Recent Observations on Australian Bat Lyssavirus Tropism and Viral Entry


Abstract
:1. Discovery of Australian Bat Lyssavirus
2. Host Reservoirs and Geographic Distribution
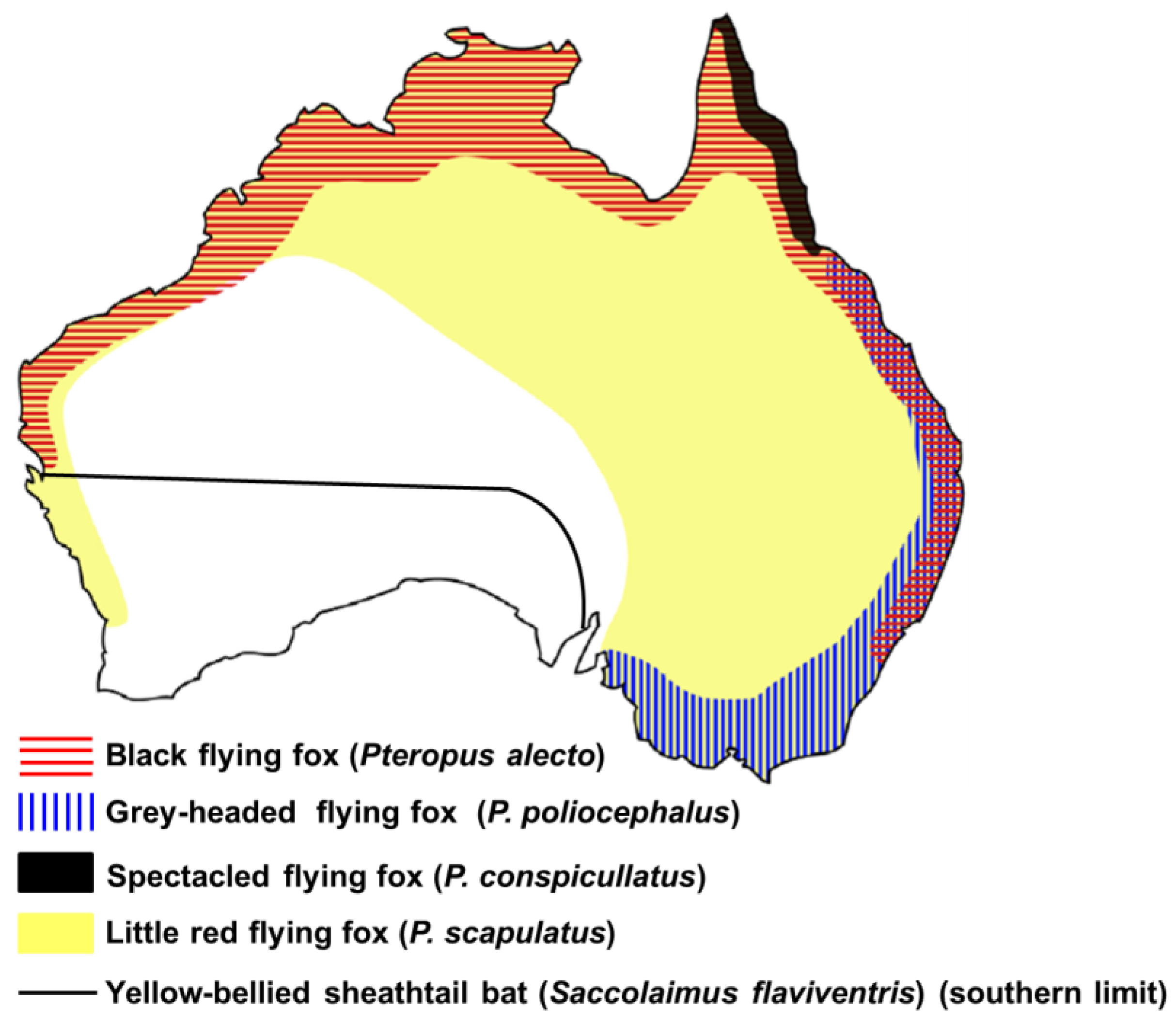
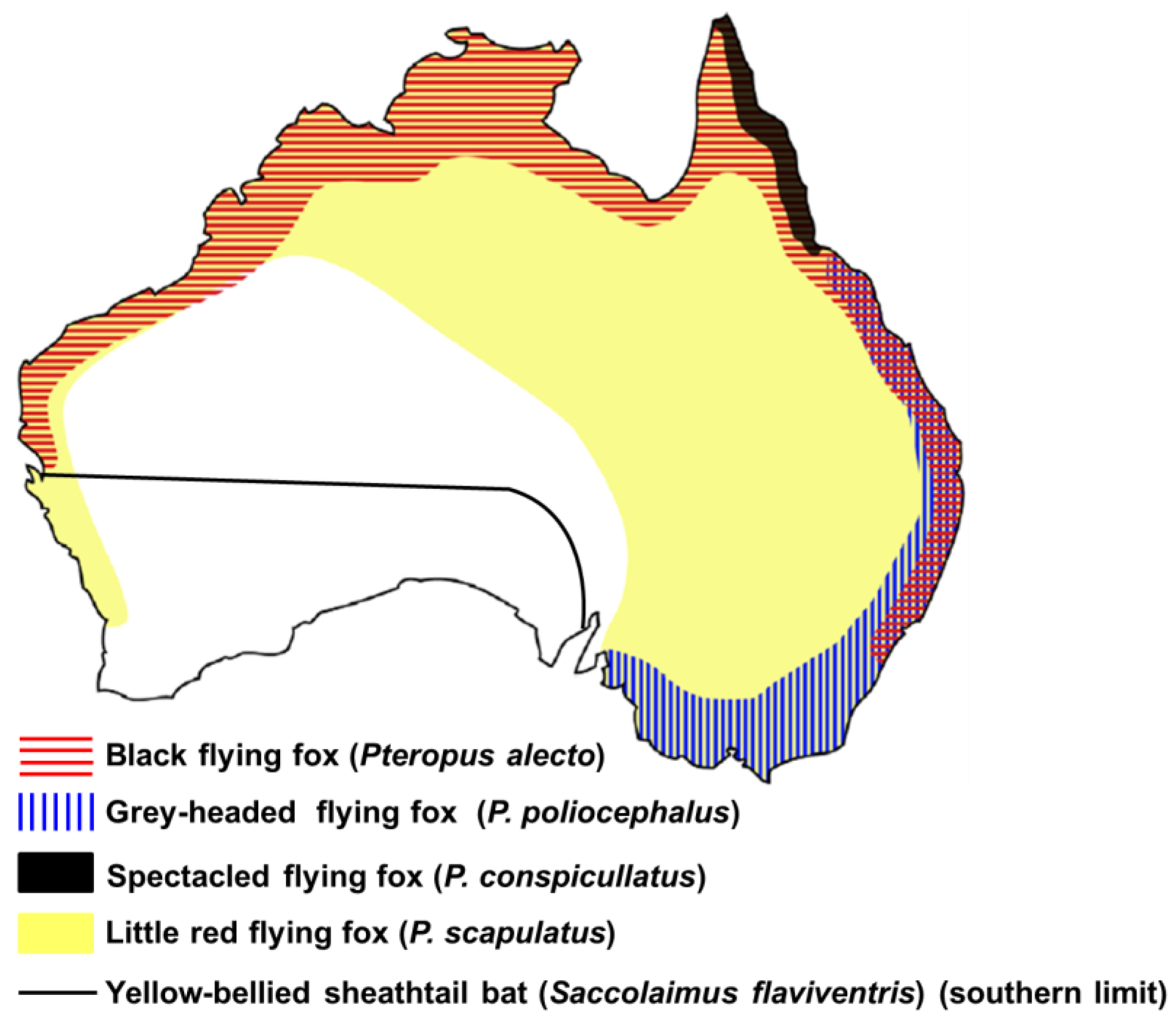
2.1 Prevalence of ABLV
3. Susceptible Species
3.1. Flying Foxes
3.2. Humans
3.3. Horses
3.4. Could Other Terrestrial Mammals be at Risk for ABLV Infection?
4. Molecular Biology
4.1. Taxonomy

{kind=link}
{kind=link}
| Lyssaviruses | Genotype | Phylogroup |
|---|---|---|
| * Rabies virus (RABV) | 1 | I |
| Lagos Bat virus (LBV) | 2 | II |
| * Mokola virus (MOKV) | 3 | II |
| * Duvenhage virus (DUVV) | 4 | I |
| * European bat lyssavirus 1 (EBLV1) | 5 | I |
| * European bat lyssavirus 2 (EBLV2) | 6 | I |
| * Australian bat lyssavirus (ABLV) | 7 | I |
| Aravan virus (ARAV) | 8 | I |
| Khujand virus (KHUV) | 9 | I |
| * Irkut virus (IRKV) | 10 | I |
| West Caucasian bat virus (WCBV) | 11 | III |
| Shimoni bat virus (SHIBV) | 12 | III |
| Bokeloh bat lyssavirus (BBLV) | NC | I |
| Ikoma lyssavirus (IKOV) | NC | III |
| Lleida bat lyssavirus (LLEBV) | NC | III |
4.2. Lyssavirus Virion Structure
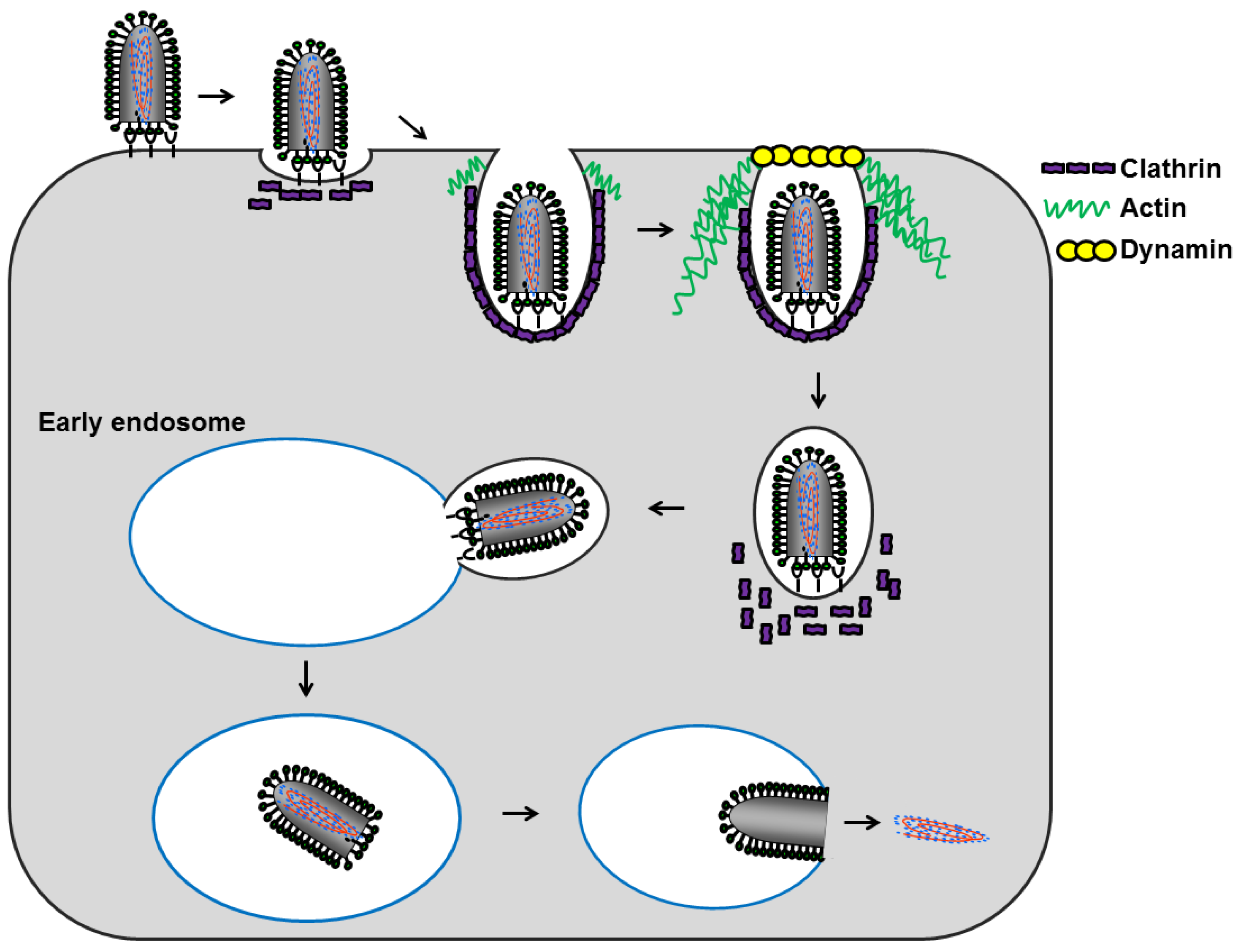
5. Lyssavirus Entry
5.1. Attachment to Host Cells
5.2. Internalization
5.3. Uncoating
5.4. Model for Lyssavirus Entry
6. Conclusions
Supplementary Files
Acknowledgments
Author Contributions
Conflicts of Interest
References and Notes
- Murray, K.; Selleck, P.; Hooper, P.; Hyatt, A.; Gould, A.; Gleeson, L.; Westbury, H.; Hiley, L.; Selvey, L.; Rodwell, B.; et al. A morbillivirus that caused fatal disease in horses and humans. Science 1995, 268, 94–97. [Google Scholar]
- Young, P.L.; Halpin, K.; Selleck, P.W.; Field, H.; Gravel, J.L.; Kelly, M.A.; Mackenzie, J.S. Serologic evidence for the presence in Pteropus bats of a paramyxovirus related to equine morbillivirus. Emerg. Infect. Dis. 1996, 2, 239–240. [Google Scholar] [CrossRef]
- Halpin, K.; Young, P.L.; Field, H.E.; Mackenzie, J.S. Isolation of Hendra virus from pteropid bats: A natural reservoir of Hendra virus. J. Gen. Virol. 2000, 81, 1927–1932. [Google Scholar]
- Fraser, G.C.; Hooper, P.T.; Lunt, R.A.; Gould, A.R.; Gleeson, L.J.; Hyatt, A.D.; Russell, G.M.; Kattenbelt, J.A. Encephalitis caused by a Lyssavirus in fruit bats in Australia. Emerg. Infect. Dis. 1996, 2, 327–331. [Google Scholar] [CrossRef]
- Gould, A.R.; Hyatt, A.D.; Lunt, R.; Kattenbelt, J.A.; Hengstberger, S.; Blacksell, S.D. Characterisation of a novel lyssavirus isolated from Pteropid bats in Australia. Virus Res. 1998, 54, 165–187. [Google Scholar] [CrossRef]
- Hooper, P.T.; Lunt, R.A.; Gould, A.R.; Samaratunga, H.; Hyatt, A.D.; Gleeson, L.J.; Rodwell, B.J.; Rupprecht, C.E.; Smith, J.S.; Murray, P.K. A new lyssavirus: The first endemic rabies-related virus recognized in Australia. Bull. Inst. Pasteur. 1997, 95, 209–218. [Google Scholar] [CrossRef]
- Gould, A.R.; Kattenbelt, J.A.; Gumley, S.G.; Lunt, R.A. Characterisation of an Australian bat lyssavirus variant isolated from an insectivorous bat. Virus Res. 2002, 89, 1–28. [Google Scholar] [CrossRef]
- Wilson, D.E.; Reeder, D.M. Mammal Species of the World. A Taxonomic and Geographic Reference, 3rd ed.; Johns Hopkins University Press: Baltimore, MD, USA, 2005; p. 2000. [Google Scholar]
- Hall, L.; Richards, G. Flying Foxes: Fruit and Blossom Bats of Australia; University of New South Wales Press: Sydney, Australia, 2000; p. 160. [Google Scholar]
- McCall, B.J.; Epstein, J.H.; Neill, A.S.; Heel, K.; Field, H.; Barrett, J.; Smith, G.A.; Selvey, L.A.; Rodwell, B.; Lunt, R. Potential exposure to Australian bat lyssavirus, Queensland, 1996–1999. Emerg. Infect. Dis. 2000, 6, 259–264. [Google Scholar] [CrossRef]
- Arguin, P.M.; Murray-Lillibridge, K.; Miranda, M.E.; Smith, J.S.; Calaor, A.B.; Rupprecht, C.E. Serologic evidence of Lyssavirus infections among bats, the Philippines. Emerg. Infect. Dis. 2002, 8, 258–262. [Google Scholar] [CrossRef]
- Van Dyke, S.; Strahan, R. The Mammals of Australia, 3rd ed.; New Holland/Queensland Museum: Brisbane, Australia, 2008; p. 888. [Google Scholar]
- Animal Health Australia. Response Policy Brief: Hendra Virus Infection (Version 3.5). In Australian Veterinary Emergency Plan (AUSVETPLAN), 3rd ed.; Standing Council on Primary Industries: Canberra, Australia, 2013. [Google Scholar]
- Richards, G.; Hall, L. A Natural History of Australian Bats: Working the Night Shift; CSIRO Publishing: Collingwood, Australia, 2012; p. 183. [Google Scholar]
- Field, H.E. Australian Bat Lyssavirus. Ph.D. Thesis, University of Queensland, Queensland, Australia, 2005. [Google Scholar]
- Barrett, J. Australian Bat Lyssavirus. Ph.D. Thesis, University of Queensland, Queensland, Australia, 2004. [Google Scholar]
- Field, H.; McCall, B.; Barrett, J. Australian bat lyssavirus infection in a captive juvenile black flying fox. Emerg. Infect. Dis. 1999, 5, 438–440. [Google Scholar] [CrossRef]
- Warrilow, D.; Harrower, B.; Smith, I.L.; Field, H.; Taylor, R.; Walker, C.; Smith, G.A. Public health surveillance for Australian bat lyssavirus in Queensland, Australia, 2000–2001. Emerg. Infect. Dis. 2003, 9, 262–264. [Google Scholar] [CrossRef]
- Allworth, A.; Murray, K.; Morgan, J. A human case of encephalitis due to a lyssavirus recently identified in fruit bats. Commun. Dis. Intellig. 1996, 20, 504. [Google Scholar]
- Samaratunga, H.; Searle, J.W.; Hudson, N. Non-rabies Lyssavirus human encephalitis from fruit bats: Australian bat Lyssavirus (pteropid Lyssavirus) infection. Neuropathol. Appl. Neurobiol. 1998, 24, 331–335. [Google Scholar] [CrossRef]
- Hanna, J.N.; Carney, I.K.; Smith, G.A.; Tannenberg, A.E.; Deverill, J.E.; Botha, J.A.; Serafin, I.L.; Harrower, B.J.; Fitzpatrick, P.F.; Searle, J.W. Australian bat lyssavirus infection: A second human case, with a long incubation period. Med. J. Aust. 2000, 172, 597–599. [Google Scholar]
- Australian bat lyssavirus—Australia (02). Available online: http://www.promedmail.org/direct.php?id=20130323.1600266 (accessed on 23 March 2013).
- Hayes, L. Australian Rabies. Available online: http://sixtyminutes.ninemsn.com.au/article.aspx?id=8667746 (accessed on 31 May 2013).
- Arthur, R.; Human, F.; Williamson, G.; Dickason, C.; Conway, M.L.; Bell, C.; Finkelstein, J. State and territory reports. In Animal Health Surveillance Quarterly Report; Animal Health Australia: Deakin, Australia, 2013; Volume 18, pp. 11–22. [Google Scholar]
- Roth, I. CVO Bulletin to NSW Veterinarians regarding Australian Bat Lyssavirus (ABLV); Bulletin Number 20130529; Veterinary Practitioners Board of New South Wales: Mascot, Australia, 29 May; 2013. [Google Scholar]
- Annand, E.A.; Shinwari, W.; Randwick Equine Centre, Sydney, Australia. Unpublished work. 2014.
- McCall, B.J.; Field, H.E.; Smith, G.A.; Storie, G.J.; Harrower, B.J. Defining the risk of human exposure to Australian bat lyssavirus through potential non-bat animal infection. Commun. Dis. Intell. 2005, 29, 202–205. [Google Scholar]
- Banyard, A.C.; Hayman, D.; Johnson, N.; McElhinney, L.; Fooks, A.R. Bats and lyssaviruses. Adv. Virus Res. 2011, 79, 239–289. [Google Scholar] [CrossRef]
- Thoulouze, M.I.; Lafage, M.; Schachner, M.; Hartmann, U.; Cremer, H.; Lafon, M. The neural cell adhesion molecule is a receptor for rabies virus. J. Virol. 1998, 72, 7181–7190. [Google Scholar]
- Weir, D.L.; Smith, I.L.; Bossart, K.N.; Wang, L.F.; Broder, C.C. Host cell tropism mediated by Australian bat lyssavirus envelope glycoproteins. Virology 2013, 444, 21–30. [Google Scholar] [CrossRef]
- Badrane, H.; Tordo, N. Host switching in Lyssavirus history from the Chiroptera to the Carnivora orders. J. Virol. 2001, 75, 8096–8104. [Google Scholar] [CrossRef]
- Leslie, M.J.; Messenger, S.; Rohde, R.E.; Smith, J.; Cheshier, R.; Hanlon, C.; Rupprecht, C.E. Bat-associated rabies virus in Skunks. Emerg. Infect. Dis. 2006, 12, 1274–1277. [Google Scholar] [CrossRef]
- McColl, K.A.; Chamberlain, T.; Lunt, R.A.; Newberry, K.M.; Westbury, H.A. Susceptibility of domestic dogs and cats to Australian bat lyssavirus (ABLV). Vet. Microbiol. 2007, 123, 15–25. [Google Scholar]
- Ceballos, N.A.; Moron, S.V.; Berciano, J.M.; Nicolas, O.; Lopez, C.A.; Juste, J.; Nevado, C.R.; Setien, A.A.; Echevarria, J.E. Novel lyssavirus in bat, Spain. Emerg. Infect. Dis. 2013, 19, 793–795. [Google Scholar] [CrossRef]
- Virus Taxonomy: 2012 release. Available online: http://www.ictvonline.org/virusTaxonomy.asp (accessed on 18 December 2013).
- Gaudin, Y.; Ruigrok, R.W.; Tuffereau, C.; Knossow, M.; Flamand, A. Rabies virus glycoprotein is a trimer. Virology 1992, 187, 627–632. [Google Scholar]
- Watson, H.D.; Tignor, G.H.; Smith, A.L. Entry of rabies virus into the peripheral nerves of mice. J. Gen. Virol. 1981, 56, 372–382. [Google Scholar]
- Preuss, M.A.; Faber, M.L.; Tan, G.S.; Bette, M.; Dietzschold, B.; Weihe, E.; Schnell, M.J. Intravenous inoculation of a bat-associated rabies virus causes lethal encephalopathy in mice through invasion of the brain via neurosecretory hypothalamic fibers. PLoS Pathog. 2009, 5, e1000485. [Google Scholar] [CrossRef]
- Lentz, T.L.; Burrage, T.G.; Smith, A.L.; Crick, J.; Tignor, G.H. Is the acetylcholine receptor a rabies virus receptor? Science 1982, 215, 182–184. [Google Scholar]
- Lentz, T.L.; Wilson, P.T.; Hawrot, E.; Speicher, D.W. Amino acid sequence similarity between rabies virus glycoprotein and snake venom curaremimetic neurotoxins. Science 1984, 226, 847–848. [Google Scholar]
- Hanham, C.A.; Zhao, F.; Tignor, G.H. Evidence from the anti-idiotypic network that the acetylcholine receptor is a rabies virus receptor. J. Virol. 1993, 67, 530–542. [Google Scholar]
- Tuffereau, C.; Benejean, J.; Blondel, D.; Kieffer, B.; Flamand, A. Low-affinity nerve-growth factor receptor (P75NTR) can serve as a receptor for rabies virus. EMBO J. 1998, 17, 7250–7259. [Google Scholar]
- Superti, F.; Hauttecoeur, B.; Morelec, M.J.; Goldoni, P.; Bizzini, B.; Tsiang, H. Involvement of gangliosides in rabies virus infection. J. Gen. Virol. 1986, 67, 47–56. [Google Scholar] [CrossRef]
- Conti, C.; Superti, F.; Tsiang, H. Membrane carbohydrate requirement for rabies virus binding to chicken embryo related cells. Intervirology 1986, 26, 164–168. [Google Scholar] [CrossRef]
- Lafon, M. Rabies virus receptors. J. Neurovirol. 2005, 11, 82–87. [Google Scholar] [CrossRef]
- McGehee, D.S.; Role, L.W. Physiological diversity of nicotinic acetylcholine receptors expressed by vertebrate neurons. Annu. Rev. Physiol. 1995, 57, 521–546. [Google Scholar] [CrossRef]
- Burrage, T.G.; Tignor, G.H.; Smith, A.L. Rabies virus binding at neuromuscular junctions. Virus Res. 1985, 2, 273–289. [Google Scholar] [CrossRef]
- Tuffereau, C.; Desmezieres, E.; Benejean, J.; Jallet, C.; Flamand, A.; Tordo, N.; Perrin, P. Interaction of lyssaviruses with the low-affinity nerve-growth factor receptor p75NTR. J. Gen. Virol. 2001, 82, 2861–2867. [Google Scholar]
- Tuffereau, C.; Schmidt, K.; Langevin, C.; Lafay, F.; Dechant, G.; Koltzenburg, M. The rabies virus glycoprotein receptor p75NTR is not essential for rabies virus infection. J. Virol. 2007, 81, 13622–13630. [Google Scholar]
- Mercer, J.; Schelhaas, M.; Helenius, A. Virus entry by endocytosis. Annu. Rev. Biochem. 2010, 79, 803–833. [Google Scholar] [CrossRef]
- Superti, F.; Derer, M.; Tsiang, H. Mechanism of rabies virus entry into CER cells. J. Gen. Virol. 1984, 65, 781–789. [Google Scholar]
- Piccinotti, S.; Kirchhausen, T.; Whelan, S.P. Uptake of rabies virus into epithelial cells by clathrin-mediated endocytosis depends upon actin. J. Virol. 2013, 87, 11637–11647. [Google Scholar]
- Weir, D.L.; Laing, E.D.; Smith, I.L.; Wang, L.F.; Broder, C.C. Host cell virus entry mediated by Australian bat lyssavirus G envelope glycoprotein occurs through a clathrin-mediated endocytic pathway that requires actin and Rab5. Virol. J. 2014. submitted for publication. [Google Scholar]
- Sun, X.; Yau, V.K.; Briggs, B.J.; Whittaker, G.R. Role of clathrin-mediated endocytosis during vesicular stomatitis virus entry into host cells. Virology 2005, 338, 53–60. [Google Scholar] [CrossRef]
- Liu, H.; Liu, Y.; Liu, S.; Pang, D.W.; Xiao, G. Clathrin-mediated endocytosis in living host cells visualized through quantum dot labeling of infectious hematopoietic necrosis virus. J. Virol. 2011, 85, 6252–6262. [Google Scholar] [CrossRef]
- Ehrlich, M.; Boll, W.; van Oijen, A.; Hariharan, R.; Chandran, K.; Nibert, M.L.; Kirchhausen, T. Endocytosis by random initiation and stabilization of clathrin-coated pits. Cell 2004, 118, 591–605. [Google Scholar] [CrossRef]
- Johannsdottir, H.K.; Mancini, R.; Kartenbeck, J.; Amato, L.; Helenius, A. Host cell factors and functions involved in vesicular stomatitis virus entry. J. Virol. 2009, 83, 440–453. [Google Scholar] [CrossRef]
- Rust, M.J.; Lakadamyali, M.; Zhang, F.; Zhuang, X. Assembly of endocytic machinery around individual influenza viruses during viral entry. Nat. Struct. Mol. Biol. 2004, 11, 567–573. [Google Scholar]
- Cureton, D.K.; Massol, R.H.; Saffarian, S.; Kirchhausen, T.L.; Whelan, S.P. Vesicular stomatitis virus enters cells through vesicles incompletely coated with clathrin that depend upon actin for internalization. PLoS Pathog. 2009, 5, e1000394. [Google Scholar]
- Cureton, D.K.; Massol, R.H.; Whelan, S.P.; Kirchhausen, T. The length of vesicular stomatitis virus particles dictates a need for actin assembly during clathrin-dependent endocytosis. PLoS Pathog. 2010, 6, e1001127. [Google Scholar]
- Boulant, S.; Kural, C.; Zeeh, J.C.; Ubelmann, F.; Kirchhausen, T. Actin dynamics counteract membrane tension during clathrin-mediated endocytosis. Nat. Cell Biol. 2011, 13, 1124–1131. [Google Scholar] [CrossRef]
- Hinshaw, J.E.; Schmid, S.L. Dynamin self-assembles into rings suggesting a mechanism for coated vesicle budding. Nature 1995, 374, 190–192. [Google Scholar] [CrossRef]
- Sweitzer, S.M.; Hinshaw, J.E. Dynamin undergoes a GTP-dependent conformational change causing vesiculation. Cell 1998, 93, 1021–1029. [Google Scholar] [CrossRef]
- Bashkirov, P.V.; Akimov, S.A.; Evseev, A.I.; Schmid, S.L.; Zimmerberg, J.; Frolov, V.A. GTPase cycle of dynamin is coupled to membrane squeeze and release, leading to spontaneous fission. Cell 2008, 135, 1276–1286. [Google Scholar] [CrossRef]
- Ungewickell, E.; Ungewickell, H.; Holstein, S.E.; Lindner, R.; Prasad, K.; Barouch, W.; Martin, B.; Greene, L.E.; Eisenberg, E. Role of auxilin in uncoating clathrin-coated vesicles. Nature 1995, 378, 632–635. [Google Scholar] [CrossRef]
- Blumenthal, R.; Bali-Puri, A.; Walter, A.; Covell, D.; Eidelman, O. pH-dependent fusion of vesicular stomatitis virus with Vero cells. Measurement by dequenching of octadecyl rhodamine fluorescence. J. Biol. Chem. 1987, 262, 13614–13619. [Google Scholar]
- Van der Schaar, H.M.; Rust, M.J.; Waarts, B.L.; van der Ende-Metselaar, H.; Kuhn, R.J.; Wilschut, J.; Zhuang, X.; Smit, J.M. Characterization of the early events in dengue virus cell entry by biochemical assays and single-virus tracking. J. Virol. 2007, 81, 12019–12028. [Google Scholar] [CrossRef]
- Lakadamyali, M.; Rust, M.J.; Zhuang, X. Endocytosis of influenza viruses. Microbes Infect. 2004, 6, 929–936. [Google Scholar] [CrossRef]
- Lakadamyali, M.; Rust, M.J.; Zhuang, X. Ligands for clathrin-mediated endocytosis are differentially sorted into distinct populations of early endosomes. Cell 2006, 124, 997–1009. [Google Scholar] [CrossRef]
- Stenmark, H. Rab GTPases as coordinators of vesicle traffic. Nat. Rev. Mol. Cell Biol. 2009, 10, 513–525. [Google Scholar] [CrossRef]
- Semerdjieva, S.; Shortt, B.; Maxwell, E.; Singh, S.; Fonarev, P.; Hansen, J.; Schiavo, G.; Grant, B.D.; Smythe, E. Coordinated regulation of AP2 uncoating from clathrin-coated vesicles by rab5 and hRME-6. J. Cell Biol. 2008, 183, 499–511. [Google Scholar] [CrossRef]
- Mifune, K.; Ohuchi, M.; Mannen, K. Hemolysis and cell fusion by rhabdoviruses. FEBS Lett. 1982, 137, 293–297. [Google Scholar] [CrossRef]
- St Pierre, C.A.; Leonard, D.; Corvera, S.; Kurt-Jones, E.A.; Finberg, R.W. Antibodies to cell surface proteins redirect intracellular trafficking pathways. Exp. Mol. Pathol. 2011, 91, 723–732. [Google Scholar] [CrossRef]
- Lewis, P.; Fu, Y.; Lentz, T.L. Rabies virus entry into endosomes in IMR-32 human neuroblastoma cells. Exp. Neurol. 1998, 153, 65–73. [Google Scholar] [CrossRef]
- Lewis, P.; Lentz, T.L. Rabies virus entry into cultured rat hippocampal neurons. J. Neurocytol. 1998, 27, 559–573. [Google Scholar] [CrossRef]
- Gaudin, Y.; Ruigrok, R.W.; Knossow, M.; Flamand, A. Low-pH conformational changes of rabies virus glycoprotein and their role in membrane fusion. J. Virol. 1993, 67, 1365–1372. [Google Scholar]
- Weir, D.L.; Uniformed Services University, Bethesda, MD, USA. Unpublished work. 2014.
- Ugolini, G. Specificity of rabies virus as a transneuronal tracer of motor networks: Transfer from hypoglossal motoneurons to connected second-order and higher order central nervous system cell groups. J. Comp. Neurol. 1995, 356, 457–480. [Google Scholar] [CrossRef]
- Malgaroli, A.; Vallar, L.; Zimarino, V. Protein homeostasis in neurons and its pathological alterations. Curr. Opin. Neurobiol. 2006, 16, 270–274. [Google Scholar] [CrossRef]
- Schnell, M.J.; McGettigan, J.P.; Wirblich, C.; Papaneri, A. The cell biology of rabies virus: Using stealth to reach the brain. Nat. Rev. Microbiol. 2010, 8, 51–61. [Google Scholar]
- Albertini, A.A.; Baquero, E.; Ferlin, A.; Gaudin, Y. Molecular and cellular aspects of rhabdovirus entry. Viruses 2012, 4, 117–139. [Google Scholar] [CrossRef]
- CDNA National Guidelines for Public Health Units: Rabies Virus and Other Lyssavirus (Including Australian Bat Lyssavirus) Exposures and Infections; Communicable Disease Network Australia: Canberra, Australia, 2012.
- Brookes, S.M.; Parsons, G.; Johnson, N.; McElhinney, L.M.; Fooks, A.R. Rabies human diploid cell vaccine elicits cross-neutralising and cross-protecting immune responses against European and Australian bat lyssaviruses. Vaccine 2005, 23, 4101–4109. [Google Scholar] [CrossRef]
- Moore, P.R. Characterisation of Australian Bat Lyssavirus and an Evaluation of the Rabies Vaccines. Ph.D. Thesis, University of Queensland, Queensland, Australia, 2011. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Weir, D.L.; Annand, E.J.; Reid, P.A.; Broder, C.C. Recent Observations on Australian Bat Lyssavirus Tropism and Viral Entry. Viruses 2014, 6, 909-926. https://doi.org/10.3390/v6020909
Weir DL, Annand EJ, Reid PA, Broder CC. Recent Observations on Australian Bat Lyssavirus Tropism and Viral Entry. Viruses. 2014; 6(2):909-926. https://doi.org/10.3390/v6020909
Chicago/Turabian StyleWeir, Dawn L., Edward J. Annand, Peter A. Reid, and Christopher C. Broder. 2014. "Recent Observations on Australian Bat Lyssavirus Tropism and Viral Entry" Viruses 6, no. 2: 909-926. https://doi.org/10.3390/v6020909
APA StyleWeir, D. L., Annand, E. J., Reid, P. A., & Broder, C. C. (2014). Recent Observations on Australian Bat Lyssavirus Tropism and Viral Entry. Viruses, 6(2), 909-926. https://doi.org/10.3390/v6020909