 | 5 | 33.33 (11.8–61.6) | 22 | 5 | 22.73 (7.8–45.4) |
| M.daubentonii | 1 | 0 | 0 | 1 | 0 | 0 | 2 | 0 | 0 |
| M. myotis | nd | nd | nd | 1 | 0 | 0 | 1 | 0 | 0 |
| P. austriacus | 76 | 13 | 17.10 (9.4–27.5) | 56 | 8 | 14.28 (6.4–26.2) | 132 | 21 | 15.91 (10.1–23.3) |
| P. kuhlii | 6 | 1 | 16.67 (0.4–64.1) | 2 | 0 | 0 | 8 | 1 | 12.50 (0.3–52.6) |
| P. pipistrellus | 9 | 2 | 20.22 (2.8–60.0) | 19 | 2 | 10.53 (1.3 –33.1) | 28 | 4 | 14.28 (4.0–32.7) |
| P. pygmaeus | nd | nd | nd | 2 | 0 | 0 | 2 | 0 | 0 |
| T. teniotis | 94 | 23 | 24.47 (16.2–34.4) | 108 | 16 | 14.81 (8.7–22.9) | 202 | 39 | 19.31 (14.1–25.4) |
| Total | 193 | 39 | 20.21 (14.8–26.6) | 213 | 32 | 15.02 (10.5–20.5) | 406 | 71 | 17.49 (13.9–21.5) |
| Sex | Id | 2004 | 2005 | 2006 | 2007 | 2008 | 2009 | 2010 | 2011 | 2012 |
|---|---|---|---|---|---|---|---|---|---|---|
| Females | 1 | 0 | ns | ns | ns | ns | 0 | 0 | ns | 52 |
| 2 | ns | 0 | ns | ns | 56 | ns | 43 | ns | ns | |
| 3 | ns | 0 | ns | 56 | ns | ns | ns | ns | ns | |
| 4 | ns | ns | ns | ns | 35 | ns | ns | 0 | ns | |
| 5 | ns | ns | ns | 58 | 0 | ns | ns | ns | ns | |
| 6 | ns | ns | ns | 147 | 0 | ns | ns | ns | ns | |
| Males | 7 | ns | ns | 35 | 48 | ns | ns | ns | 0 | ns |
| 8 | ns | ns | ns | nd | 53 | ns | ns | ns | ns | |
| 9 | ns | ns | nd | 49 | ns | ns | ns | ns | ns |
3.2. Temporal Variation of EBLV-1 Antibodies
| Years | Females | Males | Total | ||||||
|---|---|---|---|---|---|---|---|---|---|
| n | n+ | % (95 CI) | n | n+ | % (95 CI) | n | n+ | % (95 CI) | |
| 2004 | 55 | 6 | 10.91 (4.1–22.2) | 55 | 1 | 1.82 (0.0–9.7) | 110 | 7 | 6.36 (2.6–12.7) |
| 2005 | 7 | 0 | 0 | 15 | 0 | 0 | 22 | 0 | 0 |
| 2006 | 30 | 9 | 30.00 (14.7–49.4) | 20 | 3 | 15.00 (3.2–37.9) | 50 | 12 | 24.00 (13.1–38.2) |
| 2007 | 18 | 14 | 77.78 (52.4–93.6) | 16 | 10 | 62.50 (35.4–84.8) | 34 | 24 | 70.59 (52.5–84.9) |
| 2008 | 13 | 3 | 23.08 (5.0–53.8) | 30 | 6 | 20.00 (7.7–38.6) | 43 | 9 | 20.93 (10.0–36.0) |
| 2009 | 17 | 0 | 0 | 24 | 0 | 0 | 41 | 0 | 0 |
| 2010 | 16 | 1 | 6.25 (0.1–30.2) | 14 | 2 | 14.29 (1.8–42.8) | 30 | 3 | 10.00 (2.1–26.5) |
| 2011 | 26 | 4 | 15.38 (4.4–34.9) | 23 | 8 | 34.78 (16.4–57.3) | 49 | 12 | 24.49 (13.3–38.9) |
| 2012 | 11 | 2 | 18.18 (2.3–51.8) | 16 | 2 | 12.50 (1.5–38.3) | 27 | 4 | 14.81 (4.2–33.7) |
| Years | E. serotinus | P. kuhlii | H. savii | ||||||
|---|---|---|---|---|---|---|---|---|---|
| n | n+ | % (95 CI) | n | n+ | % (95 CI) | n | n+ | % (95 CI) | |
| 2004 | 5 | 0 | 0 | 4 | 1 | 25.00 (0.6–80.6) | 3 | 0 | 0 |
| 2005 | nd | nd | nd | 1 | 0 | 0 | 1 | 0 | 0 |
| 2006 | 1 | 0 | 0 | nd | nd | nd | 1 | 0 | 0 |
| 2007 | nd | nd | nd | nd | nd | nd | 2 | 2 | 100.00 (22.4–100.0) |
| 2008 | nd | nd | nd | nd | nd | nd | 2 | 2 | 100.00 (22.4–100.0) |
| 2009 | nd | nd | nd | nd | nd | nd | 9 | 0 | 0 |
| 2010 | 1 | 0 | 0 | nd | nd | nd | nd | nd | nd |
| 2011 | nd | nd | nd | 3 | 0 | 0 | 4 | 1 | 25.00 (0.6–80.6) |
| 2012 | 2 | 1 | 50.00 (1.3–98.7) | nd | nd | nd | nd | nd | nd |
| Years | P. pipistrellus | P. austriacus | T. teniotis | ||||||
|---|---|---|---|---|---|---|---|---|---|
| n | n+ | % (95 CI) | n | n+ | % (95 CI) | n | n+ | % (95 CI) | |
| 2004 | 7 | 0 | 0 | 34 | 0 | 0 | 57 | 6 | 10.53 (4.0–21.5) |
| 2005 | 2 | 0 | 0 | 13 | 0 | 0 | 5 | 0 | 0 |
| 2006 | 2 | 0 | 0 | 11 | 2 | 18.18 (2.3–51.8) | 35 | 10 | 28.57 (14.6–46.3) |
| 2007 | 1 | 0 | 0 | 14 | 11 | 78.57 (49.2–95.3) | 17 | 11 | 64.71 (38.3–85.8) |
| 2008 | 3 | 3 | 100.00 (36.8–100.0) | 16 | 4 | 25.00 (7.3–52.4) | 22 | 0 | 0 |
| 2009 | 7 | 0 | 0 | 5 | 0 | 0 | 16 | 0 | 0 |
| 2010 | nd | nd | nd | 12 | 1 | 8.33 (0.2–38.5) | 17 | 2 | 11.76 (1.5–36.4) |
| 2011 | 4 | 1 | 25.00 (0.6–80.6) | 16 | 1 | 6.25 (0.2–30.2) | 21 | 9 | 42.86 (21.8–66.0) |
| 2012 | 2 | 0 | 0 | 11 | 2 | 18.18 (2.3–51.8) | 12 | 1 | 8.33 (0.2–38.5) |
| GAM model expression | AICc | ΔAICc |
|---|---|---|
| 1- seroprevalence ~ s(year,by = P. austriacus) + s(year,by = T. teniotis) | 273.22 | 0.00 |
| 2- seroprevalence ~ sex + s(year) | 286.49 | 13.27 |
| 3- seroprevalence ~ s(year) | 286.90 | 13.68 |
| 4- seroprevalence ~ sex + species + s(year) | 287.75 | 14.53 |
| 5- seroprevalence ~ species + s(year) | 288.39 | 15.17 |
| 6- seroprevalence ~ sex × species + s(year) | 289.53 | 16.31 |
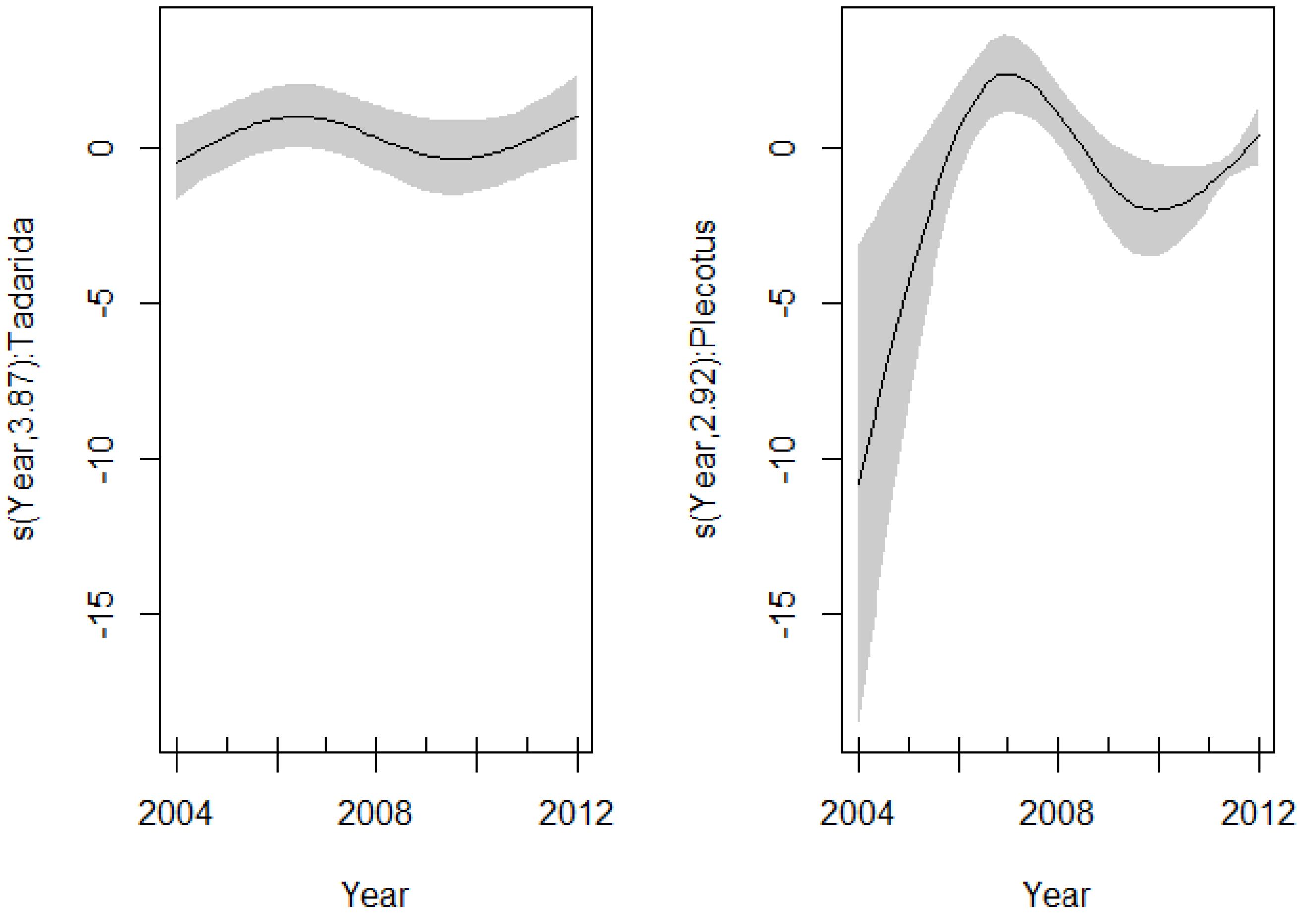
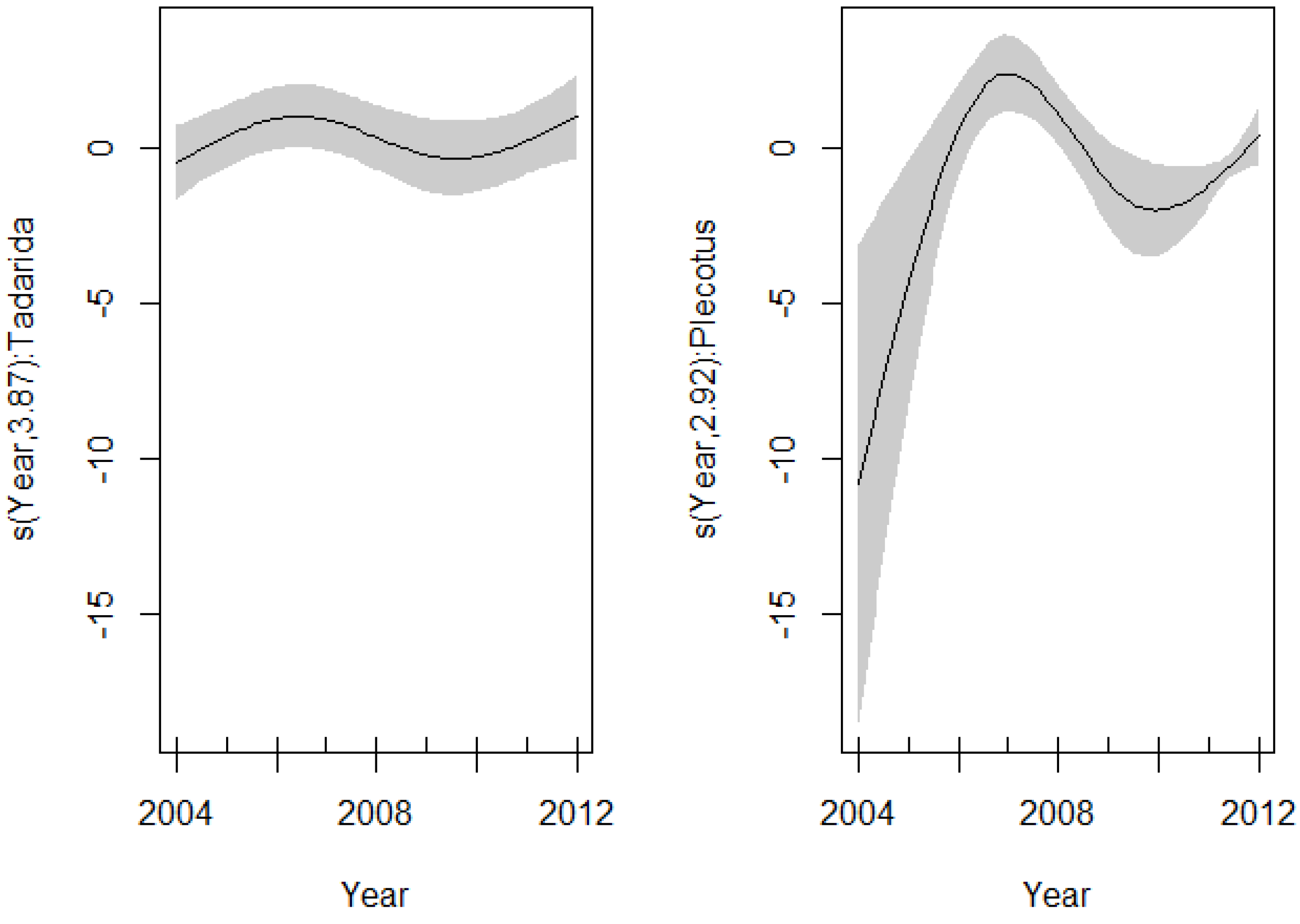
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References and Notes
- Daszak, P.; Cunninham, A.A.; Hyatt, A.D. Emerging infectious diseases of wildlife — Threats of biodiversity and human health. Science 2000, 287, 443–449. [Google Scholar] [CrossRef]
- Karesh, W.B.; Dobson, A.; Lloyd-Smith, J.O.; Lubroth, J.; Dixon, M.A.; Bennett, M.; Aldrich, S.; Harrington, T.; Formenty, P.; Loh, E.H.; et al. Ecology of zoonoses: natural and unnatural histories. Lancet 2012, 380, 1936–1945. [Google Scholar]
- Calisher, C.H.; Childs, J.E.; Field, H.E.; Holmes, K.V.; Schountz, T. Bats: Important reservoir hosts of emerging viruses. Clin. Microbiol. Rev. 2006, 19, 531–545. [Google Scholar] [CrossRef]
- Wong, S.; Lau, S.; Woo, P.; Yuen, K.Y. Bats as a continuing source of emerging infections in humans. Rev. Med. Virol. 2007, 17, 67–91. [Google Scholar] [CrossRef]
- Dacheux, L.; Cervantes-Gonzalez, M.; Guigon, G.; Thiberge, J.-M.; Vandenbogaert, M.; Moufrais, C.; Caro, V.; Bourhy, H. A Preliminary Study of Viral Metagenomics of French Bat Species in Contact with Humans: Identification of New Mammalian Viruses. PLoS One 2014, 9, e87194. [Google Scholar]
- Brosset, A. The migrations of Pipistrellus nathusii in France—Possible implication on the spreading of rabies (in French). Mammal 1990, 54, 207–212. [Google Scholar]
- Halloran, M.E. Concepts of infectious disease epidemiology. In Modern epidemiology; Rothman, K.J., Greenland, S., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 1998; pp. 529–554. [Google Scholar]
- George, D.B.; Webb, C.T.; Farnsworth, M.L.; O’Shea, T.J.; Bowen, R.A.; Smith, D.L.; Stanley, T.R.; Ellison, L.E.; Rupprecht, C.E. Host and viral ecology determine bat rabies seasonality and maintenance. Proc. Natl. Acad. Sci. USA 2011, 108, 10208–10213. [Google Scholar] [CrossRef]
- Hayman, D.T.S.; Bowen, R.A.; Cryan, P.M.; McCracken, G.F.; O’Shea, T.J.; Peel, A.J.; Gilbert, A.; Webb, C.T.; Wood, J.L.N. Ecology of Zoonotic Infectious Diseases in Bats: Current Knowledge and Future Directions. Zoon. Public Health 2013, 14, 2–21. [Google Scholar]
- Kuzmin, I.V.; Bozick, B.; Guagliardo, S.A.; Kunkel, R.; Shak, J.R.; Tong, S.; Rupprecht, C.E. Bats, emerging infectious diseases, and the rabies paradigm revisited. Emerg. Health Threats J. 2011, 4, 7159–7176. [Google Scholar]
- Kuzmin, I.V.; Mayer, A.E.; Niezgoda, M.; Markotter, W.; Agwanda, B.; Breiman, R.F.; Rupprecht, C.E. Shimoni bat virus, a new representative of the Lyssavirus genus. Virus Res. 2010, 149, 197–210. [Google Scholar] [CrossRef]
- Freuling, C.M.; Beer, M.; Conraths, F.J.; Finke, S.; Hoffmann, B.; Keller, B.; Kliemt, J.; Mettenleiter, T.C.; Mühlbach, E.; Teifke, J.P.; et al. Novel Lyssavirus in Natterer’s Bat, Germany. Emerg. Infect. Dis. 2011, 17, 1519–1522. [Google Scholar]
- Marston, D.A.; Horton, D.L.; Ngeleja, C.; Hampson, K.; McElhinney, L.M.; Banyard, A.C.; Haydon, D.; Cleaveland, S.; Rupprecht, C.E.; Bigambo, M.; et al. Ikoma Lyssavirus, Highly Divergent Novel Lyssavirus in an African Civet. Emerg. Infect. Dis. 2012, 18, 664–667. [Google Scholar]
- Badrane, H.; Tordo, N. Host switching in Lyssavirus history from the Chiroptera to the Carnivora orders. J. Virol. 2001, 75, 8096–8104. [Google Scholar]
- Delmas, O.; Holmes, E.C.; Talbi, C.; Larrous, F.; Dacheux, L.; Bouchier, C.; Bourhy, H. Genomic diversity and evolution of the lyssaviruses. PLoS One 2008, 3, e2057. [Google Scholar]
- Ceballos, N.A.; Vázquez-Morón, S.; Berciano, J.M.; Nicolás, O.; Aznar-López, C.; Juste, J.; Rodríguez-Nevado, C.; Aguilar-Setién, A.; Echevarría, J. Novel Lyssavirus in bat, Spain. Emerg. Infect. Dis. 2013, 19, 793–795. [Google Scholar] [CrossRef]
- Kuzmin, I. V.; Hughes, G.J.; Botvinkin, A.D.; Orciari, L.A.; Rupprecht, C.E. Phylogenetic relationships of Irkut and West Caucasian bat viruses within the Lyssavirus genus and suggested quantitative criteria based on the N gene sequence for lyssavirus genotype definition. Virus Res. 2005, 111, 28–43. [Google Scholar] [CrossRef]
- Davis, P.; Holmes, E.C.; Larrous, F.; Van der Poel, W.H.M.; Tjørnehøj, K.; Alonso, W.J.; Bourhy, H. Phylogeography, population dynamics, and molecular evolution of European bat lyssaviruses. J. Virol. 2005, 79, 10487–10497. [Google Scholar] [CrossRef]
- Turmelle, A.S.; Allen, L.C.; Jackson, F.R.; Kunz, T.H.; Rupprecht, C.E.; McCracken, G.F. Ecology of rabies virus exposure in colonies of Brazilian free-tailed bats (Tadarida brasiliensis) at natural and man-made roosts in Texas. Vector Borne Zoonotic Dis. 2010, 10, 165–175. [Google Scholar] [CrossRef]
- Drexler, J.F.; Corman, V.M.; Wegner, T.; Tateno, A.F.; Zerbinati, R.M.; Gloza-Rausch, F.; Seebens, A.; Muller, M.A.; Drosten, C. Amplification of emerging viruses in a bat colony. Emerg. Infect. Dis. 2011, 17, 449–456. [Google Scholar]
- Plowright, R.K.; Foley, P.; Field, H.E.; Dobson, A.P.; Foley, J.E.; Eby, P.; Daszak, P. Urban habituation, ecological connectivity and epidemic dampening: the emergence of Hendra virus from flying foxes (Pteropus spp.). Proc. R. Soc. B. 2011, 278, 3703–3712. [Google Scholar] [CrossRef]
- Serra-Cobo, J.; López-Roig, M.; Seguí, M.; Sánchez, L.P.; Nadal, J.; Borràs, M.; Lavenir, R.; Bourhy, H. Ecological Factors Associated with European Bat Lyssavirus Seroprevalence in Spanish Bats. PLoS One 2013, 8, e64467. [Google Scholar]
- Serra-Cobo, J.; Amengual, B.; Abellán, C.; Bourhy, H. European bat Lyssavirus infection in Spanish bat populations. Emerg. Infect. Dis. 2002, 8, 413–420. [Google Scholar] [CrossRef]
- Amengual, B.; Bourhy, H.; López-Roig, M.; Serra-Cobo, J. Temporal dynamics of European bat Lyssavirus type 1 and survival of Myotis myotis bats in natural colonies. PLoS One 2007, 2, e566. [Google Scholar] [CrossRef]
- Serra-Cobo, J.; Barbault, R.; Estada-Peña, A. Le gouffre de San Pedro de los Griegos (Oliete, Teruel, Espagne): un refuge de biodiversité sans équivalent en Europe (in French). Rev. Ecol. (Terre et Vie) 1993, 48, 341–348. [Google Scholar]
- Dietz, C.; Von Helversen, O. Illustrated identification key to the bats of Europe. Available online: http://biocenosi.dipbsf.uninsubria.it/didattica/bat_key1.pdf (accessed on 14 March 2014).
- Racey, P.A. Ageing and assessment of reproductive status of pipistrelle bats, Pipistrellus pipistrellus. J. Zool. 1974, 173, 264–271. [Google Scholar] [CrossRef]
- Bourhy, H.; Sureau, P. Rapid fluorescent focus inhibitions test (RFFIT). In Commission des Laboratoires de Référence et d’Expertise, editors. Méthodes de Laboratoire pour le Diagnostique de la Rage (in French); Institut Pasteur: Paris, France, 1990; pp. 191–193. [Google Scholar]
- Lumlertdacha, B.; Boongird, K.; Wanghongsa, S.; Wacharapluesadee, S.; Chanhome, L.; Khawplod, P.; Hemachudha, T.; Kuzmin, I.; Rupprecht, C.E. Survey for bat Lyssavirus, Thailand. Emerg. Infect. Dis. 2005, 11, 232–236. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model selection and multimodal inference: A practical information–theoretic approach; Springer: New York, NY, USA, 2002; p. 496. [Google Scholar]
- R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Available online: http://www.r-project.org/ (accessed on 14 March 2014).
- López-Roig, M.; Abellán, C.; Amengual, B.; Sánchez-Serrano, L.; Martínez, B.; Lapuente, J.; Borrás, M.; Mas, O.; Serra-Cobo, J. Análisis demográficos y sanitarios en las colonias de Plecotus austriacus y Tadarida teniotis de la sima de San Pedro (Oliete, Parque Cultural del Río Martín) (in Spanish). Cauce 2007, 27, 28–30. [Google Scholar]
- Picard-Meyer, E.; Dubourg-Savage, M.J.; Arthur, L.; Barataud, M.; Bécu, D.; Bracco, S.; Borel, C.; Larcher, G.; Meme-Lafond, B.; Moinet, M.; et al. Active surveillance of bat rabies in France: A 5-year study (2004–2009). Vet. Microbiol. 2011, 151, 390–395. [Google Scholar] [CrossRef]
- Plowright, R.K.; Field, H.E.; Smith, C.; Divljan, A.; Palmer, C.; Tabor, G.M.; Daszak, P.; Foley, J.E. Reproduction and nutritional stress are risk factors for Hendra virus infection in little red flying foxes (Pteropus scapulatus). Proc. R. Soc. B. 2008, 275, 861–869. [Google Scholar] [CrossRef]
- Lloyd, S.S. Immunosuppression during pregnancy and lactation. Ir. Vet. J. 1983, 37, 64–70. [Google Scholar]
- Pons-Salort, M.; Serra-Cobo, J.; Jay, F.; López-Roig, M.; Lavenir, R.; Guillemot, D.; Letort, V.; Bourhy, H.; Opatowski, L. Insights into persistence mechanisms of a zoonotic virus in bat colonies using a multispecies metapopulation model. PLoS One 2014, 9, e95610. [Google Scholar]
- Hutterer, R.; Ivanova, T.; Meyer-Cords, C.; Rodrigues, L. Bat migrations in Europe. A review of banding data and literature; Federal Agency for Nature Conservation: Bonn, Germany, 2005; p. 162. [Google Scholar]
- Altizer, S.; Bartel, R.; Han, B.A. Animal migration and infectious disease risk. Science 2011, 331, 296–302. [Google Scholar] [CrossRef]
- Schatz, J.; Fooks, A.R.; McElhinney, L.; Horton, D.; Echevarria, J.; Vazquez- Moron, S.; Kooi, E.A.; Rasmussen, T.B.; Müller, T.; Freuling, C.M. Bat rabies surveillance in Europe. Zoonoses Public Health. 2013, 60, 22–34. [Google Scholar] [CrossRef]
- Schatz, J.; Ohlendorf, B.; Busse, P.; Pelz, G.; Dolch, D.; Teubner, J.; Encarnacão, J.A.; Mühle, R.U.; Fischer, M.; Hoffmann, B.; et al. Twenty years of active bat rabies surveillance in Germany: A detailed analysis and future perspectives. Epidemiol. Infect. 2013, 142, 1155–1166. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
López-Roig, M.; Bourhy, H.; Lavenir, R.; Serra-Cobo, J. Seroprevalence Dynamics of European Bat Lyssavirus Type 1 in a Multispecies Bat Colony. Viruses 2014, 6, 3386-3399. https://doi.org/10.3390/v6093386
López-Roig M, Bourhy H, Lavenir R, Serra-Cobo J. Seroprevalence Dynamics of European Bat Lyssavirus Type 1 in a Multispecies Bat Colony. Viruses. 2014; 6(9):3386-3399. https://doi.org/10.3390/v6093386
Chicago/Turabian StyleLópez-Roig, Marc, Hervé Bourhy, Rachel Lavenir, and Jordi Serra-Cobo. 2014. "Seroprevalence Dynamics of European Bat Lyssavirus Type 1 in a Multispecies Bat Colony" Viruses 6, no. 9: 3386-3399. https://doi.org/10.3390/v6093386
APA StyleLópez-Roig, M., Bourhy, H., Lavenir, R., & Serra-Cobo, J. (2014). Seroprevalence Dynamics of European Bat Lyssavirus Type 1 in a Multispecies Bat Colony. Viruses, 6(9), 3386-3399. https://doi.org/10.3390/v6093386